Reusable Amperometric Biosensor for Measuring Protein Tyrosine
Kinase Activity
Chung-Liang Wang,
†Lan-Yi Wei,
†Chiun-Jye Yuan,*
,†,‡and Kuo Chu Hwang
§†Department of Biological Science and Technology and‡Institute of Molecular Medicine and Bioengineering, National Chiao Tung
University, Hsinchu, Taiwan, ROC,
§Department of Chemistry, National Tsing Hua University, Hsinchu, Taiwan, ROC
ABSTRACT: This work presents a simple, low-cost and reusable label-free method for detecting protein tyrosine kinase activity using a tyrosinase-based amperometric biosensor (tyrosine kinase biosensor). This method is based on the observation that phosphorylation can block the tyrosinase-catalyzed oxidation of tyrosine or tyrosyl residue in peptides. Therefore, the activity of p60c-src protein tyrosine kinase (Src) on the developed tyrosine kinase biosensor could be quickly determined when its specific peptide substrate, p60c-src substrate I, was used. The tyrosine kinase biosensor was highly sensitive to the activity of Src with a linear dynamic range of 1.9−237.6 U/mL and the lowest detection limit of 0.23 U/mL. Interestingly, the tyrosine kinase activity can be
measured using the developed tyrosine kinase biosensor repetitively without regeneration. The inhibitory effect of various kinase inhibitors on the Src activity could be determined on the tyrosine kinase biosensor. Src-specific inhibitors, PP2 and Src inhibitor I, effectively suppressed Src activity, whereas PD153035, an inhibitor of the epidermal growth factor receptor, was ineffective. Staurosporine, a universal kinase inhibitor, inhibited Src activity in an ATP concentration-dependent manner. These results suggests that the activities of tyrosine kinases and their behaviors toward various reagents can be effectively measured using the developed tyrosine kinase biosensor.
P
rotein kinase is an enzyme that catalyzes the phosphor-ylation of the serine, threonine, or tyrosine residues in proteins by transferring the γ-phosphate from an adenosine-triphosphate (ATP) molecule. At least 518 putative protein kinase genes have been identified in human genome1and can be grouped into two main classes, protein serine/threonine kinases and protein tyrosine kinases.2 Among protein kinases identified, around 90 protein tyrosine kinases are identified and found only in Metazoans.1 Protein tyrosine kinases regulate many cellular important events, such as embryo development, immune responses, cell proliferation, differentiation, and programmed cell death.1,3,4 The dysregulation of protein tyrosine kinases may results in cancer, diabetes, immune diseases, and cardiac diseases.5−8Since the activity of protein tyrosine kinases in biological systems may be indicative of the physiological and/or pathophysiological conditions of cells, these enzymes represent attractive targets for drug screen-ing.9−12 Additionally, information about the activity profile of protein tyrosine kinases may be useful for the development of effective therapeutic protocols for diseases.Various approaches for detecting protein phosphorylation have been developed. Conventionally, the32P radioisotope is
used to label peptides and proteins in solution13,14 or on an array chip.15,16 Although these methods sensitively detect phospho-peptides and -proteins in vivo and in vitro, they depend on specific instruments and a skillful technician, and suffer from a high risk of radioactive contamination. To
eliminate the problems that are associated with radioactive methods, the optical method17−19 the fluorescence-based method,20−22 phospho-specific antibodies,23,24 the metal-chelating method25,26 and mass spectrometry27,28 have been developed. However, these methods have the disadvantages of being expensive, suffering from the high background, being time-consuming, requiring tedious sample handling and/or sophisticated operating procedures.
Electrochemical methods for determining the activity of protein kinases in vitro29−37 have been demonstrated to be simpler and more cost-effective than previous methods. However, one of the shortcomings of these methods is the need for labor-intensive postlabeling procedures owing to the use of ATP analogs contain biotin or thiol conjugate as the cosubstrate of the kinase reactions. Although the use of adenosine-5′-[γ-ferrocene] triphosphate simplifies the electro-chemical sensing process34,38−40 and allows the structural and electronic factors surrounding the kinase-catalyzed reactions to be explored electrochemically, the immobilization of the peptide substrate on the electrode surface makes these electrochemical sensors difficult to repetitively measure the protein kinase activity. Although using alkaline phosphatase to
Received: September 6, 2011 Accepted: December 21, 2011 Published: December 21, 2011
remove the phosphate sites and regenerate phosphate-free peptide can solve this problem,41 this procedure costs and involves tedious handling process. Therefore, the use of these electrochemical methods to fabricate a high-throughput sensing system for systematic large-scale measurements of the kinase activity is limited.
