中 國 醫 藥 大 學
新進教師及研究人員學術研究經費補助成果報告
計畫名稱:宿主 T 細胞影響沙門氏桿菌抑制腫瘤之功效
計畫編號:CMU 99–N2-08
執行期限: 2011 年 4 月 1 日至 2012 年 3 月 31 日
單位名稱:醫學系微生物科
主持人:李哲欣
中
華
民
國 1 0 1 年
3
月
8
日
T cell augments the antitumor activity of tumor-targeting Salmonella
Abstract
Systemic administration of Salmonella to tumor-bearing mice leads to the preferential accumulation
within tumor sites and retardation the tumor growth. However, the detailed mechanism of the
Salmonella-induced antitumor immune response via host T cell remained uncertain. Herein, we
used wild-type, CD4+T-cell-deficient, and CD8+T-cell-deficient mice to study the role of T cell in
the antitumor immune responses induced by Salmonella enterica serovar Choleraesuis (S.
Choleraesuis). When systemically administered into mice bearing tumors, S. Choleraesuis
significantly inhibited tumor growth by 50%. By contrast, in T-cell-deficient mice, there was only
34%~42% in inhibition of tumor growth. We found that the treatment of S. Choleraesuis
significantly upregulate interferon-γin wild-type and CD8+T-cell-deficient mice, but not in CD4+
T-cell-deficient mice. Furthermore, immunohistochemical staining of the tumors revealed more
infiltration of macrophages, and neutrophils in wild-type mice after S. Choleraesuis treatment
compared with those in T-cell-deficient mice. The antitumor therapeutic effect mediated by S.
Choleraesuis is associated with an inflammatory immune response at tumor site and a tumor T
helper 1-type immune response. In conclusion, these results suggest that tumor-targeted therapy
using S. Choleraesuis, which exerts tumoricidal effects and stimulates T cell activities, represents a
potential strategy for the treatment of tumor.
Introduction
The exploration of wild-type and genetically modified bacteria strains as potential
therapeutics in the treatment of cancer has been inspired by the incidental observations of cancer
regression following spontaneous bacterial infection. Studies in which cancer patients were
infected with Steptococcus pyogenes were initiated by Busch in 1868, Fehleisen in 1883 and Coley
in 1891(Pawelek et al. 2003) In 1964, a series of reports described the use of nonpathogenic
Clostridia in experimental treatment of cancer in rodents (Engelbart and Gericke 1964; Thiele et al.
1964). The rationale for using Clostridium as an anticancer agent is based on the fact that the
hypoxic regions of solid tumors provide anaerobic bacteria such as Clostridium a suitable
environment to germinate and growth. The proliferation of the bacteria presumably occurs in the
hypoxic and necrotic regions of the tumor, but not in well-oxygenated normal tissues. Although
promising, this antitumor strategy may have major limitations. Tumor lysis is restricted to large,
well-establish tumors but is undetectable in smaller metastatic deposits, probably because of the
lack of hypoxic regions in these lesions. Apart from obligate anaerobes which target
hypoxic/necrotic areas of solid tumors, Salmonella typhimurium, a facultative anaerobe capable of
growing under both aerobic and anaerobic conditions, has also been exploited as a potential
oncolytic agent (Pawelek et al.1997; Low et al. 1999). Salmonellae are gram-negative, facultative
anaerobes that are a common cause of intestinal infections. As Salmonellae survive and replicate
in both oxygenated and hypoxic conditions, they would be expected to colonize small metastatic
desirable properties as tumor-targeted anticancer agents, which include targeting of tumors from a
distant inoculation site, selective replication within tumors, and tumor growth suppression.
Although Salmonella possesses properties of targeting of multiple tumors, selective replication
within tumors, tumor retardation, and gene delivery, the mechanisms contributing to its initial
infection to tumors, preferential accumulation within tumors to high levels relative to normal
tissues, and oncolytic effects are largely not understood. There are some factors influenced the
tumor colonization of Salmonella. It has been shown that Salmonella has limited ability to adhere
to tumor vasculature and migrate within tumors and only survives in tissue that becomes necrotic
(Forbes et al. 2003). The immune response against Salmonella is composed of an immediate
response mediated by the innate arm of the immune system followed by antigen-specific adaptive
immunity. Together, these two arms of immune system work to eradicate infection and provide
long-lasting immunity and memory. To improve the efficacy and safety of the oncolytic therapy of
Salmonella, a further understanding of Salmonella interactions with the immune system is
required.
