A genome-wide association study on chronic HBV infection and
its clinical progression in male Han-Taiwanese
Su-Wei Chang1, Cathy Shen-Jang Fann2,Wen-Hui Su3, Yu Chen Wang4, Chia
Chan Weng4, Chia-Jung Yu5, Chia-Lin Hsu2, Ai-Ru Hsieh6, Rong-Nan
Chien4,Chia-Ming Chu4 and Dar-In Tai4
1Clinical Informatics and Medical Statistics Research Center, Chang Gung
University College of Medicine, Taoyuan, Taiwan; 2Institute of Biomedical
Sciences, Academia Sinica, Taipei, Taiwan; 3Department of Biomedical
Sciences, Graduate Institute of Biomedical Sciences, Chang Gung Molecular Medicine Research Center, Chang Gung University, Taoyuan, Taiwan; 4Division
of Hepatology, Department of Gastroenterology and Hepatology, Chang Gung Memorial Hospital at Linkou, Taoyuan, Taiwan; 5Department of Cell and Molecular Biology, College of Medicine and Molecular Medicine Research Center, Chang Gung University, Taoyuan, Taiwan; 6Graduate Institute of
Biostatistics, China Medical University, Taichung, Taiwan. E-mail:
Su-Wei Chang: [email protected]
Cathy Shen-Jang Fann: [email protected] Wen-Hui Su: [email protected]
Yu Chen Wang: [email protected] Chia Chan Weng: [email protected]
Chia-Jung Yu: [email protected] Chia-Lin Hsu: [email protected] Ai-Ru Hsieh: [email protected] Rong-Nan Chien: [email protected] Chia-Ming Chu: [email protected]
Dar-In Tai: [email protected] Corresponding Author:
Dar-In Tai, Division of Hepatology, Department of Gastroenterology and
of Medicine
199 Tung-Hwa North Road, Taipei, Taiwan 105 Tel: 886-3-3281200 ext 8107
Fax: 886-3-3272236
Abstract
It is common to observe the clustering of chronic hepatitis B surface antigen (HBsAg) carriers in families. Intra-familial transmission of hepatitis B virus (HBV) could be the reason for the familial clustering of HBsAg carriers. Additionally, genetic and gender factors have been reported to be involved. We conducted a three-stage genome-wide association study to identify genetic factors associated with chronic HBV susceptibility. A total of 1065 male controls and 1623 male HBsAg carriers were included. The whole-genome genotyping was done on Illumina HumanHap550 beadchips in 304 healthy controls and HumanHap610 beadchips in 321 cases. We found that rs9277535 (HLA-DPB1, P = 4.87x 10-14),
rs9276370 (HLA-DQA2, P = 1.9 x 10-12), rs7756516 and rs7453920 (HLA-DQB2,
P = 1.48 x 10-11 and P = 6.66 x 10-15 respectively) were significantly associated
with persistent HBV infection. A novel SNP rs9366816 near HLA-DPA3 also showed significant association (P = 2.58 x 10-10). The “T-T-G-G-T” haplotype of
the five SNPs further signified their association with the disease (P = 1.48×10-12;
OR = 1.49). The “T-T” haplotype composed of rs7756516 and rs9276370 was more prevalent in severe disease subgroups and associated with non-sustained therapeutic response (P = 0.0262). The “G-C” haplotype was associated with sustained therapeutic response (P = 0.0132; OR = 2.49). We confirmed that HLA-DPB1, HLA-DQA2 and HLA-DQB2 loci were associated with persistent HBV infection in male Taiwan Han-Chinese. In addition, the HLA-DQA2 and -DQB2 complex was associated with clinical progression and therapeutic response.
Keywords: antiviral therapy HBV, hepatitis B surface antigen, single nucleotide polymorphism, GWAS, HLA-DP, HLA-DQ
Introduction
It is common to observe clustering of chronic hepatitis B surface antigen (HBsAg) carriers and hepatocellular carcinoma (HCC) in families of affected individuals [1-5].A considerable number of patients exposed to HBV in the early stage of life became chronic HBsAg carriers [3, 6-8]. Most of the children born to the mothers with active HBV replications became HBsAg carriers. This type of perinatal infection gives little room for genetic roles to participate in [3].However, horizontal transmission is as important as perinatal infection in HCC families. The total number of HBsAg carriers was higher in children of male HCC patients (high horizontal transmission) than in children of female HCC patients (high perinatal infection) [5]. Genetic roles could be operating in patients with the horizontal transmission as well as in those mothers with inactive HBV
replications. Previous candidate gene studies suggested that human leukocyte antigen (HLA), cytokines,DNA repair and others were involved in HBV clearance or progression to hepatocarcinogenesis [8-10]. Recent genome-wide association studies (GWAS) confirmed that human gene variations in HLA-DP and HLA-DQ areas were associated with HBsAg persistence [11-12]. However, the clinical significance of these genes on HBV-related disease progression remained unclear. It could be shielded by multi-factorial problems associated with chronic HBV infection. It is well-known that the prevalence of HBsAg carriers in males is much higher than that in females [6]. Therefore, we conducted a GWAS in male Taiwan-Han Chinese to examine the differences between non-HBsAg carriers, inactive HBsAg carriers and active HBV related liver diseases.
Materials and Methods
Study Participants
A total of 1623 male case samples (321 in the GWAS scan, 646 in the first replication and 656 in the second replication) were recruited from Chang Gung Memorial Hospital (CGMH) at Linkou or Taiwan Liver Cancer Network (TLCN). The cases with age > 30 years were enrolled when they were seropositive for the HBsAg for > 6 months and seronegative for the anti-hepatitis C antibody (anti-HCV). All of the patients denied HIV infection. The 1623 HBsAg carriers were classified into three groups. The first group includes unrelated HBsAg carriers with persistent normal alanine aminotransferase (ALT) level (PNALT) for > 5 years and with HBV DNA < 105 cps/mL, which were carefully selected from
HBsAg carriers who visited the carrier clinic of Chang Gung Memorial Hospital and were followed for a long run [13, 14]. The second group was comprised of unrelated chronic hepatitis B (CHB) patients with fluctuated ALT levels and HBV DNA > 105 cps/mL. The third group included unrelated HCC patients. The age
and sample size information for this three-stage study is given in Table 1.