This work presents a simple, sensitive, reusable and label-free amperometric biosensor for measuring the activity of protein tyrosine kinases. This approach is based on the hypothesis that tyrosinase, a binuclear copper containing enzyme,42 can catalyze the oxidation of tyrosine (Tyr) to L-DOPA (L-3,4-dihydroxyphenylalanine) and L-DOPA-quinone (Scheme 1A).
Subsequently, the L-DOPA-quinone is reduced to L-DOPA under a working potential of−0.2 V. Interestingly, initiation of the oxidation reaction by tyrosinase depends on the hydroxyl group in the phenolic compounds.43Phospho-tyrosine (pTyr) is reasonably assumed to be unrecognizable by tyrosinase. Therefore, tyrosine kinase activity can be simply determined by measuring the tyrosyl residues that remain in peptides following kinase reactions. p60c-src Protein tyrosine kinase (Src), a gene product of proto-oncogene c-src, was used as a model to prove the feasibility of the hypothesis. Src, originally identified as the mammalian homologue of viral v-src gene from Rous sarcoma virus that causes viral-sarcoma, phosphorylates tyrosyl residues in protein substrates.44The development of cancers of the lung, prostate, pancreas, breast, and colon has been associated with the overexpression and aberrant activation of Src.45−47 The
proposed amperometric biosensor was proved herein to detect effectively and repetitively the Src-catalyzed peptide phosphor-ylation. The inhibitory profile and potency of various protein kinase inhibitors on developed electrochemical sensing system were also analyzed.
■
MATERIALS AND METHODSReagents. Carbon fiber paper TGP-H-060 (thickness: 0.17 cm) was purchased from Toray Ind. (Japan). Teflon glue was supplied by DuPont (USA). The recombinant Src (1.16× 105
U/mg) was bought from GeneScript (USA). Tyrosinase (EC 1.14.18.1, 5370 U/mg from mushroom), Tyr, pTyr, dipeptides, Gly-Tyr and Tyr-Ala, Src inhibitor-1 and PP2 were bought from Sigma-Aldrich. Poly vinyl alcohol functionalized with pyridinium methyl sulfate (PVA-SbQ) was bought from Toyo Gosei Kogyo Co. Ltd. (Japan). The putative peptide substrate for Src, NH3-Tyr-Ile-Tyr-Gly-Ser-Phe-Lys-COOH or YIYGSFK
(p60c-src substrate I), was synthesis by GL Biochem Ltd. (Shanghai, China). ATP was from BioVision. PD153035 hydrochloride, an epidermal growth factor receptor (EGFR) inhibitor, was purchased from Tocris Bioscience (UK). Staurosporine was obtained from Calbiochem (U.S.A.). Fetal bovine serum (FBS), Dulbecco’s modified eagle medium (DMEM), penicillin G, streptomycin, and lipofectamine were obtained from Life Technologies. All other reagents were reagent grade.
Nitrogen Plasma Treatment. The CFP was pretreated with nitrogen (N2) plasma that was jetted out of a nozzle directly onto the surface of the CFP under the atmospheric pressure. The nozzle was situated 1 cm above the surface of CFP. The power of the plasma was set between 253 V and 260 V with a current of 2.3 Amp. The scan rate was set at 5 cm s−1 with a scan length of 0.3 cm.
Preparation of the Enzyme Electrode. The working electrode was prepared as described previously.48 PVA-SbQ/ tyrosinase mixture was first prepared by mixing 16% PVA-SbQ stock solution with tyrosinase stock solution (50 UnitsμL−1) in a 1:1 ratio (v/v). PVA-SbQ/tyrosinase mixture (2.5 μL) was spread on the surface of the plasma-pretreated CFP electrode. The PVA-SbQ/tyrosinase/CFP electrode was then sealed in a box containing saturated glutaraldehyde vapor at 4°C for 12 h to allow cross-linking reactions to occur. Photopolymerization of PVA-SbQ was achieved by placing the enzyme electrode on ice under an illuminating light for 2 h. The enzyme electrode was then washed once with phosphate buffer saline (PBS buffer) (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.76 mM KH2PO4, pH 6.8) prior to storage in PBS buffer at 4°C
overnight before use.
Electrochemical Measurement. All experiments were carried out with a conventional three-electrode system with the enzyme electrode as the working electrode and a standard platinum electrode as the counter electrode. An Ag/AgCl electrode was used as a reference. Input and output signals from the potentiostat CHI 440 (CH Instruments, West Lafayette, IN, U.S.A.) were coupled to a PC. Amperometric measure-ments of the developed biosensors were carried out in a reaction chamber containing 10 mL PBS buffer. The working potential was maintained at a fixed working potential of−0.2 V vs Ag/AgCl for amperometric measurements. Unless otherwise stated, all measurements were carried out under the room temperature. The solution in the reaction chamber was constantly magnetic stirred during the reaction.