The attenuated Salmonella enterica serovar Choleraesuis (S. Choleraesuis) described here has
been used previously as a live vaccine for swine and as a DNA vaccine delivery vector for mice
(Shiau et al. 2001). However, its genetic characteristics have not been well defined. We previously
reported to exploit an attenuated S. Choleraesuis as an antitumor agent capable of preferentially
accumulating and amplifying within tumors, and as a gene delivery vector to transfer gene at the
been used as an antitumor agent in previous studies (Pawelek et al. 1997; Low et al. 1999). Yu et al.
demonstrated that not only did attenuated bacterial strains, such as S. typhimurium, Vibrio cholerae,
and Listeria monocytogenes, gain entry and replicate in the tumor tissue, E. coli DH5α strain also
exhibited tumor-specific localization, indicating that no mutations affecting the survival of the
bacteria are required for tumor-targeted potential (Yu et al. 2004).
The use of T-cell-deficient mice allowed us to analyze the role of T cells in tumor-bear mice
after Salmonella administration. These results not only indicate that mechanisms that T cells are
important for control of systemic Salmonella treatment, but also that T cells participate in antitumor
activities of Salmonella. Herein, we want to identify that T cell involved in the antitumor effector of
Material and methods
Cells, Bacteria and Mice
Lewis lung carcinoma (LL2) was cultured in Dulbecco’smodified Eagle’smedium (DMEM) supplemented with 50 μg/mlgentamicin,2 mM L-glutamine, and 10% heat-inactivated fetal bovine serum (FBS) at 37℃ in 5% CO2. A vaccine strain of S. Choleraesuis (ATCC 15480) was obtained
from Bioresources Collection and Research Center (Hsinchu, Taiwan). This rough variant of S.
Choleraesuis, designated vaccine 51, was obtained by spreading an 18-h broth culture of the
virulent strain 188 of S. Choleraesuis strain Dublin over the surface of a dried nutrient agar plate
and placing a drop of a suspension of salmonella anti-O phage no. 1, and selecting for a
phage-resistant colony after incubation at 37oC for 24 h (Lee et al. 2009). Male C57BL/6 mice at
the age of 6 to 8 weeks were obtained from the Laboratory Animal Center of the National Cheng
Kung University. Male CD8+, and CD4+T -cell-deficient mice were purchased from the Jackson
and was approved by the Laboratory Animal Care and Use Committee of the National Cheng Kung
University.
Animal Studies
The mice were inoculated subcutaneously (s.c.) with 106tumor cells. When the tumors had grown
to 50 mm3to 100 mm3, the mice were injected intravenously (i.v.) with 2 × 106cfu of S.
Choleraesuis. At various time points postinfection, 3-4 mice in each group were sacrificed, and the
numbers of S. Choleraesuis in the tumors, livers, and spleens were determined on LB agar plates
and expressed as cfu per gram of tissues. In a separate experiment, palpable tumors were measured
every 3 days in two perpendicular axes with a tissue caliper and the tumor volume was calculated as:
(length of tumor) × (width of tumor)2× 0.45.
Assessment of Cytokines and Immunohistochemical Staining
To determine the expression of IFN-γafter S. Choleraesuis injection, mice were inoculated with
LL2 cells (106) at day 0. Then, the groups of mice were treated with S. Choleraesuis (2 × 106cfu)
by intravenous injection at day 10. To detect the protein and cytokine expressions, the organs were
collected at day 11. Levels of IFN-γin the tissue homogenates or sera were determined by ELISA
(R&D, Minneapolis, MN). The protein content in each sample was determined by bicinchoninic
acid (BCA) protein assay (Pierce Biotechnology, Rockford, IL). To analyze cell infiltrates in the
with either 2 × 106cfu of S. Choleraesuis at day 7. Control mice received PBS. The tumors were
excised and snap frozen at time point.Cryostatsections(5 μm)werealso prepared,fixed,and
incubated with rat anti-mouse Ly-6G (Gr-1) (RB6-8C5, BD Biosciences, San Diego, CA), rat
anti-mouse Mac-3 (M3/84, BD Biosciences), rat anti-mouse CD4 (L3T4) (GK1.5, BD Biosciences),
or rat anti-mouse CD8a (Ly-2) (53-6.7, BD Biosciences) antibody. After sequential incubation with
appropriate peroxidase-labeled secondary antibody and aminoethyl carbazole (AEC) as substrate
chromogen, the slides were counterstained with hematoxylin. The infiltrating cells were quantified
by averaging the number of each cell type in three areas of highest cell density at × 200
magnification in each section (Lee et al. 2008a).