The 304 male control samples for the first stage GWAS scan were obtained from the Taiwan Han Chinese Cell and Genome Bank (who had no liver related disease and were seronegative for HBsAg and anti-HCV) [15]. The 761 male control individuals in the two replication studies were selected from local residents of Taoyuan County, Taiwan through a project designated for ‘‘Integrated Delivery System of Health Screening, Taoyuan, Taiwan’’. We included subjects of Han-Chinese origin with age > 30 years. The inclusion
criteria for the controls were seronegative for HBsAg and anti-HCV, and with normal liver biochemistry records. We did not use anti-Hepatitis B core protein (anti-HBc) in our inclusion criteria because nearly 90% of general population above the age of 30 were infected with HBV [16,17]. Those anti-HBc negative subjects who have been living in Taiwan, an HBV endemic area, for more than 30 years were not excluded because they had chances to be exposed to HBV, but may be genetically resistant to HBV infection [3].
This study as well as the informed consent was approved by the Ethics Committee of Chang Gung Memorial Hospital (IRB 94-581). A written informed consent was obtained from all the participants before the study. This clinical investigation had been conducted according to the principles expressed in the Declaration of Helsinki.
Genome-wide SNP genotyping and quality control
The genomic DNA was extracted from peripheral blood lymphocytes using the MagNA Pure LC DNA Isolation Kit with automatic DNA isolation instruments (MagNA Pure LC II; Roche Diagnostics GmbH, Mannheim, Germany). In the first-stage GWAS scan, the genome-wide SNP genotyping was done by Genizon Biosciences (Quebec, Canada) using Illumina HumanHap610 beadchips in the case group; while the genotyping for the control group was done by DeCode Genetics (Reykjavik, Iceland) using Illumina HumanHap550 beadchips. Only the overlapping SNPs on both chips were included in analysis. For filtering high-quality genotype data and for enlarging the number of potentially associated
SNPs, we carried out the following quality control procedures rather than the widely-used standard of the call rate of 95% and a minor allele frequency (MAF) of 5%; we included SNPs if: (1) a successful call rate in both cases and controls was > 90%; (2) an MAF in the controls was > 1%; (3) the Hardy-Weinberg equilibrium (HWE) was not violated in the controls (HWE test P-value > 10-7).
SNP Genotyping
In the two replication studies, the genotyping of the SNPs identified by GWAS was performed using TaqMan® Genotyping assays (Applied Biosystems Inc.) or Sequenom MassARRAY System (San Diego, CA). The experiment with TaqMan assays was done by Vita Genomics (New Taipei City, Taiwan), and the experiment with Sequenom MassArray was done by the Academia Sinica National Genotyping Center (Taipei, Taiwan).
Statistical analysis
We performed three single-locus association tests: genotype test, allele test and Cochran-Armitage trend test to compare allele and genotype frequencies assuming three modes of inheritance: additive, dominant and recessive between the cases and the controls for each SNP. To increase the chances of identifying potentially associated SNPs in the GWAS scan, a SNP marker with –log 10
(P-value) ≥ 5 in any of the three above mentioned tests would be considered for further analysis. In the first and the second replication studies, the significance level after Bonferroni correction for multiple comparisons (P = 0.05 divided by the
number of SNPs in the analysis) was used instead. The meta-analysis was then carried out using the Mantel-Haenszel method. The odds ratios (ORs) and 95% confidence intervals (CIs) were calculated by logistic regression analysis with age adjustment. The linkage disequilibrium (LD) map and values for the analyzed SNPs were generated by the Haploview software [18]. Haplotypes were inferred and maximum likelihood estimates of haplotype frequencies were generated by using the expectation-maximization (EM) algorithm. The haplotype categories with estimated frequencies of less than 5% were combined into one category called “All other”. Effects of each haplotype category with estimated frequencies of at least 5% on persistent HBV infection were then assessed between the HBsAg carriers and the controls using logistic regression.
To detect the effect of potential population stratification, we used the principle component analysis (PCA) based software “EIGENSTRAT” and
Multidimensional scaling analysis (MDS) of PLINK to investigate the structure of the first stage GWAS samples. Based on the distribution of the observed –log
10(trend test P-value) for the whole-genome SNPs against the theoretical
distribution of expected –log 10 (trend test P-value), the quantile-quantile (Q-Q)
plot was produced to imply the potential existence of HBV-associated genetic variants.
In the evaluation of the cumulative effects of the risk alleles identified, we calculated the ORs and 95% CIs using logistic regression models with the
median number of total risk alleles used as the reference group for the total 2688 individuals.
To assess the association between the clinical characteristics and the SNPs, the three clinical groups (PNALT, CHB and HCC) were investigated in the case-only analysis. Each of the three groups was coded as a binary trait and was tested for SNP associations using logistic regression models under assumptions of three modes of inheritance and with age adjustment. In the restrained samples who received any one of three anti-HBV treatment courses, the relationship between the SNP genotypes and the therapeutic response was evaluated using the Fisher’s exact test in each treatment group and the
Cochran-Mantel-Haenszel (CMH) test after controlling for treatment. In logistic regression analysis, the additive effect of risk alleles on the therapeutic response was assessed in each treatment group with age adjustment and in the full sample with age and treatment adjustment. In addition, association of inferred haplotype categories with therapeutic response was analyzed to evaluate the combined effects of multiple causal alleles and loci.
Results
Genome wide association study and replication
In the first-stage GWAS scan, the Illumina HumanHap610 beadchips
contained 598,821 SNPs in the cases and the Illumina HumanHap550 beadchips contained 560,184 SNPs in the controls. The gene chips covered 547,435 SNPs in common. After the quality control filtering, a total of 456,262 SNPs on the 22 autosomal chromosomes were obtained and then used in our initial discovery of
GWAS. The population structure (Figures S1A and S1B) and the Q-Q plot (Figure S2) using the whole genome genotype data of 321 cases and 304 controls indicated no evidence of population stratification between the HBV cases and the controls (inflation factor λ = 1.002).