Scheme 1. (A) Working Model for the Determination of the Activity of Protein Tyrosine Kinases and (B) Schematic Illustration of the Principal for Tryosinase-Based Detection of the Phosphorylation of Peptidea
aIn a test tube, the kinase reaction is first performed in the presence of
peptide substrate and ATP. An aliquot of reaction mixture is then subjected to the electrochemical measurement of tyrosyl residue in the peptide. The phosphorylation prevents the occurrence of tyrosinase-based reductive current response for tyrosyl residue on the biosensor. In the presence of a kinase inhibitor, the phosphorylation is suppressed and the tyrosinase-based reductive current response for tyrosyl residue can be detected.
Tyrosine Kinase Activity Assay. Src stock solution (1.65 μM) was kept in the preserving buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.25 mM DTT, 0.1 mM EGTA, and 30% glycerol) at −80 °C until use. Before reaction, Src stock solution was diluted to the concentrations indicated in each experiment with the reaction buffer (50 mM Tris-HCl, pH 7.5, 2.8 mM MnCl2, 30 mM MgCl2and 137 mM NaCl). The p60c-src substrate I, a heptapeptide containing two tyrosyl residues, was dissolved in the reaction buffer to make a stock solution of 4.35 mM. The Src kinase reaction was carried out in a reaction buffer containing 4.22 nM Src, 3 mM p60c-src substrate I and 5 mM ATP at 30 °C for 30 min or as indicated in each experiment. Following kinase reaction, 5μL of 1 M acetic acid was added into the reaction mixture to stop the kinase reaction. Subsequently, an aliquot of kinase reaction mixture (20 μL) was subjected to the electrochemical measurements.
Cell Culture and Treatment. Human cervical cancer cell line (HeLa) was maintained in DMEM containing 10% FBS and 100 U/mL penicillin and 100μg/mL streptomycin. Cells were incubated in a humidified 37 °C incubation chamber containing 5% CO2. Epidermal growth factor (EGF) treatment was performed by incubating cells with 100 ng/mL EGF at 37 °C for 60 min. After treatment, cell lysates were prepared by incubating HeLa cells in a 100 mm culture plate with RIPA buffer (1% Triton X-100, 0.1% SDS, 50 mM Tris, pH 8.0, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Na-deoxycholate, 5 μg/mL leupeptin, and 5 μg/mL aprotinin) at room temper-ature for 15 min. Cell lysate was collected by scraping. The particulates were removed from cell lysates by centrifugation at 4°C and 14000 × rpm for 5 min.
■
RESULTS AND DISCUSSIONResponses of the Biosensor to Tyrosine and Phospho-tyrosine. Scheme 1B illustrates that tyrosine kinase assay procedures of the proposed tyrosinase-based ampero-metric biosensor48 (Tyr kinase biosensor). The phosphor-ylation reaction of substrate peptide by Src is first performed in a test tube. Phosphorylation of the tyrosyl residue in peptide may suppress the tyrosinase-based electrochemical responses as shown in Scheme 1A. A strong electrochemical response may occur when the activity of the tyrosine kinase is suppressed by inhibitor.
To prove the principle of the developed biosensor the ability of tyrosinase to distinguish tyrosine (Tyr) from phospho-tyrosine (pTyr) is first investigated. Although tyrosinase exhibits broad substrate specificity toward various phenolic compounds, its reactivity with phosphorylated tyrosine or tyrosyl residue is currently unclear. Therefore, the ability of the tyrosinase-based amperometric biosensor to distinguish Tyr from pTyr was studied herein. The Tyr kinase biosensor responded linearly to Tyr in a concentration range from 0−10 μM (R2= 0.997) with a sensitivity of 2.21 ± 0.04 μA μM−1
cm−2(Figure 1A, closed circle). The amperometric responses of Tyr and pTyr mixtures were also determined to study the effect of pTyr on the measurement of Tyr. Interestingly, a linear response to Tyr/pTyr mixtures at concentration ratios of 0:10, 2:8, 4:6, 6:4, 8:2, and 10:0 (μM/μM) (R2 = 0.998) was observed with a sensitivity of 2.13± 0.05 μA (μM Tyr)−1cm−2 (Figure 1A, open circle). The responses of the Tyr kinase biosensor to Tyr alone and to the Tyr/pTyr mixture were strongly correlated, with a correlation coefficient (r) of 0.996 (Figure 1B), suggesting that pTyr neither is recognized by the Tyr kinase biosensor nor interferes with the tyrosinase
catalyzed tyrosine oxidation, even at a concentration of 10 μM. These characteristics are important to the use of the developed Tyr kinase biosensor to measure the activity of Src, the reaction of which yields phospho-tyrosyl residue as the major product.
