Elsevier Scientific Publishers Ireland Ltd.
R E G U L A T I O N O F P R O L I N E A C C U M U L A T I O N I N D E T A C H E D R I C E L E A V E S
IRENE T. CHOU, CHIEN TEH CHEN and CHING HUEI KAO*
Department of Agronomy, National Taiwan University, Taipei, Taiwan (Republic of ChinaJ (Received December 20th, 1989)
(Revision received March 23rd, 1990} (Accepted March 29th, 1990)
Regulation of proline accumulation in detached rice leaves was investigated. Under dark conditions, proline content in detached rice leaves remained unchanged 8 h after incubation in distilled water, but increased 2-fold by 24 h. However, proline content did not increase during the entire 24-h incubation period in the light. Cell sap pH did not change at 8 h, but decreased 0.2 units at 24 h after incubation in darkness. No significant decrease in cell sap pH was observed in detached rice leaves incu- bated in the light. The increase of proline content in ammonium vanadate-treated detached rice leaves was associated with cell
sap
acidification. However, ammonium chloride also decreased cell sap pH and increased the proline content to some extent. The addition of fusicoccin resulted in an increase in proline content and a decrease in cell sap pH. Although fusicoccin prom- oted isobutyric acid-induced increase of proline, fusicoccin did not result in further decrease in cell sap pH induced by isobu- tyric acid. Cycloheximide or cordycepin increased proline content without changing cell sap pH. It is concluded that factors apart from cell sap acidification may also be involved in the regulation of proline accumulation.Key words: cell sap pH; fusicoccin; isobutyric acid; Oryza sativa; proline accumulation; vanadate
I n t r o d u c t i o n
I t is well e s t a b l i s h e d t h a t proline accumu- lates r a p i d l y in excised and i n t a c t leaves u n d e r w a t e r s t r e s s [ 1 - 3 ] . P r o l i n e a c c u m u l a t i o n can also be induced b y abscisic acid in b o t h excised leaves and i n t a c t p l a n t s [4,5]. R e c e n t r e p o r t s s u g g e s t e d t h a t d e c r e a s e of i n t r a c e l l u l a r pH, m e a s u r e d as cell sap pH, w a s i n v o l v e d in stress-, abscisic acid- and w e a k acid-induced proline a c c u m u l a t i o n in w h e a t coleoptiles a n d b a r l e y leaves [ 6 - 9 ] . V e n e k a m p [10] s h o w e d t h a t o r g a n i c acids w e r e t h e s o u r c e for d r o u g h t - induced proline s y n t h e s i s in field-grown b e a n plants. P r e v i o u s l y , we h a v e r e p o r t e d t h a t pro- line a c c u m u l a t e d in d e t a c h e d rice leaves d u r i n g d a r k - i n d u c e d s e n e s c e n c e [11]. W e also s h o w e d t h a t t r e a t m e n t s which r e t a r d s e n e s c e n c e inhibit proline accumulation, while a g e n t s which p r o m o t e s e n e s c e n c e p r o m o t e proline *To whom all correspondence should be addressed.
a c c u m u l a t i o n [12]. R e c e n t l y , we d e m o n s t r a t e d t h a t t h e d e c r e a s e in r a t e of p r o t o n s e c r e t i o n of d e t a c h e d rice leaves p l a y e d a r e g u l a t o r y role in s e n e s c e n c e of d e t a c h e d rice leaves [13]. T h e c o n s e q u e n c e of t h e d e c r e a s e in r a t e of p r o t o n s e c r e t i o n would be a d e c r e a s e in i n t r a c e l l u l a r pH. T h e p r e s e n t i n v e s t i g a t i o n w a s t h u s con- d u c t e d to s t u d y t h e possible i n v o l v e m e n t of l o w e r e d i n t r a c e l l u l a r p H in r e g u l a t i n g proline a c c u m u l a t i o n in d e t a c h e d rice leaves.
M a t e r i a l s a n d M e t h o d s
Rice (Oryza sativa L. cv. T a i c h u n g N a t i v e 1) seedlings w e r e g r o w n in a g r e e n h o u s e with n a t u r a l light at 30 °C day/25 °C night. T h e apical 3-cm s e g m e n t s excised f r o m t h e t h i r d leaves of 12-day-old seedlings w e r e used. A g r o u p of 20 s e g m e n t s w e r e floated in a P e t r i dish contain- ing 20 ml of distilled w a t e r or t e s t solutions. Unless o t h e r w i s e indicated, all t e s t solutions and distilled w a t e r w e r e a d j u s t e d to pH 5.5. 0168-9452/90/$03.50 © 1990 Elsevier Scientific Publishers Ireland Ltd.