A total of 61 SNPs which reached the significance level of 10-5 were found in
the whole genome (Table S1 and Figure S3). After eliminating the SNPs which failed to be validated by Taqman genotyping, we focused our study on thirty-eight SNPs on chromosome 6 (P < 10-5 in any of the three tests; Figure 1 and
Table S1). Based on the linkage disequilibrium (LD) block and the trend test P-value, fifteen SNPs were selected for the first replication in an independent cohort of 646 cases and 345 controls (Table S2). Seven SNPs showed significant association with P < 0.01 in the first replication.
Two SNPs rs9277554 and rs2051549 were not considered further due to their close proximity to rs9277535 and rs7453920. The other five significantly associated SNPs (rs9276370, rs7756516, rs7453920, rs9277535 and
rs9366816) in the first replication were selected for the second replication in an independent cohort with 656 cases and 416 controls.
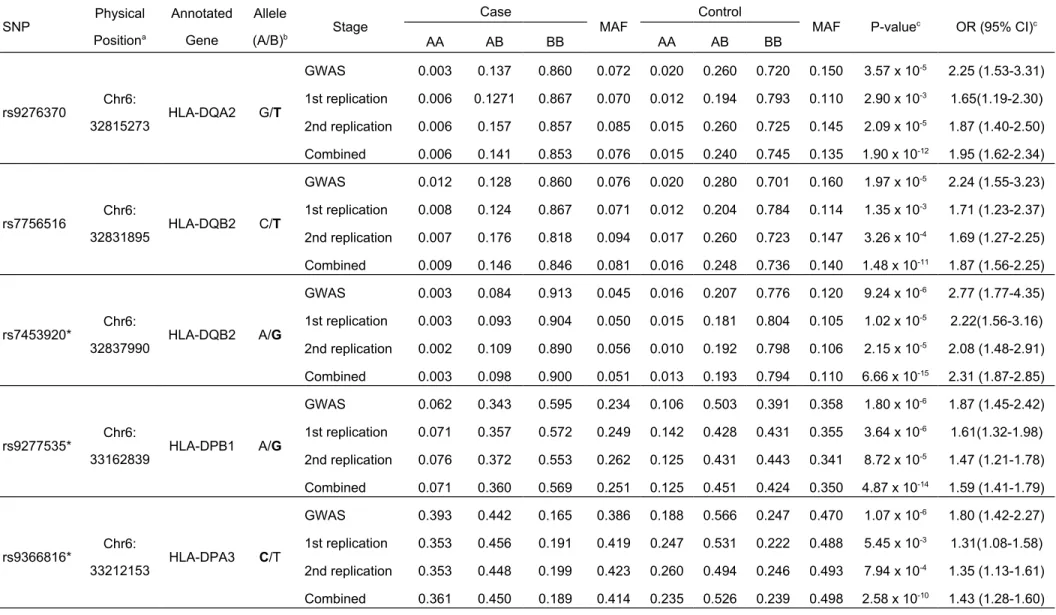
In the second replication, all the five SNPs rs9276370 (P = 2.09 x 10-5, OR =
1.87), rs7756516 (P = 3.26 x 10-4, OR = 1.69), rs7453920 (P = 2.15 x 10-5, OR =
2.08), rs9277535 (P = 8.72 x 10-5, OR = 1.47) and rs9366816 (P = 7.94 x 10-4,
OR = 1.35) remained significantly associated with HBV persistence after age adjustment (Table 2).
each single stage. The most significant association was conferred by rs7453920 (P = 6.66 x 10-15, OR = 2.31) on HLA-DQB2 and rs9277535 on HLA-DPB1 (P =
4.87x 10-14, OR = 1.59).
Pairwise LD patterns in the five associated SNPs and haplotype association
The pair wise LD of the five associated SNPs was calculated for the 625 individuals in the GWAS scan. The haplotype block was generated from the Haploview software (Figure 2). The LD block built up by the three SNPs
rs9276370, rs7756516 and rs7453920 suggested that they might not contribute independently to the disease risk. The rs9277535 and rs9366816 were
independent from the other three SNPs mentioned above, and they showed weak association with each other (r2 = 0.30). Hence we re-evaluated the
association of each one of the five SNPs using logistic regression analysis adjusted for the other specified SNP and age in the combined samples. As a result, the rs7453920 remained significant after controlling for the effect of
nearby rs9276370 and rs7756516 (Table S3; P-value = 5.49 x 10-4 and 4.06 x 10 -4, respectively). The additional rs9277535 on HLA-DPB1 and rs9366816 near
HLA-DPA3 were also associated with persistent HBV infections independently (P-value = 9.09 x 10-7). In the haplotype analysis, three major haplotypes
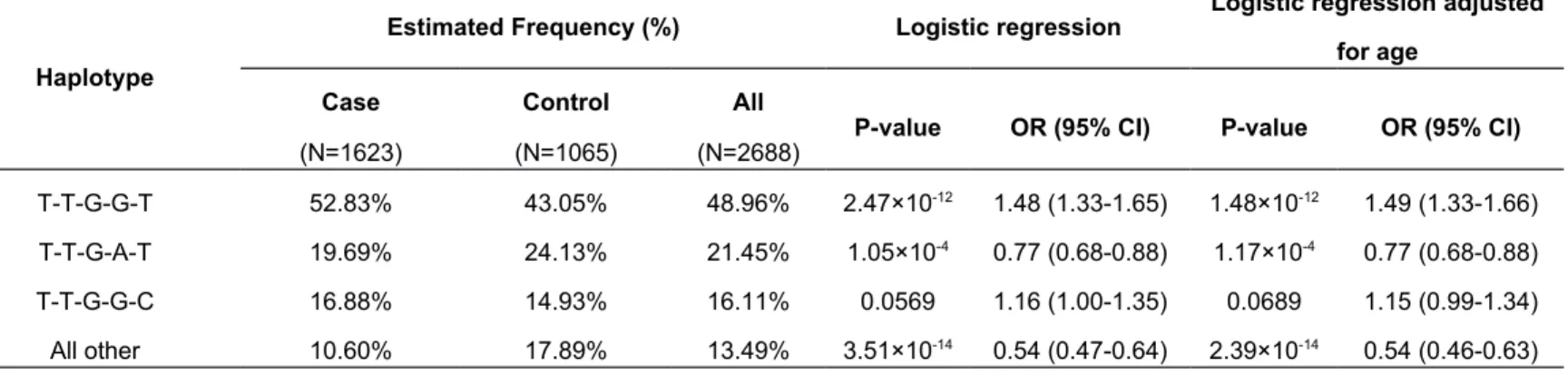
“T-T-G-G-T” (48.96%), “T-T-G-A-T” (21.45%) and “T-T-G-G-C” (16.11%) formed by the five SNPs were inferred (Table 4). After examining haplotype-disease
appeared to confer evident susceptibility to persistent HBV infection (52.83% in cases vs. 43.05% in controls; P-value = 1.48×10-12, OR = 1.49, 95% CI:
1.33-1.66 after age adjustment). We also found that the “T-T-G-A-T” haplotype had a protective effect against developing the disease (19.69% in cases vs. 24.13% in controls; P-value = 1.17×10-4, OR = 0.77, 95% CI: 0.68-0.88 after age
adjustment). Estimating and analyzing the haplotypes made up of four SNPs excluding rs9366816, the opposite effects of the rs9277535 G and A alleles contributing to persistent HBV infection appeared to be more prominent (Table S5).