Statistical Analysis
The unpaired, two-t
ai
l
ed
St
udent
’
s
t test was used to
determine differences between groups for the comparisons
of tumor volume, tumor weight, and the numbers of
macrophages, and neutrophils. Any p value less than 0.05 is
regarded statistically significant.
Results
Increased infiltrating CD4+and CD8+ T cells in the tumors following the treatment of
Salmonella
The persistence of high levels of Salmonella within tumors may induce inflammatory responses,
leading to the recruitment of immune cells to the tumor site (Lee et al. 2008a; Lee et al. 2004; Lee
et al. 2005). The clearance of Salmonella from infected tissues is controlled by the acquisition of
adaptive immunity. The tumors from mice bearing the LL2 tumors treated with S. Choleraesuis
were analyzed for cell infiltrates by immunohistochemical staining. Representative results for
immunohistochemistry are shown in Fig. 1a. Notable increases of CD8+and CD4+T cells that
infiltrated into the tumors were observed in the mice treated with S. Choleraesuis (Fig. 1b). These
results indicate that S. Choleraesuis resulted in increasing infiltrating CD8+and CD4+T cells in the
tumors. As bacterial replication in tumors and subsequent lysis of tumor cells may induce
cell-mediated immune responses to tumor cells, higher oncolysis could account, in part, for an
increased infiltrate of T cells in S. Choleraesuis-treated tumors. The T cell response against tumor
cells presumably enhanced the antitumor efficacy of S. Choleraesuis.
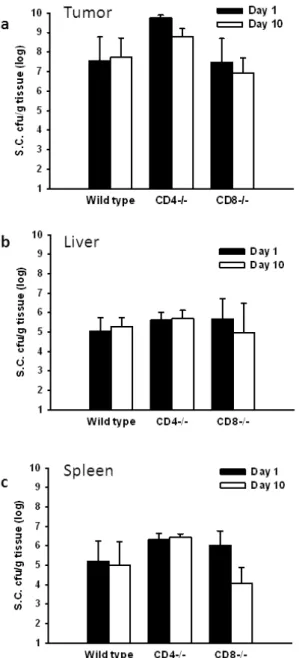
Tissue distributions of Salmonella in mice
Previously, We investigated the impact of host immune responses on the tumor-targeting
Choleraesuis, and the amounts of accumulated S. Choleraesuis in the tumors, livers, and spleens
were determined at various time points (Lee et al. 2004). The bacterial amount was much higher in
the tumors than in the livers and spleens at all the time points examined in both strains of mice.
Their amounts in the tumors reached a peak level at day 2 and maintained in similar high levels
during the first 6 days. Notably, bacterial load in the tumors was reduced by approximately 4 orders
of magnitude in BALB/c mice at day 12, whereas there was no equivalent decrease in the bacterial
number in NOD/SCID mice. Similarly, there was a decline in the bacterial numbers in the livers
and spleens at 6 days and further decreased to undetectable levels in BALB/c mice at day 12. In
contrast, in NOD/SCID mice bacterial levels in the three tissues remained fairly constant and
persisted for at least 12 days, with constant 10000:1 ratio of bacterial accumulation in the tumor to
accumulation in the liver or spleen. Therefore, our data, combined with those reported previously
(Forbes et al. 2003), suggest that the abilities of Salmonella in initial localization and subsequent replication within tumorsarenotdependenton the host’simmunesystem;however,by around 1 week when adaptive immune responses are elicited, Salmonella may be cleared more rapidly from
livers and spleens, where abundant immune cells are present, than from tumors by host immune
surveillance, thereby persisting longer within the tumor. Herein, to investigate the impact of T cell
on the tissue distributions of Salmonella, the wild-type ,CD4+T-cell-deficient, and CD8+
T-cell-deficient mice bearing tumors were injected with S. Choleraesuis and the amounts of S.