I n c u b a t i o n w a s c a r r i e d out at 27 °C in d a r k n e s s
or in t h e light (16.7 W m -2) p r o v i d e d by a mix-
t u r e of cool-white a n d G r o l u x l a m p s .
P r o l i n e w a s e x t r a c t e d a n d its c o n c e n t r a t i o n
d e t e r m i n e d following t h e m e t h o d of B a t e s et al.
[15]. L e a f s e g m e n t s w e r e h o m o g e n i z e d in 30/o
(w/v) sulfosalicylic acid a n d c e n t r i f u g e d . T h e
s u p e r n a t a n t fluid w a s t r e a t e d w i t h acetic acid
a n d acid-ninhydrin, boiled for 1 h and t h e
a b s o r b a n c e w a s d e t e r m i n e d at 520 nm. P r o l i n e
c o n t e n t w a s e x p r e s s e d as nmol/20 s e g m e n t s .
F o r t h e m e a s u r e m e n t of t h e cell s a p p H , t h e
m e t h o d of P e s c i a n d B e f f a g n a [6] w a s used. L e a f
s e g m e n t s w e r e t r a n s f e r r e d at t h e end of t h e
t r e a t m e n t s d i r e c t l y into s y r i n g e s and w e r e
frozen in liquid n i t r o g e n . A f t e r t h a w i n g t h e cell
s a p w a s p r e s s e d o u t f r o m t h e s y r i n g e and t h e
p H w a s m e a s u r e d .
All e x p e r i m e n t s w e r e r e p e a t e d one or m o r e
t i m e s w i t h s i m i l a r r e s u l t s . T h e d a t a r e p o r t e d
h e r e a r e f r o m a single e x p e r i m e n t .
R e s u l t sT a b l e I s h o w s t h e c h a n g e s of p r o l i n e c o n t e n t
and t h e cell s a p p H of d e t a c h e d rice l e a v e s incu-
b a t e d in distilled w a t e r u n d e r d a r k a n d light
conditions. In d a r k n e s s , p r o l i n e c o n t e n t in
d e t a c h e d rice l e a v e s did n o t i n c r e a s e a t 8 h
a f t e r i n c u b a t i o n b u t i n c r e a s e d 2-fold a t 24 h,
which c o n f i r m e d o u r p r e v i o u s r e s u l t s [11]. Pro-
line c o n t e n t , h o w e v e r , in d e t a c h e d rice l e a v e s
i n c u b a t e d in t h e light d e c r e a s e d s i g n i f i c a n t l y
b y 8 h and s u b s e q u e n t l y r e m a i n e d low t h r o u g h -
out t h e i n c u b a t i o n period. Cell s a p p H in
d e t a c h e d rice l e a v e s r e m a i n e d u n c h a n g e d a t 8
Table I.
Changes of proline content and cell sap pH in
detached rice leaves incubated in distilled water under dark
and light conditions. Mean + S.E., three repetitions.
Time Proline content
Cell sap pH
(h)
Light
Dark
Light
Dark
0
47 _+ 2
6.24 _+ 0.03
8
22 _+ 1
50 _+ 3
6.24 _+ 0.03
24
23 _+ 2
92 _+
5 6.18 _+ 0.016.24 _+ 0.01
6.05 _+ 0.04
h, b u t had d e c r e a s e d 0.2 units b y 24 h of incuba-
tion in d a r k n e s s . No significant d e c r e a s e in cell
s a p p H w a s o b s e r v e d w h e n d e t a c h e d rice
l e a v e s w e r e i n c u b a t e d in t h e light. Clearly, pro-
line a c c u m u l a t i o n is a s s o c i a t e d w i t h a d e c r e a s e
in cell s a p p H of d e t a c h e d rice l e a v e s .