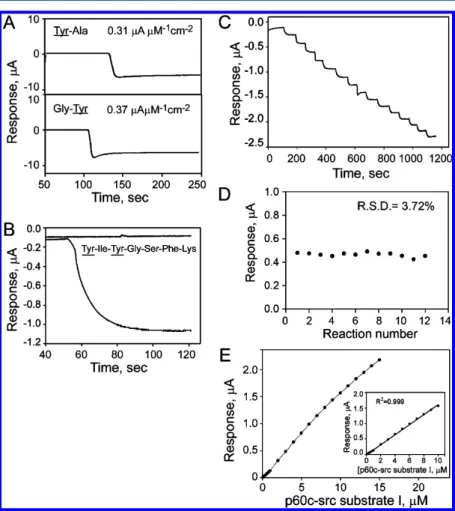
Responses of Tyr Kinase Biosensor to Tyrosyl-Containing Peptides. The electrocatalytic activity of Tyr kinase biosensor to tyrosyl residues in peptides was further studied using tyrosyl residue-containing dipeptides (Gly-Tyr and Tyr-Ala) and p60c-src substrate I, which is a heptapeptide (NH3-Tyr-Ile-Tyr-Gly-Ser-Phe-Lys-COOH) containing two tyrosyl residues, as models. The p60c-src substrate I has been demonstrated to be a specific and efficient substrate for Src with a Kmvalue of 55μM.
49
Two dipeptides were used herein because of their bulk group (such as an amino acid residue) on either the amino or the carboxyl side of the tyrosyl residue. These bulk groups may limit the access of tyrosyl residue to the immobilized tyrosinase on the CFP electrode. Interestingly, the Tyr kinase biosensor recognized the tyrosyl residue in both dipeptides, with a sensitivity of 0.37 and 0.31μA μM−1cm−2for Gly-Tyr and Tyr-Ala, respectively (Figure 2A). The response of the Tyr kinase biosensor to the tyrosyl residue in peptides was around 84% lower than that of free Tyr. The amino acids that surround the tyrosyl residue in a peptide may have a steric effect, partially blocking the accessibility of the tyrosyl residue to the active site of tyrosinase.
The feasibility of using the Tyr kinase biosensor to recognize the tyrosyl residues in the p60c-src substrate I was studied. As shown in Figure 2B, the Tyr kinase biosensor recognized p60c-src substrate I as a substrate with a sensitivity of 0.83 μA
Figure 1.Electrochemical responses of Tyr kinase biosensor to Tyr and pTyr. (A) Measured electrochemical responses of tyrosine kinase biosensor to various concentrations (0, 2, 4, 6, 8, and 10 μM) of tyrosine (●) or ratios (0:10, 2:8, 4:6, 6:4, 8:2, and 10:0μM/μM) of Tyr and pTyr (○). (B) Plot of correlation of electrochemical response of Tyr kinase biosensor to Tyr and Tyr/pTyr mixture.
μM−1cm−2. A steady-state baseline current was reached shortly
after each addition of the peptide solution, with a response time (95% of the steady-state current) of 25 s. Although the reactivity of tyrosinase to each of two tyrosyl residues is unknow, the equal distribution is postulated owing to the observation that the developed Tyr kinase biosensor exhibited a response to p60c-src substrate I, a heptapeptide, twice as that it did to the dipeptides. For each tyrosyl residue in the p60c-src substrate I, the sensitivity was around 0.42 μA (μM tyrosyl residue)−1 cm−2, which was similar to that of the developed biosensor to dipeptides. These results indicate that the Tyr kinase biosensor recognized tyrosyl residue(s) in the heptapeptide. However, an attempt to detect the tyrosyl residues in full length protein was unsuccessful (data not shown). A typical current−time (i-t) plot of the Tyr kinase biosensor was obtained in a PBS buffer by successively adding portions of p60c-src substrate I (Figure 2C). The capability of the developed Tyr kinase biosensor to repetitively measure the p60c-src substrate I was also performed (Figure 2D). The developed Tyr kinase biosensor exhibited a good repeatability of 3.72% RSD (Relative standard deviation) for a repetitive measurement of 3 μM p60c-src substrate I (n = 12). In addition, the developed Tyr kinase biosensor exhibited a linear dynamic range of 0.1−10 μM (R2 = 0.998) for p60c-src
substrate I (Figure 2E) with the lowest detection limit (LOD) of 50 nM (S/N≥ 3). These results imply that the developed Tyr kinase biosensor can be used to make repetitive measurements of tyrosyl residue-bearing peptides.