Clinical significance of the SNPs
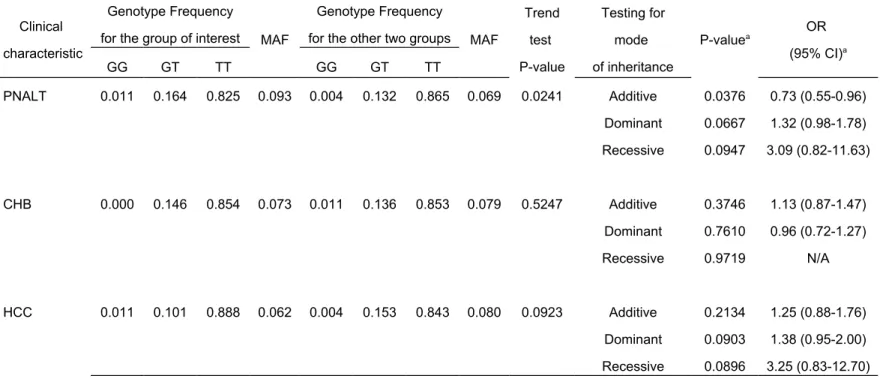
Among the 1623 HBsAg carriers, the SNP rs9276370 showed a lower TT genotype frequency in the PNALT group than in the other two groups (0.825 vs. 0.865; trend test p value=0.0241; Table 3). The test for additive effects of the T alleles reached significance with P =0.0376 (OR = 0.73; 95% CI: 0.55-0.96).
Those in the PNALT group were less likely to carry the rs7756516-T alleles compared to the other two more aggressive liver disease groups (CHB and HCC, Table S4). The frequency of the TT genotype was lower in the PNALT group than in the other two groups (0.815 vs 0.865; additive genetic effects P-value = 0.0249; OR = 0.74, 95% CI: 0.56-0.96; Table S4).
Therapeutic response
on the patients who received a course of the anti-HBV therapy. A variety of regimens developed in the past 20 to 30 years. We only considered the patients who received three widely used regimens: (1) lamivudine 100mg daily, which has been used widely since its availability in early 2000 [19]; (2) entevavir 0.5 mg daily, soon replacing lamivudine as the most widely used oral antiviral
agent accredited to its high potency of antiviral effects and low rate of drug resistance [20]; and (3) pegylated interferon α-2a 135-180 mcg weakly for one year [21]. Most of the patients received free therapies according to the
nationwide health insurance policy. The regimen initially covered a one-year therapy only but changed to an up to 4–year regimen in recent yearsinstead [22-24]. Therefore, we excluded those who received lamivudine or entecavir for < 1 year or > 4 year, and those who received pegylated interferon α-2a for < 1 year. A total of 226 patients were included for this analysis. Those patients with HBV DNA < 104 cps/mL with normal ALT at the 1-year post-treatment and then
remained persistent normal ALT, or transient ALT elevation to < 2 times the upper limit of normal during follow-up were defined as sustained responders [25-26]. Those patients with ALT > 2 times the upper limit of normal or HBV DNA > 104 cps/mL during the post-treatment follow-up period were defined as
non-sustained responders.
The treatment naive rate was highest in the lamivudine group (69.8%), followed by 40.6% in the entecavir group and 39.5% in the pegylated interferon α-2a group (P < 10-4; data not shown). The mean age at treatment was older in
cases, the frequency of GG and GT genotypes in the non-sustained responders was only 12.43% compared to 26.32% in the sustained responders (Fisher’s exact P = 0.0121 and CMH associated P = 0.0190). Similar trends were found in the treatment groups of lamivudine and pegylated interferon α-2a, but not in the entecavir group. Furthermore, in logistic regression analysis, the association of rs9276370 G alleles revealed significant additive effects on therapeutic response after adjusting for age and treatment (P = 0.0138, OR = 2.51, 95% CI: 1.21-5.22, Table 5). In the estimation of the haplotypes composed of two risk SNPs
rs9276370 and rs7756516, two major haplotypes “TT” (90.93%) and “GC” (7.74%) were obtained (Table S6). The “G-C” haplotype had a positive effect on sustained therapeutic response (P-value = 0.0132, OR = 2.49, 95% CI: 1.21-5.11 after adjusting for age at treatment); while the majority “T-T” haplotype showed a negative effect on sustained therapeutic response (P-value = 0.0262, OR = 0.46, 95% CI: 0.23-0.91 after adjusting for age at treatment) as given in Table S6. The result indicated the potential use of the “T-T” and “G-C” for distinguishing the two types of therapeutic responders, although further replication studies with larger sample size should definitely be carried out to validate their effects.
Discussion
From a total of 1065 male controls and 1623 male HBsAg carriers, this multi-stage GWAS study identified that three SNPs within HLA-DPB1 (rs9277535), HLA-DQB2 (7453920) and HLA-DPA3 (rs9366816) loci were
independently associated with persistence HBV infection in a male Taiwanese Han Chinese population. The former two SNP associations were also reported in previous Japanese and Mainland Chinese GWAS studies [11,12, 27]. Our findings confirmed that HLA-DP and -DQ loci played important roles in the development of persistent HBV infection.
