Cd induced changes in proline level and peroxidase activity in roots of rice
seedlings
Sheng Lee Chen & Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China (*authorfor correspondence)
Received 4 October 1994; accepted 5 February 1995
Key words: cadmium, Oryza sativa, peroxidase, proline, root growth
Abstract
The effects of Cd on changes in proline level and peroxidase activity in roots of rice seedlings were investigated. CdClz was effective in inhibiting root growth and in accumulating proline in roots. The inhibition of root growth by Cd is reversible. The reduction of root growth induced by Cd is closely associated with accumulation of proline in roots. External application of proline markedly inhibited root growth of rice seedlings in the absence of Cd. Ionically bound, but not soluble, peroxidase activity in roots was increased by CdClz. Proline treatment also resulted in an increase in ionically bound peroxidase activity in roots. The relationship between growth inhibition of roots induced by Cd and changes of proline level and peroxidase activity is discussed.
Abbreviations: POX - peroxidase.
1. Introduction
Proline has been shown to accumulate in tissue/organs of plants subjected to drought-, salt- or temperature stress and/or infected by some pathogens [ 1, 3, 151, exposed to gases such as sulphur dioxide, ammonia or nitrogen dioxide [2]. Recently, Saradhi and Saradhi [17] provided evidence to show that Cd was the strongest inducer, among 4 tested heavy metals, for proline accumulation. Cd-induced increase in peroxi- dase (POX) activities has previously been reported for soybean and bean leaves [9, 131.
There are reports that proline accumulation is corre- lated with growth inhibition induced by NaCl [S]. It is not known whether such correlation exists in Cd- stressed plants. Growth inhibition may result from cell wall tightening process related to formation of cross-linkages among cell wall polymers by POX [8]. The ionically bound fraction of POX, removable from homogenized tissues with high ionic strength buffers, has been generally accepted to represent cell wall activity, and several studies have reported an associa- tion between increase in ionically bound POX activity
and the timing or location of cessation of cell expan- sion [ 10, 1 1,141. No information is available about the correlation between ionically bound POX activity and plant growth under Cd stress. The present investigation was designed to study the changes in proline level and POX activity in Cd-stressed roots of rice seedlings and their correlation with root growth.
2. Materials and methods
Rice (Oryza sativa L., cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri dish (20 cm) con- taining distilled water at 37 ‘C under dark condition. After 1 -day incubation, uniformly germinated seeds were selected and transferred to Petri dishes (9.0 cm) containing two sheets of Whatman No. 1 filter paper moistened with 10 mL of distilled water or test solu- tions. Each Petri dish contained 20 germinated seeds. Each treatment was replicated 4 times. The germinated seeds were allowed to grow at 27 “C in darkness and
3 mL of distilled water or test solutions was added to each Petri dish on day 3 of the growth. Fresh weight of roots was measured at the times indicated.
Proline was extracted and its concentration deter- mined by the method of Bates et al. [4]. Roots were homogenized with 3% sulfosalicylic acid and the homogenate was centrifuged. The supernatant was treated with acetic acid and acid ninhydrin, boiled for 1 h and then absorbance at 520 nm was deter- mined. Levels of proline are express as pm01 per g fresh weight.
POX was extracted according to the method described by MacAdam et al. [ 141. Briefly, roots were frozen in liquid N and ground with a mortar and pestle and mixed with either 0.05 M potassium phosphate buffer (pH 5.8) to extract soluble POX, or with the same buffer containing 0.8 M KC1 to extract both sol- uble and ionically bound (“total”) POX. POX activity was measured using a modification of the procedure described by Curtis [6]. The assay medium contained 0.05 M potassium phosphate buffer (pH 5.8), 7.2 mM guaiacol, 11.8 mM Hz02 and 0.1 mL enzyme extract in a final assay volume of 3.0 mL. The reaction was initiated by the addition of Hz02 and the change in absorbance at 470 nm was measured. Activity was calculated using the extinction coefficient (26.6 mM-’ cm- ’ at 470 nm) for tetraguaiacol. One unit of POX was defined as the amount of enzyme that caused the formation 1 pm01 tetraguaiacol per min.
3. Results
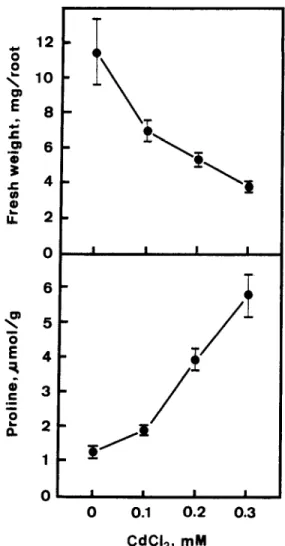
Root growth was tracked by measuring fresh weight of roots. Figure 1 shows the effects of CdClz on growth of and proline levels in roots of rice seedlings. Increasing concentrations of CdClz from 0.1 to 0.3 mM progressively decreased root growth and increased proline level in roots. The reduction of root growth is closely associated with the accumulation of proline in roots.