Determining Src Activity. The feasibility of using the Tyr kinase biosensor to measure Src activity was further studied using p60c-src substrate I and ATP as substrates. Src phosphorylation was expected to reduce the electrochemical responses of the biosensor to the p60c-src substrate I. As shown in Figure 3A, the available tyrosyl residues in the reaction mixture without Src remained almost unchanged by 2 h of incubation. However, in the presence of Src, the electro-chemical response of Tyr kinase biosensor declined in a time-dependent manner for 30 min, and then more slowly for a further 90 min (Figure 3A). This result suggests Src indeed phosphorylates p60c-src substrate I, and that this process can be detected electrochemically using the Tyr kinase biosensor. Although p60c-src substrate I contains two tyrosyl residues, the second tyrosyl residue from the N-terminus is the major phosphorylation site for Src.50 Therefore, the electrochemical response of the developed biosensor to p60c-src substrate I reached a plateau even after 120 min of reaction (Figure 3A). The kinase activity of various concentrations of Src (0.26, 0.53, 1.06, 2.11, 4.22, 8.45, 16.86, and 33.78 nM) at 30°C for 30 min
Figure 2.Electrochemical responses of Tyr kinase biosensor to various peptides. (A) Steady state amperometric response of Tyr kinase biosensor to dipeptides, 100 μM Tyr-Ala (top penal) and Gly-Tyr (bottom panel). (B) Amperometric response of Tyr kinase biosensor to 3 μM p60c-src substrate I. (C) Step responses of Tyr kinase biosensor with successive additions of 1μM p60c-src substrate I in PBS buffer. (D) Repeatability of the developed Tyr kinase biosensor. Repetitive measurements of the responses of Tyr kinase biosensor to 3μM p60c-src substrate I were carried out 12 times in PBS (pH 6.5) with constant stirring. The working potential was set at−0.2 V vs Ag/AgCl. (E) Calibration curves of p60c-src substrate I on Tyr kinase biosensor. Amperometic responses to p60c-src substrate I at various concentrations were determined in PBS buffer at working potential of−0.2 V vs Ag/AgCl. Measurements were made in PBS buffer with constant stirring. Inset presents linear range of detection of p60c-src substrate I. Data are presented as the mean± SD from three independent measurements.
was also measured (Figure 3B). As shown in Figure 3B (inset), the kinase activity increased linearly with the logarithm of the Src concentration from 0.26 to 33.78 nM, or from 1.9 to 237.6 U/mL, with the lowest detection limit for Src of 0.23 U/mL (S/N ≥ 3). For comparison, the previously developed electrochemical sensing systems for Src had a linear range of detection and the lowest detection limit of 10−200 U/mL and 5 U/mL,31,32 respectively.
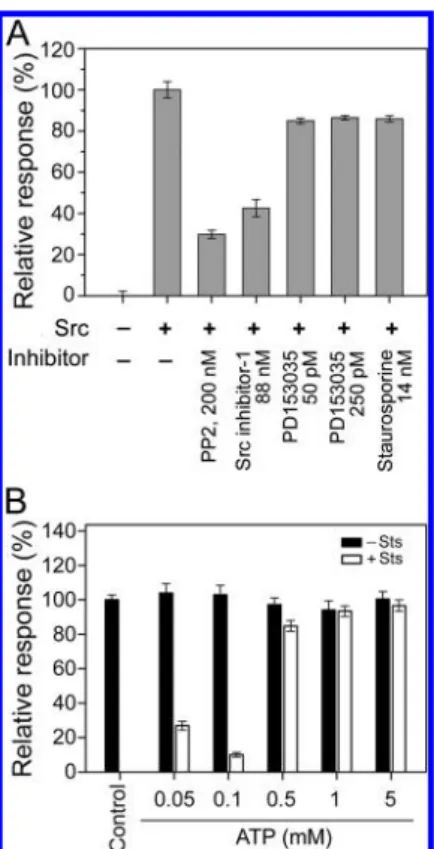
Effect of Protein Kinase Inhibitors on Src Activity. The capacity of Tyr kinase biosensor to detect the activity of Src was also studied by measuring the effectiveness of various protein kinase inhibitors PP2, Src inhibitor-1, PD153035 and staurosporine. PP2 is a specific inhibitor of Src with an IC50
of around 100 nM.51,32Src inhibitor-1 is a selective, competitive inhibitor of Src and represents a gold standard for Src inhibition with an IC50 of 44 nM.
52
As expected, 200 nM PP2 and 88 nM Src inhibitor-1 effectively suppressed the activity of Src by around 65% and 58%, respectively (Figure 4A). PD153035, a specific kinase inhibitor of the epidermal growth factor receptor (EGFR) with an IC50 of 25 pM,53,54
slightly inhibited the activity of Src (by around 14%) at concentrations of 50 and 250 pM.