Choleraesuis in the tumors, livers, and spleens were determined at day 1 and day10. As shown in
points in all examined mice. Their amounts in the tumors maintained in high levels during the 10
days. Notably, bacterial load in the tumors was less in wild-type mice than that in CD4+
T-cell-deficient mice (Fig. 2a). The amounts of S. Choleraesuis in tumors were not significantly
different between wild-type and CD8+T-cell-deficient mice. Taken together, these results suggest
that CD4+- T-cell influenced the tumor-targeting potential of S. Choleraesuis.
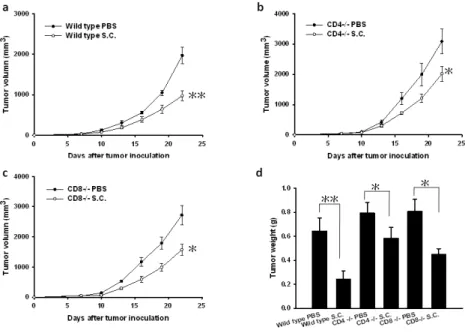
Antitumor effects of Salmonella on T-cell-deficient and wild-type mice
We next compared the antitumor effects of systemic administration of S. Choleraesuis on wild-type,
CD4+T-cell-deficient, and CD8+T-cell-deficient mice bearing syngeneic LL2 tumor (Fig. 3).
Twenty-two days after tumor inoculation, wild- type mice receiving S. Choleraesuis had small
tumor volume than the control mice receiving PBS (50.46%, p<0.01) (Fig. 3a). By contrast, there
was a slight difference in mean tumor volumes in CD4+T-cell-deficient mice (P<0.05) (Fig. 3b).
The mean tumor volume in CD4+T-cell-deficient mice treated with S. Choleraesuis was lowered by
34.75% compared with that treated with PBS, but that was lowered by 42.15% in CD8+
T-cell-deficient mice (Fig. 3b and c). To further quantify tumor burden, the tumor weight was
measured. The wild- type mice receiving S. Choleraesuis significantly inhibited the growth of
tumor compared with the control mice receiving PBS (62.17%, p<0.01). The CD4+T-cell-deficient
mice treated with S. Choleraesuis had 26.47% less tumor weight compared with those treated with
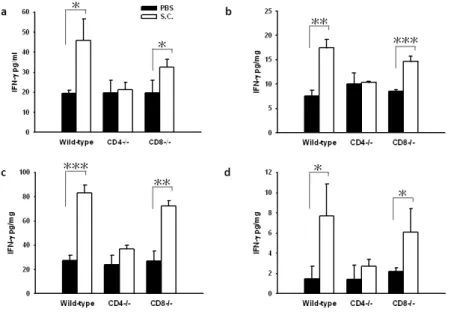
Effects of Salmonella on interferon-γinduction in vivo
Previously, we found that IFN-γplay an important role in antitumor activity of Salmonella
treatment (Lee et al. 2008b). To examine the role of T cell on cytokine induction by Salmonella,
wild-type and CD4+T-cell-deficient, and CD8+T cell-deficient mice were given an intravenous
injection of S. Choleraesuis. IFN-γlevelsof sera, spleens, liver, and tumors were measured (Fig. 4).
The level of IFN-γwasnotsignificantly changed in thesera,spleens,livers, and tumors derived
from mice treated with PBS. Compared with CD4+T-cell-deficient mice that received Salmonella,
the levels of IFN-γweresignificantly increased in thesera,spleens,livers, and tumors derived from
wild-type or CD8+T-cell-deficient mice treated with S. Choleraesuis. However, IFN-γproduction
induced by the bacteria was not observed in CD4+T-cell-deficient mice. Collectively, these findings
demonstrate that S. Choleraesuis elicited IFN-γfor the most part via CD4+T cell.
Increased infiltrating immune cells in the tumors following the treatment of Salmonella in
mice
The tumors from mice bearing the tumors treated with S. Choleraesuis were analyzed for cell
infiltrates by immunohistochemical staining. Representative results for immunohistochemistry are
were observed in the mice treated with S. Choleraesuis and, in particular, in wild-type mice. The
numbers of infiltrating immune cells in the tumors derived from CD8+T-cell-deficient mice treated
with S. Choleraesuis were significantly increased compared with those in the tumors derived from
CD4+T-cell-deficient mice treated with Salmonella, whereas no such difference was found in
control mice treated with PBS (Fig. 5). Taken together, these results indicate that S. Choleraesuis
increased more infiltrating immune cells in the tumors derived from CD8+T-cell-deficient mice
Discussion
The successful induction of immunity against poorly immunogenic malignancies is a major
challenge for cancer therapy. Previously, we have demonstrated that host immune responses
cooperate with bacteria-mediated tumor destruction during S. Choleraesuis treatment (Lee et al.