Since n e i t h e r i n c r e a s e in p r o l i n e c o n t e n t nor
cell s a p acidification w a s o b s e r v e d at 8 h a f t e r
incubation of d e t a c h e d rice l e a v e s in d a r k n e s s
or t h e light and in o r d e r to avoid i n t e r f e r e n c e
by t h e f a c t o r s r e l a t e d to s e n e s c e n c e in long-
t e r m incubations, an 8-h i n c u b a t i o n t i m e w a s
u s e d for all e x p e r i m e n t s d i s c u s s e d below.
S e v e r a l lines of e v i d e n c e i n d i c a t e t h a t a plas-
m a l e m m a A T P a s e a c t s as an e l e c t r o g e n i c pro-
ton p u m p in h i g h e r p l a n t s [16]. R e c e n t w o r k b y
B e f f a g n a a n d R o m a n i [17] s h o w e d t h a t vana-
d a t e inhibited t h e a c t i v i t y of t h e p r o t o n p u m p ,
c a u s i n g a d e c r e a s e in t h e i n t r a c e l l u l a r p H ,
m e a s u r e d as cell s a p p H . T h e d a t a in T a b l e I I
s h o w t h a t t h e addition of v a n a d a t e leads to a
significant d e c r e a s e in cell s a p p H . I f cell s a p
acidification is of i m p o r t a n c e in r e g u l a t i n g
proline a c c u m u l a t i o n , t h e n t h e i n c r e a s e in pro-
line c o n t e n t is to be e x p e c t e d in v a n a d a t e -
t r e a t e d d e t a c h e d rice l e a v e s . A s i n d i c a t e d in
Fig. 1, this is indeed t h e case. R e s u l t s in Fig. 1
also indicate t h a t a m m o n i u m chloride i n c r e a s e d
t h e proline c o n t e n t in d e t a c h e d rice l e a v e s
a l t h o u g h less m a r k e d l y . T h i s i n c r e a s e is a p p a r -
e n t l y a s s o c i a t e d w i t h a d e c r e a s e in cell s a p p H
(Table II).
Table
II.
Effect of ammonium vanadate on the cell sap
pH in detached rice leaves. Cell sap pH was measured at 8 h
after incubation. Initial cell sap pH was 6.39 __. 0.04. Mean
_+ S.E., three repetitions.
Treatment
Cell sap pH
Light
Dark
Water
6.35 _+ 0.02
6.34 _+ 0.02
Ammonium chloride
6.04 _+ 0.03
6.09 _+ 0.01
(5raM)
Ammonium vanadate
5.94 _+ 0.01
5.90 _+ 0.02
(5mM)
LIGHT
8 0 t/) I - 4 0|
" ' O
a . ~ 120
"6
E
©
8O 4 0O
DARK 120 iH20
N H 4 C I NH4VO
3
Fig. 1. Effect of ammonium vanadate on proline content in detached rice leaves. Detached rice leaves were incu- bated in deionized water, ammonium chloride (5 raM) or ammonium vanadate (5 mM) for 8 h. Initial proline content was 29 ± 1 nmol/20 segments. Bars represent S.E., three repetitions.
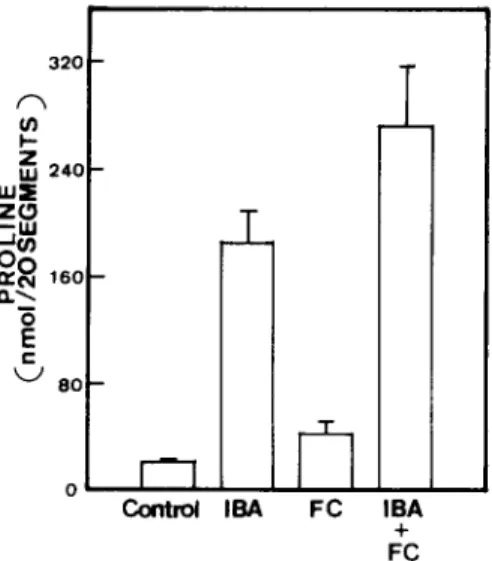
T h e r e is w i d e c o n s e n s u s t h a t p h y t o t o x i n f u s i c o c c i n (FC) s t i m u l a t e s a c t i v e p r o t o n e x p o r t [18]. S u r p r i s i n g l y , t r e a t m e n t w i t h F C c a u s e d n o t o n l y a n i n c r e a s e i n p r o l i n e c o n t e n t b u t a l s o a d e c r e a s e i n cell s a p p H (Fig. 2, T a b l e III). T r e a t m e n t w i t h i s o b u t y r i c a c i d (IBA) for 8 h r e s u l t e d i n a n i n c r e a s e i n p r o l i n e c o n t e n t {Fig. 3). T h e t r e a t m e n t w i t h i s o b u t y r i c acid is a c c o m - p a n i e d b y a d e c r e a s e i n t h e i n t r a c e l l u l a r p H ( T a b l e III). A l t h o u g h F C e f f e c t i v e l y i n c r e a s e d I B A - i n d u c e d p r o l i n e a c c u m u l a t i o n , F C d i d n o t r e s u l t i n a n y a d d i t i o n a l d e c r e a s e i n cell s a p p H g r e a t e r t h a n t h a t c a u s e d b y I B A (Fig. 3, T a b l e III).