Staurosporine is a prototypic ATP-competitive inhibitor of both protein serine/threonine kinases and tyrosine kinases, with an IC50 in the nanomolar range.55,56ATP competes with staurosporine for the nucleotide binding site in the protein kinases.57 Therefore, the failure of 14 nM staurosporine to suppress Src activity in the presence of 5 mM ATP (Figure 4A) is not surprising. The effect of ATP was further verified by the
fact that staurosporine suppressed the activity of Src by 70 to 90% in the presence of low concentrations of ATP (≤100 μM), but this effect was largely eliminated at high concentrations (ATP ≥ 500 μM) (Figure 4B). The dose-dependence of inhibition by Src inhibitor-1 of Src activity was further studied. Src inhibitor-1 effectively suppressed Src activity in a dose-dependent manner at concentrations from 5 to 400 nM, with an IC50of around 45 nM (Figure 5A), which is very close to the
value determined elsewhere.52 These results suggest that the effect of various inhibitors on protein tyrosine kinases can be easily determined using the developed amperometric tyrosine kinase biosensor.
The capability of the developed Tyr kinase biosensor to detect Src activity in cell lysate prepared from cells treated with and without epidermal growth factor (EGF), which has been shown to induce Src activity. As shown in Figure 5B, the activity of Src was almost undetectable in HeLa cells without EGF treatment. However, after EGF treatment the activity of Src was induced. This result clearly showed that the developed
Figure 3.Electrochemical measurements of Src activity. (A) Time-dependent reaction curve of Src. Activity of Src (4.22 nM) was measured in reaction buffer that contained 3 mM p60c-src substrate I and 5 mM ATP at 30 °C for 0, 30, 60, 90, and 120 min, before electrochemical measurements were made. (B) Dose−response curve of Src obtained by incubating various concentrations of Src (0.26, 0.53, 1.06, 2.11, 4.22, 8.45, 16.86, and 33.78 nM) in reaction buffer that contained 3 mM p60c-src substrate I and 5 mM ATP at 30°C for 30 min. (Inset) Semilog plot of response as a function of Src concentration. Data are presented as mean ± SD from three independent measurements.
Figure 4.Inhibition of Src activity by protein kinase inhibitors. (A) Activity of Src was determined by incubating Src (4.22 nM) with 3 mM p60c-src substrate I and 5 mM ATP in the absence or presence of 200 nM PP2, 88 nM Src inhibitor I, 25 and 250 pM PD153035, and 14 nM staurosporine. The reaction was performed at 30°C for 30 min before electrochemical determination. (B) Effect of various concen-trations of ATP on inhibitory activity of staurosporine (Sts). Src (4.22 nM) was incubated with 3 mM p60c-src substrate I and 14 nM staurosporine in the presence of various concentrations of ATP (0.05, 0.1, 0.5, 1, and 5 mM) at 30°C for 30 min, before electrochemical determination. Dimethyl sulfoxide (DMSO) was added into the kinase reaction mixtures without staurosporine. The Src reaction mixture containing 3 mM p60c-src substrate I and 0.5 mM ATP without DMSO and staurosprine was used as an external control (Control). The relative response (%) of all other reactions was calculated by using the Control as 100%. Data are presented as mean± SD from three independent measurements.
Tyr kinase biosensor is capable of detecting the Src activity in cell lysate.
■
CONCLUSIONA simple, sensitive, reusable, and label-free electrochemical system for sensing the activity of protein tyrosine kinases was developed. The measurement of protein tyrosine kinase activity is based on the ability of tyrosinase to differentially recognize phosphorylated and unphosphorylated tyrosyl residues in peptides before and after the reaction of tyrosine kinases (Scheme 1). Phosphorylation at the hydroxyl group may block the access of the tyrosyl residues to the active site of tyrosinase. The activity of protein tyrosine kinase can be easily determined using the developed tyrosine kinase biosensor by measuring the amount of remaining tyrosyl residues in the peptide substrates. Unlike previously developed electrochemical sensing systems, the experimental system developed herein requires no peptide substrate immobilization on the electrode surface. Most importantly, the activity of tyrosine kinases can be measured repetitively on the Tyr kinase biosensor without alkaline phosphatase treatment. In conclusion, the developed tyrosine kinase sensor was demonstrated to be highly sensitive to tyrosine kinase activity, with a linear response over a wide range. It has great potential for use in drug screen and diagnostic applications.
■
AUTHOR INFORMATIONCorresponding Author
*Address: Department of Biological Science and Technology, National Chiao Tung University, 75 Po-Ai Street, Hsinchu, Taiwan 300, ROC. Tel: 5731735. Fax: +886-3-5729288. E-mail: [email protected].
■
ACKNOWLEDGMENTSC.-L.W. and L.-Y.W. contributed equally to this article. The authors would like to thank the Ministry of Education, Executive Yuan, Republic of China (Taiwan) and National Science Council of the Republic of China (Taiwan), for financially supporting this research under Contract No. NSC-97-2311-B-009-001 MY3.