A study from Mainland China revealed that HLA-DPA1 was highly
associated with persistent HBV infection in northern Han Chinese; while HLA-DPB1 was associated with the Southern minorities [28]. The Han Chinese in Taiwan mainly migrated from southern China [29]. This could be the reason why we did not identify SNPs in HLA-DPA1 in the GWAS scan. In addition to HLA-DP and HLA-DQ, a recent publication from Hu et al. (2013) reported that two novel SNPs rs3130542 (near HLA-C) and rs4821116 (in UBE2L3) were associated with persistent HBV infection in central China [27]. However, both of the SNPs did not show significant signals in our screening stage.
The rs9277535 in 3’UTR of HLA-DPB1 showed significant association with persistent HBV replication in our series (Table 2). The results of the haplotype analysis implied the crucial role that the rs9277535 SNP might play in the HBV persistence and viral clearance. In both of our haplotype analyses composed of five and four (including and excluding rs9366816), the individuals with the “T-T-G-G-T” or “T-T-G-G” haplotype were more likely to be persistent HBsAg carriers compared to those without the haplotypes (OR = 1.49 and 1.67 respectively); whereas those with the “T-T-G-A-T” or “T-T-G-A” haplotype were more likely to clear the HBV virus (Table 4 and S5).
In a study of European and Africa-American populations, rs9277534 rather than rs9277535 in the HLA-DPB1 3'UTR region was associated with HBV recovery [30]. In their flow cytometry and mRNA analysis, the HLA-DP expression level was higher with the GG genotype than with the other
genotypes. The higher HLA-DP expression might promote mechanisms of high HBV clearance in the controls [30, 31].
The SNP rs7453920 (HLA-DQB2) showed significant association with persistent HBV infection but was not associated with clinical outcome [32]. The SNP rs7756516 on 3’UTR of HLA-DQB2 identified in our cohort also showed significant association between the PNALT group and the other two more aggressive disease groups (CHB and HCC) (Table S4, the additive effect P = 0.0249; OR=0.74). There is a lower prevalence of rs7756516 genotype TT in the PNALT group than in the other two groups (0.815 vs. 0.865, trend test P =
0.0134). From the NIH web site, the highest prevalence of TT genotype of rs7756516 (0.788) was reported for the Han Chinese in Metropolitan Denver (HAPMAP-CHD). Their prevalence is similar to the prevalence in our controls (0.736) and lower than that in our cases (0.846, Table 2). These findings are compatible with a high prevalence of HBsAg in Taiwan [6].
The SNP rs9276370 near the 5’ region of HLA-DQA2 was also associated with persistent HBV infection (Table 1) and clinical progression (Table 3).
Together with data of HLA-DQB2, the HLA-DQA2 and HLA-DQB2 complex may participate in HBV infection persistence and disease progression. In the 226 cases who received a course of anti-HBV therapy, a higher non-sustained
responder rate was found in the TT genotype of rs9276370. This is especially true in the lamivudine group (P = 0.0074) and the pegylated interferon α-2a group (P = 0.0814, Table 4), whereas the entecavir group did not show this trend. The mean age at treatment in the lamivudine group was younger and it had been followed for a longer period (P =10-4, Table 5). It also had a higher
prevalence of treatment naive cases (69.8%) than the other two groups (40.6% and 39.5%; P <10-4). Nevertheless, whether these differences may explain the
therapeutic difference between Lamivudine and entecavir groups remains uncertain. Entecavir is a strong nucleoside analog that may suppress HBV replication completely [33]. Low HBV protein expression may decrease the impact of antigen presentation of HLA. The results of our haplotype analysis indicated the important role that HLA-DQA2 rs9276370 and HLA-DQB2
rs7756516 might play in therapeutic response. The “T-T” haplotype composed of rs9276370 and rs7756516 was associated with non-sustained therapeutic
response (P = 0.0262, Table S6); while the “G-C” haplotype was associated with sustained therapeutic response (P = 0.0132; OR = 2.49). The “T-T” and “G-C” haplotypes showed the potential for predicting the two types of therapeutic responders, but the sample size of the current analysis was too small and further replication studies should be carried out to validate the combined effects of these two SNPs.
Recently, Pan et al. discovered that four tagging SNPs rs477515,
rs28366298, rs3763316 and rs13204672 showed significant associations with non-response to hepatitis B vaccination [34]. The SNP rs477515 (located ∼12 kb
upstream of HLA-DRB1) as well as rs2516049 achieved the significance level in our initial screening stage, but we failed to validate them due to multiple
repetitive sequences near these SNPs.
The rs9366816 near HLA-DPA3 was associated with persistent HBV
infection (Figure 2 and Table S3). The rs9366816 is close to HLA-DPB2 as well. The information about these two HLA class II pseudogenes is limited [35]. The SNP rs9366816 in HLA-DPA3 is not in LD to with all the other associated SNPs except for rs9277535 (r2 = 0.30; Figure 2), which implies its independent
contribution to the disease susceptibility.
We concluded that HLA-DPB1, HLA-DQA2 and HLA-DQB2 were associated with persistent HBV infection in male Han Taiwanese. HLA-DQA2 and -DQB2 complex was associated with the clinical progression and therapeutic response of chronic HBV infection.
Acknowledgements
We would like to thank Translational Resource Center for Genomic Medicine (TRC) of National Research Program for Biopharmaceuticals (NRPB) for their
service. We also thank Taiwan Han Chinese Cell and Genome Bank of Academic Sinica, Taipei and Taiwan Liver Cancer Network (TLCN) for their support of human genomic DNA samples and National Center for Genome
Medicine (NCGM), Genomic Medicine Research Core Laboratory, Chang Gung Memorial Hospital, Linkou and Vita Genomics for their genotyping service. We
appreciate the Institute of Biomedical Sciences, Academia Sinica, Taiwan for their supports in the study.
References
1 Tai DI, Changchien CS, Hung CS, Chen CJ (1999) Replication of hepatitis B virus in first-degree relatives of patients with hepatocellular carcinoma. Am J Trop Med Hyg 61:16-9.
2 Sung JL, Chen DS (1977) Geographical distribution of the subtype of hepatitis B surface antigen in Chinese. Gastroenterol Jpn 12: 58-63.