Table III. Effect of FC and isobutyric acid (IBA) on the cell sap pH in detached rice leaves. Detached rice leaves were incubated in the light for 8 h in 10 mM potassium phos- phate buffer in the absence or presence of FC (10 -5 M), IBA (10 raM) or FC (10 -s M) + IBA (10 mM). Initial cell sap pH was 6.14 ± 0.02 and 6.17 _+ 0.01, respectively, for Experi- ments I and II. Mean _+ S.E., three repetitions.
Treatment Cell sap pH
Experiment I Control 6.16 _ 0.01 FC 6.05 ± 0.02 Experiment H Control 6.19 _ 0.01 IBA 6.02 _+ 0.01 FC 6.09 _+ 0.01 IBA + FC 6.05 ± 0.02 50 4 0
_z-,
3o E~.~ 20
10 nO-C) UGH"
o - e DARK i ,/ I I I 0 10 -7 10- 6 10- s F C ( M )Fig. 2. Effect of different concentrations of FC on proline content in detached rice leaves. Detached rice leaves were incubated for 8 h in 10 mM potassium phosphate buffer in the presence of FC at different concentrations. Initial pro- line content was 46 +-- 6 nmol/20 segments. Bars represent S.E., three repetitions.
LIGHT 320 I- UJ~ 2 4 0 z O C ~ 80 O Control IBA m FC IBA + FC
Fig. 3. Effect of fusicoccin (FC) and isobutyric acid (IBA)
on proline content in detached rice leaves. Detached rice leaves were incubated in the light for 8 h in 10 mM potas- sium phosphate buffer in the absence or presence of FC (10 -5 M), IBA (10 mM) or FC (10 -~ M) + IBA (10 mM). Initial proline content was 30 _+ 3 nmol/20 segments. Bars repre- sent S.E., t h r e e repetitions.
Our previous work showed that proton
secretion by detached rice leaves is inhibited
by cycloheximide (CHI) and suggested that pro-
tein synthesis is required for proton secre-
tion [19]. It was expected that t r e a t m e n t of
detached rice leaves with CHI would cause a
decrease in cell sap pH. Contrary to our
expectation, neither CHI nor cordycepin (COR),
an inhibitor of RNA synthesis, lowered the cell
sap pH (Table IV). Figure 4 shows the effect of
CHI and COR on proline content in detached
rice leaves. The addition of CHI or COR
resulted in a significant increase in proline con-
Table IV. Effect of cycloheximide (CHI) and cordycepin
(COR) on the cell sap pH in detached rice leaves. Detached rice leaves were incubated in the light for 8 h. Initial cell pH was 6.07 ± 0.02. Mean _+ S.E., t h r e e repetitions.
Treatment Cell sap pH
Water 6.08 ___ 0.01 COR (0.1 raM) 6.08 ± 0.02 CHI (0.1 raM) 6.12 ± 0.03 80 40 (J 8O 4 o Z I¢1 o o a. ~_ 1 2 0 0 E H20 CHI COR DARK
,2o
Fig. 4. Effect of cycloheximide (CHI) and cordycepin
(COR) on proline content in detached rice leaves. Detached rice leaves were incubated in CHI (0.1 raM) or COR (0.1 mM) for 8 h. Initial proline content was 21 + 1 nmol/20 seg- ments. Bars r e p r e s e n t S.E., t h r e e repetitions.
tent. Obviously, CHI- or COR-induced increase
of proline content is unrealted to the shift of
cell sap pH.