2004; Lee et al. 2005). The interactions between oncolytic S. Choleraesuis therapy and immune
mechanisms are likely to be complex. In the present study, we have identified one important
mechanism involved in the recruitment of effector immune cells. Previously, it was detected strong
IFN-γproduction in the tumors derived from mice after S. Choleraesuis treatment (Lee et al. 2008b).
Herein, we demonstrate that S. Choleraesuis induced IFN-γproduction and polarized theT-cell
response to a Th1-dominant state in wild-type mice, but not in CD4+T-cell-deficient mice. Thus,
bacteria-activated CD4+T cell infiltrating tumor may be a relevant source of IFN-γin the tumor
microenvironment and may contribute, a least in part, to the host antitumor immunity induced by S.
Choleraesuis. Meanwhile, the accumulation of Salmonella in tumor sites provoked a potent
inflammatory response, which recruited large numbers of immune cells. IFN-dependent chemokines,
such as IFN-inducible chemokines CXCL9 (MIG), and CXCL10 (IP-10) (Lee et al. 2008a; Lee et al.
2008b), induced by S. Choleraesuis are expected to recruit activated effector cells within the tumor.
Actuality, we found a large number of infiltrating immune cells such as macrophages, neutrophils
within tumor microenvironment. Antitumor effects of neutrophils, in particular, after being
activated by substances derived from microorganisms have also been demonstrated in various
cause tissue damage and this represents the basis for tumor destruction. In addition, macrophages
activated by bacterial products such as lipopolysaccharide (LPS) and Th1 cytokine are capable of
lysing tumor cells, expressing immunostimulatory cytokines, and presenting tumor-associated
antigens to recruit T cells (Luo et al. 2006). Recently, the CD8+T cell immune response induced by
Salmonella reported by Saccheri et al. was observed antimicrobial response present in tumors to
activate cytotoxic CD8+T cells that could recognize and kill tumor cells (Saccheri et al. 2010). In
our study, we also demonstrated that CD8+T cell is involved in the antitumor effector of S.
Choleraesuis. The current findings and our previous reports strongly suggest that Salmonella elicit
antitumor effects by stimulating host immune responses (Lee et al. 2004; Lee et al. 2008b; Saccheri
et al. 2010).
In CD8+T-cell deficient mice, we also observed the antitumor effect of S. Choleraesuis, the
slight production of IFN-γand the infiltrating immune cells in the tumors. These results implicate
that the antitumor effectors of S. Choleraesuis is partially mediated through CD4+T cell.
Furthermore, LPS from Salmonella may induce apoptosis of tumor and endothelial cells to enhance
the activity of antitumor (Bannerman et al. 2001). Meanwhile, the competition for nutrients between
tumor cells and Salmonella may also induce the suppression of tumor growth (Pawelek et al. 1997).
The mechanisms involved in the antitumor effects of S. Choleraesuis are likely to be multifaceted.
Thus, further work is warranted to elucidate the more underlying mechanism of antitumor effects of
S. Choleraesuis.
responsible for recruiting peripheral immune cells to the tumor in wild-type mice, but not in
T-cell-deficient mice. We suggest the T cell is involved in the regulation of Salmonella-induced host
antitumor immunity in tumor-bearing mice. Thus, this study may provide a cellular basis for
understanding the recruitment of effector immune cells and the synergism between the oncolytic
effect of S. Choleraesuis and adaptive antitumor immune mechanisms. It is possible to exploit an
immune response to infection to inhibit tumor growth by bypassing tumor immune evasion
mechanism (Saccheri et al. 2010). In phase I study, patients received VNP20009 that rapidly cleared
from blood, and most tumor were not detectable the colonization of Salmonella (Toso et al. 2002).