3 Burk RD, Hwang LY, Ho GY, Shafritz DA, Beasley RP (1994) Outcome of perinatal hepatitis B virus exposure is dependent on maternal virus load. J Infect Dis 170:1418-23.
4 Tai DI, Chen CH, Chang TT, Chen SC, Liao LY, et al. (2002) Eight-year nationwide survival analysis in relatives of hepatocellular carcinoma: role of viral infection. J Gastroenterol Hepatol 17:682-9.
5 Chen CH, Chen YY, Chen GH, Yang SS, Tang HS, et al. (2004) Hepatitis B virus transmission and hepatocarcinogenesis: a 9 year retrospective cohort of 13676 relatives with hepatocellular carcinoma. J Hepatol 40:653-9.
6 Tsay PK, Tai DI, Chen YM, Yu CP, Wan SY, et al. (2009) Impact of Gender, Viral Transmission and Aging in the Prevalence of Hepatitis B Surface Antigen. Chang Gung Med J 32:155-64.
7 Beasley RP, Hwang LY, Lin CC, Leu ML, Stevens CE, et al. (1982) Incidence of hepatitis B virus infections in preschool children in Taiwan. J Infect Dis 146:198-204.
associations with hepatitis B and C viral infections across global populations. World J Gastroenterol 13:1770-87.
9 Chen CC, Yang SY, Liu CJ, Lin CL, Liaw YF, et al. (2005) Association of cytokine and DNA repair gene polymorphisms with hepatitis B-related
hepatocellular carcinoma. Int J Epidemiol 34:1310-8.
10 Yu MW, Yang SY, Pan IJ, Lin CL, Liu CJ, et al. (2003) Polymorphisms in XRCC1 and glutathione S-transferase genes and hepatitis B-related
hepatocellular carcinoma. J Natl Cancer Inst 95:1485-8.
11 Kamatani Y, Wattanapokayakit S, Ochi H, Kawaguchi T, Takahashi A, et al. (2009) A genome-wide association study identifies variants in the HLA-DP locus associated with chronic hepatitis B in Asians. Nat Genet 41:591-5. 12 Mbarek H, Ochi H, Urabe Y, Kumar V, Kubo M, et al. (2011) A
genome-wide association study of chronic hepatitis B identified novel risk locus in a Japanese population. Hum Mol Genet 20:3884-92.
13 Tai DI, Lin SM, Sheen IS, Chu CM, Lin DY, Liaw YF (2009) Long-term outcome of HBeAg-negative HBsAg carriers in relation to changes of alanine aminotransferase levels over time. Hepatology 49:1859-67.
14 Tai DI, Tsay PK, Chen WT, Chu CM, Liaw YF (2010) Relative role of HBsAg seroclearance and mortality in the decline of HBsAg prevalence with increasing age. Am J gastroenterol 105:1102-1109.
15 Pan WH, Fann CS, Wu JY, Hung YT, Ho MS, et al. (2006) Han Chinese cell and genome bank in Taiwan: purpose, design and ethical considerations. Hum Herred 61:27-30.
16 Chung DC, Ko YC, Chen CJ, Wu CC, Chen ER, et al. (1988)
Seroepidemiological studies on hepatitis B and D viruses infection among five ethnic groups in southern Taiwan. J Med Virol 26:411-8.
17 Lin-Chu M, Tsai SJ, Watanabe J, Nishioka K (1990) The prevalence of anti-HCV among Chinese voluntary blood donors in Taiwan. Transfusion 30:471-3.
18 Barrett JC, Fry B, Maller J, Daly MJ (2005) Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21:263-265. 19 Liaw YF, Sung JJ, Chow WC, Farrell G, Lee CZ, et al. (2004)
Lamivudine for patients with chronic hepatitis B and advanced liver disease. N Engl J Med 351:1521-31.
20 Chang TT, Liaw YF, Wu SS, Schiff E, Han KH, et al. (2010) Long-term entecavir therapy results in the reversal of fibrosis/cirrhosis and continued histological improvement in patients with chronic hepatitis B. Hepatology 52:886-93.
21 Liaw YF, Jia JD, Chan HL, Han KH, Tanwandee T, et al. (2011) Shorter durations and lower doses of peginterferon alfa-2a are associated with inferior hepatitis B e antigen seroconversion rates in hepatitis B virus genotypes B or C. Hepatology 54:1591-9.
22 Sullivan SD, Veenstra DL, Chen PJ, Daly MJ (2007) Cost-effectiveness of peginterferon alpha -2a compared to lamivudine treatment in patients with hepatitis B e antigen positive chronic hepatitis B in Taiwan. J Gastroenterol Hepatol 22:1494-9.
23 Veenstra DL, Sullivan SD, Lai MY, Lee CM, Tsai CM, Patel KK (2008) HBeAg-negative chronic hepatitis B: cost-effectiveness of peginterferon alfa -2a compared to lamivudine in Taiwan. Value Health 11:131-8.
24 Dai CY, Tseng TC, Wong GL, Huang JF, Wong VW, et al. (2013) Consolidation therapy for HBeAg-positive Asian chronic hepatitis B patients receiving lamivudine treatment: a multicentre study. J Antimicrob Chemother 68:2332-8.
25 Liaw YF, Leung N, Kao JH, Piratvisuth T, Gane E, et al. (2008) Chronic Hepatitis B Guideline Working Party of the Asian-Pacific Association for the Study of the Liver. Asian-Pacific consensus statement on the management of chronic hepatitis B: a 2008 update. Hepatol Int 2:263-83.
26 Jeng WJ, Sheen IS, Chen YC, Hsu CW, Chien RN, et al. (2013) Off-therapy durability of response to entecavir Off-therapy in hepatitis B e antigen-negative chronic hepatitis B patients. Hepatology 58:1888-96.
27 Hu Z, Liu Y, Zhai X, Dai J, Jin G, et al. (2013) New loci associated with chronic hepatitis B virus infection in Han Chinese. Nat Genet 45:1499-503. 28 Wang L, Wu XP, Zhang W, Zhu DH, Wang Y, et al. (2011) Evaluation of
genetic susceptibility loci for chronic hepatitis B in Chinese: two independent case-control studies. PLoS One 6(3):e17608.