To characterize further the role of proline accumu- lation on Cd-induced inhibition of root growth, seeds were germinated in CdClz solution (0.3 mM) for 3 days and, after this, the seedlings were incubated in distilled water and CdClz (0.3 mM), respectively, for another 3 days. When 3-day-old Cd-treated seedlings were remained in CdClz, only slight root growth was observed (Fig. 2). However, when seedlings were transferred to distilled water, root growth rapidly resumed and increased linearly with increasing dura-
I I I I
0 0.1 0.2 0.3 Cd&, mM
Fig. 1. Effects of CdClz on root growth and proline level in roots of rice seedlings. Root growth and proline level were measured after 5 days of treatment. Vertical bars represent standard errors (n = 4).
tion of incubation (Fig. 2). Figure 2 also shows that proline level in roots of seedlings remained in CdClz solution is higher than that of seedlings transferred to distilled water.
If proline accumulation is important in regulating root growth, then exogenous application of proline is expected to increase proline level in roots and to inhibit root growth of rice seedlings. As indicated in Fig. 3, this is indeed the case. Increasing concentrations of external proline from 2.5 to 10 m&f progressively increased endogenous proline level and decreased root growth.
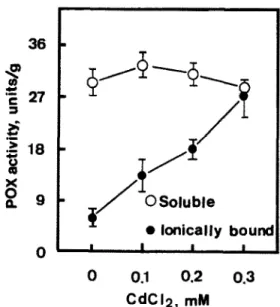
Figure 4 shows the effects of CdClz on soluble and ionically bound POX activities in roots of rice seedlings. CdCl2 had no effect on soluble POX activity
0 CdCI,- Hz0
I 1 I I
3 4 5 6
Days
Fig. 2. Changes of fresh weight of and proline levels in roots of Cd-pretreated rice seedlings grown in the presence or absence of CdCla (0.3 m&f). Rice seeds were germinated in CdCi2 (0.3 m&f) for 3 days and then the seedlings were incubated in distilled water (o) and CdClz (e), respectively. Vertical bars represent standard errors (n = 4).
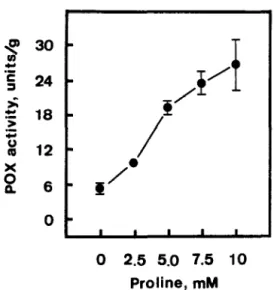
in roots. However, increasing concentrations of CdClz progressively increased ionically bound POX activity. Similar to the effect of CdCl2, proline treatment also resulted in an increase in POX activity in roots (Fig. 3.
4. Discussion
In rice, proline accumulation has been demonstrated in water-stressed and senescing detached leaves [12, 181 and in NaCl-stressed seedling roots [16]. The present investigation shows that proline accumulation
0 Proline
l Root growth
Proiine, mM
Fig. 3. Effects of prohne on root growth and proline level in roots of rice seedlings. Root growth and proline level was measured after 5 days of treatment. Vertical bars represent standard errors (n = 4).
l Ionically bow
I I I i
0 0.1 0.2 0.3 CdCl2, mM
Fig. 4. Effects of CdClz on soluble and ionicafly bound peroxidase (POX) activities in roots of rice seedlings. Enzyme activities were measured after 5 days of treatments. Vertical bars represent standard errors (n = 4).
can be induced in roots by treating seedling with Cd. The ability of Cd to accumulate proline has also been observed in detached rice leaves (unpubl. data). Our results are in general consistent with those reported by Saradbi and Saradbi [17], who showed that heavy
OU
0 2.5 5.0 7.5 10 Proline, mM
Fig. 5. Effects of proline on ionically bound peroxidase (POX) activity in roots of rice seedlings. Enzyme activities were measured after 5 days of treatments. Vertical bars represent standard errors (n = 4).
metals including Cd, can induce proline accumula- tion.
The present investigation also shows that proline accumulation is closely associated with growth inhibi- tion induced by Cd. It seems that Cd-induced inhibition of root growth is a consequence of proline accumula- tion, since Cd-induced inhibition of root growth was produced in rice seedlings fed with proline. When rice seedlings pretreated with Cd were transferred to distilled water, root growth resumed and proline level in roots declined. These results suggest that Cd- induced inhibition of root growth is not irreversible and further supports the close link between proline accumulation and growth inhibition of roots induced by Cd. Proline accumulation induced by NaCl has also been demonstrated to be correlated with growth inhi- bition of Brussicu napus callus [5].