Patient had pre-existing anti-Salmonella antibodies and was not accompanied the colonization of
Salmonella in the tumor sites after systemically administration. In agreement with clinical study,
our previous results indicated that the higher anti-Salmonella antibody titers in the host cause fewer
amounts of Salmonella in the tumor sites (Lee et al. 2009). In this work described here, we suggest
that host T cells also play important roles in the tumor-reducing effect of Salmonella in
tumor-bearing mice. Previously, we demonstrated that S. Choleraesuis in combination with cisplatin
appears to hold promise for the treatment of solid tumors (Lee et al. 2005). The combination
therapy may facilitate the bacterial therapies with improve antitumor efficacy. Herein, this new
understanding of the mechanisms of antitumor activities of Salmonella in host will allow for the
References
Bannerman DD, Tupper JC, Ricketts WA Bennett CF, Winn RK, Harlan JM (2001) A constitutive
cytoprotective pathway protects endothelial cells from lipopolysaccharide-induced apoptosis.
J Biol Chem 276:14924-14932.
Engelbart K, Gericke D (1964) Oncollysis by Clostrida. V. transolaten tumors of the hamster.
Cancer Res 24:399-342.
Forbes NS, Munn LL, Fukumura D, Jain RK (2003) Sparse initial entrapment of systemically
injected Salmonella typhimurium leads to heterogeneous accumulation within tumors. Cancer
Res 63:5188-5193.
Fujii Y, Kimura S, Arai S, Sendo F (1987) In vivo antitumor effect of lymphokine-activated rodent
polymorphonuclear leukocytes. Cancer Res 47:6000-6005.
Kasai S, Fujimoto S, Nitta K, Baba H, Kunimoto T (1991) Antitumor activity of polymorphonuclear leukocytesactivated by a β-1,3-D-glucan. J Pharmacobiodyn 14:519-525.
Lee CH, Wu CL, Chen SH, Shiau AL (2009) Humoral immune responses inhibit the antitumor
activities mediated by Salmonella enterica serovar Choleraesuis. J Immunother 32:376-388.
Lee CH, Wu CL, Shiau AL (2004) Endostatin gene therapy delivered by Salmonella choleraesuis in
murine tumor models. J Gene Med 6:1382-1393.
Lee CH, Wu CL, Shiau AL (2008a) Salmonella choleraesuis as an anticancer agent in a syngeneic
model of orthotopic hepatocellular carcinoma. Int J Cancer 122(4):930-935.
induced by Salmonella choleraesuis. Clin Cancer Res 14:1905-1912.
Lee CH, Wu CL, Tai YS, Shiau AL (2005) Systemic administration of attenuated Salmonella
choleraesuis in combination with cisplatin for cancer therapy. Mol Ther 11:707-716.
Low KB, Ittensohn M, Le T, Platt J, Sodi S, Amoss M, Ash O, Carmichael E, Chakraborty A,
Fischer J, Lin SL, Luo X, Miller SI, Zheng L, King I, Pawelek JM, Bermudes D (1999) Lipid
A mutant Salmonella with suppressed virulence and TNF-αinduction retain tumor-targeting
in vivo. Nat Biotechnol 17:37-41.
Luo Y, Zhou H, Krueger J, Kaplan C, Lee SH, Dolman C, Markowitz D, Wu W, Liu C, Reisfeld RA,
Xiang R (2006) Targeting tumor-associated macrophages as a novel strategy against breast
cancer. J Clin Invest 116:2132-2141.
Pawelek JM, Low KB, Bermudes D (1997) Tumor-targeted Salmonella as a novel anticancer vector.
Cancer Res 57:4537-4544.
Pawelek JM, Low KB, Bermudes D (2003) Bacteria as tumour-targeting vectors. Lancet Oncol
4:548-556.
Saccheri F, Pozzi C, Avogadri F, Barozzi S, Faretta M, Fusi P, Rescigno M (2010)
Bacteria-induced gap junctions in tumors favor antigen cross-presentation and antitumor
immunity. Sci Transl Med 2:44ra57.
Shiau AL, Chen YL, Liao CY, Huang YS, Wu CL (2001)Prothymosinαenhancesprotective
immune responses induced by oral DNA vaccination against pseudorabies delivered by
Thiele EH, Arison RN, Boxer GE (1964) Oncolysis by clostridia. IV. Effect of nonpathogenic
clostridial spores in normal and pathological tissues. Cancer Res 24:234-238.