29 Shaw CK, Chen LL, Lee A, Lee TD (1999) Distribution of HLA gene and haplotype frequencies in Taiwan: a comparative study among Min-nan, Hakka, Aborigines and Mainland Chinese. Tissue Antigens 53:51-64.
30 O'Brien TR, Kohaar I, Pfeiffer RM, Maeder D, Yeager M, et al. (2011) Risk alleles for chronic hepatitis B are associated with decreased mRNA expression of HLA-DPA1 and HLA-DPB1 in normal human liver. Genes Immun 12:428-33.
31 Thomas R, Thio CL, Apps R, Qi Y, Gao X, et al. (2012) A novel variant marking HLA-DP expression levels predicts recovery from hepatitis B virus infection. J Virol 86:6979-85.
32 Al-Qahtani AA, Al-Anazi MR, Abdo AA, Sanai FM, Al-Hamoudi W, et al. (2014) Association between HLA Variations and Chronic Hepatitis B Virus Infection in Saudi Arabian Patients. PLoS One 9:e80445.
33 de Man RA, Wolters LM, Nevens F, Chua D, Sherman M, et al. (2001) Safety and efficacy of oral entecavir given for 28 days in patients with chronic hepatitis B virus infection. Hepatology 34:578-82.
34 Pan L, Zhang L, Zhang W, Wu X, Li Y, et al. (2014) A genome-wide association study identifies polymorphisms in the HLA-DR region associated with non-response to hepatitis B vaccination in Chinese Han populations. Hum Mol Genet 23:2210-9.
35 Stephens R, Horton R, Humphray S, Rowen L, Trowsdale J, Beck S (1999) Gene organisation, sequence variation and isochore structure at the centromeric boundary of the human MHC. J Mol Biol 291:789-99.
Figure legends
Figure 1 GWAS results
The multi-stage signals of association with HBV infection in the HLA region were presented.
Figure 2 LD patterns of the 5 SNPs
The LD patterns were based on the D’ (left side) and r2 (right side) measures of
the five associated SNPs for 625 individuals in the GWAS scan. The LD block built up by the rs9276370, rs7756516 and rs7453920 suggested that they were significantly associated with each other. The rs9366816 and rs9277535 SNPs were independent from each other and from the other three SNPs.
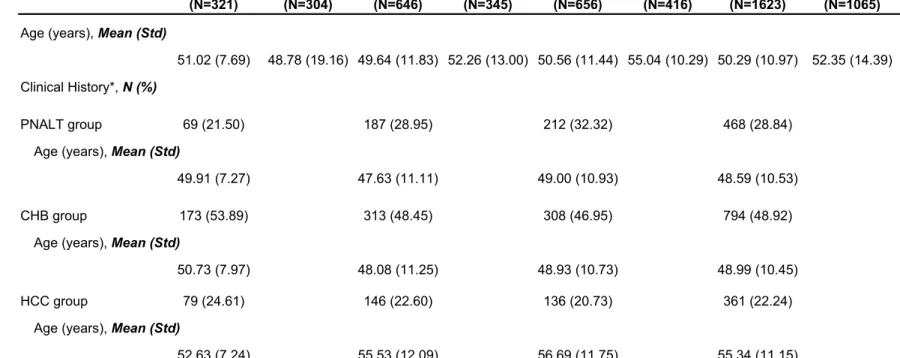
Table 1 Descriptive characteristic of the three-stage study samples in different clinical groups
Characteristics
GWAS Scan 1st replication 2nd replication Combined
Case (N=321) Control (N=304) Case (N=646) Control (N=345) Case (N=656) Control (N=416) Case (N=1623) Control (N=1065)
Age (years), Mean (Std)
51.02 (7.69) 48.78 (19.16) 49.64 (11.83) 52.26 (13.00) 50.56 (11.44) 55.04 (10.29) 50.29 (10.97) 52.35 (14.39) Clinical History*, N (%)
PNALT group 69 (21.50) 187 (28.95) 212 (32.32) 468 (28.84) Age (years), Mean (Std)
49.91 (7.27) 47.63 (11.11) 49.00 (10.93) 48.59 (10.53) CHB group 173 (53.89) 313 (48.45) 308 (46.95) 794 (48.92)
Age (years), Mean (Std)
50.73 (7.97) 48.08 (11.25) 48.93 (10.73) 48.99 (10.45) HCC group 79 (24.61) 146 (22.60) 136 (20.73) 361 (22.24)
Age (years), Mean (Std)
Table 2 Summary of results for the three-stage association study SNP Physical Positiona Annotated Gene Allele (A/B)b Stage Case
MAF Control MAF P-valuec OR (95% CI)c
AA AB BB AA AB BB rs9276370 Chr6: 32815273 HLA-DQA2 G/T GWAS 0.003 0.137 0.860 0.072 0.020 0.260 0.720 0.150 3.57 x 10-5 2.25 (1.53-3.31) 1st replication 0.006 0.1271 0.867 0.070 0.012 0.194 0.793 0.110 2.90 x 10-3 1.65(1.19-2.30) 2nd replication 0.006 0.157 0.857 0.085 0.015 0.260 0.725 0.145 2.09 x 10-5 1.87 (1.40-2.50) Combined 0.006 0.141 0.853 0.076 0.015 0.240 0.745 0.135 1.90 x 10-12 1.95 (1.62-2.34) rs7756516 Chr6: 32831895 HLA-DQB2 C/T GWAS 0.012 0.128 0.860 0.076 0.020 0.280 0.701 0.160 1.97 x 10-5 2.24 (1.55-3.23) 1st replication 0.008 0.124 0.867 0.071 0.012 0.204 0.784 0.114 1.35 x 10-3 1.71 (1.23-2.37) 2nd replication 0.007 0.176 0.818 0.094 0.017 0.260 0.723 0.147 3.26 x 10-4 1.69 (1.27-2.25) Combined 0.009 0.146 0.846 0.081 0.016 0.248 0.736 0.140 1.48 x 10-11 1.87 (1.56-2.25) rs7453920* Chr6: 32837990 HLA-DQB2 A/G GWAS 0.003 0.084 0.913 0.045 0.016 0.207 0.776 0.120 9.24 x 10-6 2.77 (1.77-4.35) 1st replication 0.003 0.093 0.904 0.050 0.015 0.181 0.804 0.105 1.02 x 10-5 2.22(1.56-3.16) 2nd replication 0.002 0.109 0.890 0.056 0.010 0.192 0.798 0.106 2.15 x 10-5 2.08 (1.48-2.91) Combined 0.003 0.098 0.900 0.051 0.013 0.193 0.794 0.110 6.66 x 