■
REFERENCES(1) Manning, G.; Whyte, D. B.; Martinez, R.; Hunter, T.; Sudarsanam, S. Science 2002, 298, 1912−1934.
(2) Hunter, T.; Plowman, G. D. Trends Biochem. Sci. 1997, 1, 18−22. (3) Hunter, T. Cell 1995, 80, 225−236.
(4) Graves, J. D.; Krebs, E. G. Pharmacol. Ther. 1999, 82, 111−121. (5) Montero1, J. C.; Seoane1, S.; Alberto Ocaña, A.; Pandiella, A. Clin. Cancer Res. 2011, 17, 5546−5552.
(6) Fischer, A.; Picard, C.; Chemin, K.; Dogniaux, S.; le Deist, F.; Hivorz, C. Semin. Immunopathol. 2010, 32, 107−116.
(7) Bertrand1, L.; Horman, S.; Beauloye, C.; Vanoverschelde, J.-L. Cardiovasc. Res. 2008, 79, 238−248.
(8) Matrougui, K. Diabetes Metab. Res. Rev. 2010, 26, 13−16. (9) Edwards, J. Expert Opin. Investig. Drugs. 2010, 19, 605−614. (10) Koresawa, M.; Okabe, T. Drug Dev. Technol. 2004, 2, 153−160. (11) Krystof, V.; Uldrijan, S. Curr. Drug Targets 2010, 11, 291−302. (12) Singh, J.; Petter, R. C.; Kluge, A. F. Curr. Opin. Chem. Biol. 2010, 14, 475−80.
(13) Pearson, R. B.; Kemp, B. E. Methods Enzymol. 1991, 200, 62− 81.
(14) Geer, P.; Hunter, T. Electrophoresis 1994, 15, 544−554. (15) Houseman, B. T.; Huh, J. H.; Kron, S. J.; Mrksich, M. Nat. Biotechnol. 2002, 20, 270−274.
(16) Diks, S. H.; Kok, K.; O’Toole, T.; Hommes, D. W.; van Dijken, P.; Joore, J.; Peppelenbosch, M. P. J. Biol. Chem. 2004, 279, 49206− 49213.
(17) Green, K. D.; Pflum, M. K. H. J. Am. Chem. Soc. 2007, 129, 10− 11.
(18) Wang, Z.; Lee, J.; Cossins, A. R.; Brust, M. Anal. Chem. 2005, 77, 5770−5774.
(19) Wang, Z.; Levy, R.; Fernig, D. G.; Brust, M. J. Am. Chem. Soc. 2006, 128, 2214−2215.
(20) Xu, X. H.; Liu, X.; Nie, Z.; Pan, Y. L.; Guo, M. L.; Yao, S. Z. Anal. Chem. 2011, 83, 52−59.
(21) Freeman, R.; Finder, T.; Gill, R.; Willner, I. Nano Lett. 2010, 10, 2192−2196.
(22) Allen, M. D.; DiPilato, L. M.; Rahdar, M.; Ren, Y. R.; Chong, C.; Liu, J. O.; Zhang, J. ACS Chem. Biol. 2006, 1, 371−376.
(23) Kane, S.; Sano, H.; Liu, S. C.; Asara, J. M.; Lane, W. S.; Garner, C. C.; Lienhard, G. E. J. Biol. Chem. 2002, 277, 22115−22118.
(24) Pandey, A.; Podtelejnikov, A. V.; Blagoev, B.; Bustelo, X. R.; Mann, M.; Lodish, H. F. Proc. Natl. Acad. Sci. U.S.A. 2000, 97, 179− 184.
(25) Chen, C. A.; Yeh, R. H.; Lawrence, D. S. J. Am. Chem. Soc. 2002, 124, 3840−3841.
(26) Inamori, K.; Kyo, M.; Nishiya, Y.; Inoue, Y.; Sonoda, T.; Kinoshita, E.; Koike, T.; Katayama, Y. Anal. Chem. 2005, 77, 3979− 3985.
(27) Ptacek, J.; Snyder, M. Trends Genet. 2006, 22, 545−554. (28) Beausoleil, S. A.; Villén, J.; Gerber, S. A.; Rush, J.; Gygi, S. P. Nat. Biotechnol. 2006, 24, 1285−1292.
Figure 5.Characterization of Src activity. (A) Activity of Src (4.22 nM) was determined in reaction buffer that contained 3 mM p60c-src substrate I, 0.1 mM ATP and various concentrations of Src inhibitor I (5, 10, 25, 45, 90, 200, and 400 nM) at 30°C for 30 min before electrochemical determination. (B) Electrochemical measurement of Src activity in cells treated with EGF. HeLa cell lysate (20μg) was incubated with the kinase reaction buffer containing 2.5 mM p60c-src substrate I and 0.1 mM ATP at 25 °C for 60 min prior to the electrochemical determination of peptide phosphorylation. (* p < 0.05) Data are presented as mean ± SD from three independent measurements.