Toso JF, Gill VJ, Hwu P, Marincola FM, Restifo NP, Schwartzentruber DJ, Sherry RM, Topalian SL,
Yang JC, Stock F, Freezer LJ, Morton KE, Seipp C, Haworth L, Mavroukakis S, White D,
MacDonald S, Mao J, Sznol M, Rosenberg SA (2002) Phase I study of the intravenous
administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J
Clin Onco 20:142-152.
Yang KD, Stone RM, Lee CS Chao TY, Cheng SN, Shaio MF (1992) Effect of picibanil (OK432)
on neutrophil-mediated antitumor activity: implication of monocyte-derived
neutrophil-activating factors. Cancer Immunol Immunother 35:277-282.
Yu YA, Shabahang S, Timiryasova TM, Zhang Q, Beltz R, Gentschev I, Goebel W, Szalay AA
(2004) Visualization of tumors and metastases in live animals with bacteria and vaccinia virus
FIGURE LEGEND
Fig. 1. Increases in CD4+T- and CD8+T-cell infiltrates in the tumors from LL2 tumor-bearing mice
treated with S. Choleraesuis (S.C.). Groups of 4 C57BL/6 mice that had been inoculated s.c. with
LL2 cells (106) at day 0 were treated i.v. with S. Choleraesuis (2 × 106cfu ) at day 7. Vehicle control
mice received PBS. A, Tumors were excised at day 20 and immunostained with antibodies against
CD4+or CD8+(× 200). B, CD4+and CD8+
averaging the cell numbers from three fields of highest positive-stained cell density at × 200
magnification in each section (mean ± SEM, n=4).*
,
P<0.05; **,
P<0.01. Each experiment wasrepeated three times with similar results.
Fig. 2. The spatial and temporal distribution of S. Choleraesuis (S.C.) in T-cell-deficient and
wild-type mice. The mice bearing LL2 tumors ranging from 50 to100 mm3were injected i.v. with S.
Choleraesuis (2 × 106cfu), and the amounts of S. Choleraesuis in the (A) tumor, (B) livers, and (C)
repeated three times with similar results.
Fig. 3. Antitumor effects of S. Choleraesuis (S.C.) on tumor growth in T-cell-deficient and
wild-type mice bearing LL2 tumors. A, Groups of 7 mice that had been inoculated s.c. with LL2
cells (106) at day 0 were treated i.v. with S. Choleraesuis (2 × 106cfu) or PBS at day 7. B, Groups of
7 CD4+T-cell-deficient mice that had been inoculated s.c. with LL2 cells (106) at day 0 were treated
i.v. with S. Choleraesuis (2 × 106cfu) or PBS at day 7. (C) Groups of 6-7 CD8+T-cell-deficient
mice that had been inoculated s.c. with LL2 cells (106) at day 0 were treated i.v. with S.
Choleraesuis (2 × 106cfu) or PBS at day 7.Tumor volumes among different treatment groups were
compared at day 22. (D) Groups of 3-4 mice that had been inoculated s.c. with LL2 cells (106) at
day 0 were treated i.v. with S. Choleraesuis (2 × 106cfu) or PBS at day 7. The tumor weight was
measured at day 10 postinfection.
Fig. 4. Effects of S. Choleraesuis (S.C.) on cytokine induction in T-cell-deficient and wild-type
mice bearing LL2 tumors. The mice bearing LL2 tumors injected i.v. with S. Choleraesuis (2 × 106
cfu). IFN-γlevelsin the(A) sera, (B) liver, (C) spleens, and (D) tumors were measured by ELISA at
12 h postinfection (mean ± SD, n = 3-4). *
,
P<0.05; **,
P<0.01; ***,
P<0.001. EachFig. 5. Increase of infiltrating cells in the tumors from T-cell-deficient mice treated with S.
Choleraesuis (S.C.). Mice bearing LL2 tumors at day 0 were injected i.v. with 2 × 106cfu of S.
Choleraesuis or with PBS at day 7. A, Tumors were excised at day 20, immunostained with
antibodies against Gr-1, and Mac-3 were used to detect infiltrating cells (× 200). (B) Neutrophils,
and (C) macrophages cells that infiltrated tumors were determined by averaging the cell numbers
from three fields of highest positive-stained cell density at × 200 magnification in each section
(mean ± SEM, n =4). **