10-15 2.31 (1.87-2.85) rs9277535* Chr6: 33162839 HLA-DPB1 A/G GWAS 0.062 0.343 0.595 0.234 0.106 0.503 0.391 0.358 1.80 x 10-6 1.87 (1.45-2.42) 1st replication 0.071 0.357 0.572 0.249 0.142 0.428 0.431 0.355 3.64 x 10-6 1.61(1.32-1.98) 2nd replication 0.076 0.372 0.553 0.262 0.125 0.431 0.443 0.341 8.72 x 10-5 1.47 (1.21-1.78) Combined 0.071 0.360 0.569 0.251 0.125 0.451 0.424 0.350 4.87 x 10-14 1.59 (1.41-1.79) rs9366816* Chr6: 33212153 HLA-DPA3 C/T GWAS 0.393 0.442 0.165 0.386 0.188 0.566 0.247 0.470 1.07 x 10-6 1.80 (1.42-2.27) 1st replication 0.353 0.456 0.191 0.419 0.247 0.531 0.222 0.488 5.45 x 10-3 1.31(1.08-1.58) 2nd replication 0.353 0.448 0.199 0.423 0.260 0.494 0.246 0.493 7.94 x 10-4 1.35 (1.13-1.61) Combined 0.361 0.450 0.189 0.414 0.235 0.526 0.239 0.498 2.58 x 10-10 1.43 (1.28-1.60)
Table 3 Association analysis of rs9276370 with clinical classifications in HBsAg positive carriers
Clinical characteristic
Genotype Frequency
for the group of interest MAF
Genotype Frequency
for the other two groups MAF
Trend test P-value Testing for mode of inheritance P-valuea OR (95% CI)a GG GT TT GG GT TT PNALT 0.011 0.164 0.825 0.093 0.004 0.132 0.865 0.069 0.0241 Additive 0.0376 0.73 (0.55-0.96) Dominant 0.0667 1.32 (0.98-1.78) Recessive 0.0947 3.09 (0.82-11.63) CHB 0.000 0.146 0.854 0.073 0.011 0.136 0.853 0.079 0.5247 Additive 0.3746 1.13 (0.87-1.47) Dominant 0.7610 0.96 (0.72-1.27) Recessive 0.9719 N/A HCC 0.011 0.101 0.888 0.062 0.004 0.153 0.843 0.080 0.0923 Additive 0.2134 1.25 (0.88-1.76) Dominant 0.0903 1.38 (0.95-2.00) Recessive 0.0896 3.25 (0.83-12.70)
Table 4 Haplotype association of the 5 risk SNPs rs9276370, rs7756516, rs7453920, rs9277535 and rs9366816 with persistent HBV infection
Haplotype
Estimated Frequency (%) Logistic regression Logistic regression adjusted
for age
Case Control All
P-value OR (95% CI) P-value OR (95% CI)
(N=1623) (N=1065) (N=2688)
T-T-G-G-T 52.83% 43.05% 48.96% 2.47×10-12 1.48 (1.33-1.65) 1.48×10-12 1.49 (1.33-1.66)
T-T-G-A-T 19.69% 24.13% 21.45% 1.05×10-4 0.77 (0.68-0.88) 1.17×10-4 0.77 (0.68-0.88)
T-T-G-G-C 16.88% 14.93% 16.11% 0.0569 1.16 (1.00-1.35) 0.0689 1.15 (0.99-1.34) All other 10.60% 17.89% 13.49% 3.51×10-14 0.54 (0.47-0.64) 2.39×10-14 0.54 (0.46-0.63)
Table 5 Association of the rs9276370 genotypes with therapeutic response in three treatment groups Treatment group Age at treatment (year) Mean ± SD Post-treatment follow-up (month) Mean ± SD rs9276370 Genotype
Therapeutic response Fisher’s exact p-value Logistic regressionb Non-sustained N (%) Sustained N (%) P-value b OR (95% CI)b Lamivudine 100 mg/daily for 1-4 years 45.49 ± 10.05 77.78 ± 26.78 GG 0 (0) 1 (3.57) 0.0074 0.0037 5.41 (1.73-16.95) GT 7 (7.69) 7 (25.00) TT 84 (92.31) 20 (71.43) Entecavir 0.5 mg/daily for 1-4 years 51.58 ± 8.65 38.34 ± 15.90 GT 11 (23.40) 3 (17.65) 0.7425 0.5954 0.68 (0.16-2.86) TT 36 (76.60) 14 (82.35) Pegasys 135-180 mcg/ weakly for 1 year
45.95 ± 8.11 39.02 ± 20.30 GT 3 (9.68) 4 (33.33) 0.0814 0.0676 5.44 (0.88-33.48) TT 28 (90.32) 8 (66.67) Total 47.30 ± 9.67 59.24 ± 30.13 GG 0 (0) 1 (1.75) 0.0121 (CMH Pa= 0.0190) 0.0138 2.51 (1.21-5.22) GT 21 (12.43) 14 (24.56) TT 148 (87.57) 42 (73.68)
aCMHP: The Cochran-Mantel-Haenszel(CMH) p-value for general association was tested for the relationship between the rs9276370 genotypes and
the therapeutic response after controlling for treatment.
bThe additive G allele effect on the therapeutic response(sustained vs. non-sustained) was evaluated in logistic regression analysis. For each of the three
Supplementary Information Legends
Figures S1A and S1B Population structure
The population structure was estimated by (A) principle component analysis (PCA) from EIGENSTRAT; and (B) multidimensional scaling analysis (MDS) from PLINK in the first stage GWAS samples.
Figure S2 Quantile-Quantile (Q-Q) plot
The Q-Q plot was generated from the trend test result of 321 cases and 304 controls in the first stage GWAS scan.
Figure S3 Manhattan plot for the GWAS of HBV infection
The Manhattan plot indicating the -log10 P from the Cochran-Armitage trend test
of 456,262 SNPs was plotted against their physical positions on the 22 autosomal chromosomes.