(29) Freeman, R.; Gill, R.; Willner, I. Chem. Commun. 2007, 3450− 3452.
(30) Ji, J.; Yang, H.; Liu, Y.; Chen, H.; Kong, J.; Liu, B. Chem. Commun. 2009, 1508−1510.
(31) Kerman, K.; Vestergaard, M.; Tamiya, E. Anal. Chem. 2007, 79, 6881−6885.
(32) Kerman, K.; Chikae, M.; Yamamura, S.; Tamiya, E. Anal. Chim. Acta 2007, 588, 26−33.
(33) Kerman, K.; Kraatz, H.-B. Biosens. Bioelectron. 2009, 24, 1484− 1489.
(34) Song, H. F.; Kerman, K.; Kraatz, H. B. Chem. Commun. 2008, 502−504.
(35) Wang, J.; Shen, M.; Cao, Y.; Li, G. X. Biosens. Bioelectron. 2010, 26, 638−642.
(36) Xu, X. H.; Nie, Z.; Chen, J. H.; Fu, Y. C.; Li, W.; Shen, Q. P.; Yao, S. Z. Chem. Commun. 2009, 6946−6948.
(37) Wang, J.; Cao, Y.; Li, Y.; Liang, Z.; Li, G. J. Electroanal. Chem. 2011, 656, 274−278.
(38) Kerman, K.; Song, H. F.; Duncan, J. S.; Litchfield, D. W.; Kraatz, H.-B. Anal. Chem. 2008, 80, 9395−9401.
(39) Martić, S.; Rains, M. K.; Freeman, D.; Kraatz, H.-B. Bioconjugate Chem. 2011, 22, 1663−1672.
(40) Martić, S.; Labib, M.; Kraatz, H.-B. Analyst 2011, 136, 107−112. (41) Wieckowska, A.; Li, D.; Gill, R.; Willner, I. Chem. Commun. 2008, 2376−2378.
(42) Zhou, H.; Liu, L.; Yin, K.; Liu, S.; Li, G. Electrochem. Commun. 2006, 8, 1168−1172.
(43) Kitajima, N.; Koda, T.; Iwata, Y.; Moro-oka, Y. J. Am. Chem. Soc. 1990, 112, 8833−8839.
(44) Martin, G. S. Nat. Rev. Mol. Cell Biol. 2001, 2, 467−475. (45) Summy, J. M.; Gallick, G. E. Cancer Metastasis Rev. 2003, 22, 337−358.
(46) Ishizawar, R.; Parsons, S. J. Cancer Cell 2004, 6, 209−214. (47) Finn, R. S. Ann. Oncol 2008, 19, 1379−1386.
(48) Yuan, C.-J.; Wang, C.-L.; Chao, W.-C.; Huang, K.-C. Biosens. Bioelectron. 2011, 26, 2858−2863.
(49) Lam, K. S.; Lam, K. S.; Wu, J.; Lou, Q. Int. J. Pept. Protein Res. 1995, 45, 587−592.
(50) Uttamchandani, M.; Chan, E. W. S.; Chen, G. Y. J.; Yao, S. Q. Bioorg. Med. Chem. Lett. 2003, 13, 2997−3000.
(51) Hanke, J. H.; Gardner, J. P.; Dow, R. L.; Changelian, P. S.; Brissette, W. H.; Weringer, E. J.; Pollok, B. A.; Connelly, P. A. J. Biol. Chem. 1996, 271, 695−701.
(52) Tian, G.; Cory, M.; Smith, A. A.; Knight, W. B. Biochem. 2001, 40, 7084−7091.
(53) Yanagihara, N.; Tachikawa, E.; Izumi, F.; Yasugawa, S.; Yamamoto, H.; Miyamoto, E. J. Neurochem. 1991, 56, 294−298.
(54) Bridges, A. J.; Zhou, H.; Cody, D. R.; Rewcastle, G. W.; McMichael, A.; Showalter, H. D.; Fry, D. W.; Kraker, A. J.; Denny, W. A. J. Med. Chem. 1996, 39, 267−276.
(55) Takahashi, I.; Saitoh, Y.; Yoshida, M.; Sano, H.; Nakano, H.; Morimoto, M.; Tamaoki, T. J. Antibiot. (Tokyo) 1989, 42, 571−576.
(56) Miyano, N.; Kinoshita, T.; Nakai, R.; Kirii, Y.; Yokota, K.; Tada, T. Bioorg. Med. Chem. Lett. 2009, 19, 6557−6560.
(57) Herbert, J. M.; Seban, E.; Maffrand, J. P. Biochem. Biophys. Res. Commun. 1990, 171, 189−195.