Cd-induced increase in soluble and ionically-bound POX has previously been reported for bean leaves [9]. However, we showed that ionically bound, but not soluble, POX activity in roots was increased by Cd treatment. Ionically bound POX has been equated with the cell wall fraction because it is the activity that remains associated with cell walls of homogenized tissue until the tissue is extracted with high ionic strength buffers. It has been postulated by several investigators that the action of POX located in cell walls would be to confer rigidity to the cell wall and prevent later expansion involved in growth [7, 8, 10,
11,141. Thus, Cd-induced inhibition in root growth of rice seedlings is likely to result from cell wall tight- ening processes related to formation of cross-linkages among cell wall polymers.
Of particular interesting is the finding that proline resulted in an increase in ionically bound POX activity in roots of rice seedlings in the absence of Cd. These results, again, support our suggestion that Cd-induced proline accumulation is associated with growth inhi- bition of roots. However, more studies are required to confirm the control of ionically bound POX activity by proline, in particular focusing on the location of proline in cell walls.
Proline accumulation in plants under stress condi- tions has been suggested to play several roles [3]. Since growth is generally thought to be an energy-required process, elevated proline level induced by Cd is most likely acting as a way to save energy by inhibiting root growth, possibly through the action of POX located in cell walls, and as a readily utilizable source of energy and amino groups once Cd-stress is relieved.
Acknowledgement
This work was supported by a research grant from the National Science Council of the Republic of China.
References 1. 2. 3. 4. 5. 6. 7.
Amberger-Ochsenbauer S and Odbendorfer J (1988) Levels of free proline in ornamental plants. I. Influence of plant age and leaf region in .S&nti pauliu and Chrysanthemum. J Plant Physiol 132: 758-761
Anbazghan M, Krishnamurthy R and Bhagwat KA (1988) Proline: an enigmatic indicator of air pollution tolerance in rice cultivars. J Plant Physiol 133: 122-123
Aspinall D and Paleg LG (198 1) Proline accumulation: phys- iological aspects. In: Paleg LG and Aspinall D (eds) The Physiology and Biochemistry of Drought Resistance in Plants, pp 280-295. Sydney: Academic Press
Bates LS, Waldren RP and Teare ID (1973) Rapid determi- nation of free proline for water stress studies. Plant Soil 39: 205-207
Chandler SF and Thorpe TA (1987) Characterization of growth, water relations, and proline accumulation in sodium sulfate tolerant callus of Brnssica napusl. cv Westar (Canola). Plant Physiol84: 106-I 11
Curtis CR (197 1) Disc electrophorectic comparisons of pro- teins and peroxidases from Phaseolus vulgaris leaves infected with Agrobacterium tumefaciens. Can J Bot 49: 333-337 Fry SC (1979) Phenolic components of the primary cell wall and their possible role in the hormonal regulation of growth. Planta 146: 343-35 1
8. Fry SC (1986) Cross-linking of matrix polymersin the growing cell walls of angiosperms. Annu Rev Plant Physiol 37: 165- 186
9. Fuhrer J (1982) Early effects of excess cadmium uptake in
Phaseolus vulgaris. Plant Cell Environ 5: 263-270
10. Gardiner MG and Cleland R (1974) Peroxidase changes dur-
ing the cessation of elongation in Pisum safivum stems. Phy-
tochemistry 13: 1095-1098
11. Goldberg R, Imberty A and Chu-Ba J (1986) Development of isoperoxidases along the growth gradient in the mung bean hypocotyl. Phytochemistry 25: 1271-1274
12. Kao CH (1981) Senescence of rice leaves VI. Comparative study of the metabolic changes of senescing and turgid and water-stressed excised leaves. Plant Cell Physio122: 683-688 13. Lee KC, Cunningham BA, Paulsen GM, Liang GA and Moore RB (1976)Effects of cadmium on respiration rate and activities of several enzymes in soybean seedlings. Physiol Plant 36: 4-6
14. MacAdam JW, Nelson CJ and Sharp RE (1992) Peroxidase activity in the leaf elongation zone of tall fescue. I. Spatial distribution of ionically bound peroxidase activity in genotypes differing in length of the elongation zone. Plant Physiol 99:
872-878
15. NikolopoulosD andManetasY (1981)Compatiblesolutes and in vitro stability of Salsola soda enzymes: Prohne incompati- bility. Phytochemistry 30: 411-413
16. Roy D, Bhunia A, Basu N and Banerjee SK (1992) Effect of NaCl-salinity on metabolism of proline in salt-sensitive and salt-resistant cultivars of rice. Biol Plant 34: 159-162 17. Saradhi A and Saradhi PP (1981) Proline accumulation under
heavy metal stress. J Plant Physiol 138: 554-5.58
18. Wang CY, Cheng SH and Kao CH (1982) Senescence of rice leaves VII. Proline accumulation in senescing excised leaves. Plant Physio169: 1348- 1349