Discussion
The present investigation was initiated to
elucidate the possible involvement of intracel-
lular pH, measured as cell sap pH, in regulating
proline accumulation in detached rice leaves.
Several lines of evidence suggested that cell
sap acidification was involved in regulating
proline accumulation [6--10]. The results pre-
sented in this paper are generally consistent
with this suggestion.
increased the proline content was unexpected.
This further suggests that proline accumula-
tion depends on cell sap pH, since acidification
of the cell sap has been established on treating
detached rice leaves with ammonium chloride.
Of particular interest is the finding that FC
increased proline content and decreased cell
sap pH. However, Pesci and Beffagna [6]
reported that FC decreased proline content and
increased cell sap pH in detached barley leaves.
The use of a different species may have led to
this discrepancy. The decrease of intracellular
pH following FC application has also been
reported by other investigators
[20--23].
Although FC promoted IBA-induced proline
accumulation in detached rice leaves, FC did
not cause a further decrease in cell sap pH
induced by IBA. It appears that factors apart
from cell sap acidification may also be involved
in regulating proline accumulation. This conclu-
sion was further supported by the observation
that CHI or COR increased proline content
without lowering cell sap pH.
Using detached tobacco leaves. Bogges and
S t e w a r t [24] demonstrated that CHI t r e a t m e n t
resulted in a striking increase in proline con-
tent. They attributed the elevated proline con-
tent observed to w a t e r stress r a t h e r than to
reduced protein synthesis. Since, in the present
work, no curling and loss of turgor were evi-
dent in CHI- or COR-treated detached rice
leaves, it seemed unlikely that proline accumu-
lation induced by CHI or COR was due to water
stress. It seems most likely that CHI- or COR-
induced proline accumulation is a consequence
of the inhibition of protein synthesis and/or
synthesis of specific enzymes responsible for
proline oxidation. At the present time, it
remains unclear w h e t h e r cell sap acidification
regulates
proline
accumulation
through
stimulation of proline synthesis, reduction of
proline utilization, or both. F u r t h e r work along
this line would help in elucidating the mecha-
nism of proline accumulation.
Acknowledgement
This research was supported by the National
Science Council, Republic of China.
References
1 T.C. Hsiao, P l a n t response to w a t e r stress. Annu. Rev. Plant Physiol., 24 (1973) 519--570.
2 T.N. Singh, L.G. Aspinall and L.G. Paleg, Nitrogen metabolism and growth in the barley plant during w a t e r stress. Aust. J. Biol. Sci., 26 (1973145- 56. 3 R.P. Waldren and I.P. Teare, F r e e proline accumula-
tion in drought stressed plants u n d e r laboratory condi- tions. Plant Soil, 40 (1974)689--692.
4 D. Aspinall, T.N. Singh and L.G. Paleg, Stress metabo- lism. V. Abscisic acid and nitrogen metabolism in bar- ley and Lolium temulentum L. Aust. J. Biol. Sci., 26 (1973) 319-- 329.
5 C.R. Stewart, The mechanism of abscisic acid-induced proline accumulation in barley leaves. Plant Physiol., 66 (1980) 2 3 0 - 233.
6 P. Pesci and N. Beffagna, Inhibiting effect of fusicoccin on abscisic acid-induced proline accumulation in barley leaves. Plant Sci. Lett., 36 (1984) 7 - 12.
7 P. Pesci and N. Beffagna, Effects of weak acids on pro- line accumulation in barley leaves: a comparison between abscisic acid and isobutyric acid. Plant, Cell Environ.; 8 (1985) 1 2 9 - 1 3 3 .
8 H. Goring and F. Plescher, Proline accumulation induced by weak acids and IAA in coleoptiles of wheat seedlings. Biol. Plant., 28 (1986) 401 - 406.
9 K. Zoglauer, H. Dembny and H. Goring, Inhibition of IAA-induced ethylene production and proline accumu- lation in what coleoptiles by fusicoccin. Biochem. Phy- siol. Pflanzen, 182 (1987) 2 3 - 2 9 .
10 J.H. Venekamp, Regulation of cytosol acidity in plants under conditions of drought. Physiol. Plant., 76 (1989) 1 1 2 - 1 1 7 .
11 C.Y. Wang, S~I. Cheng and C.H. Kao, Senescence of rice leaves VII. Proline accumulation in senescing excised leaves. Plant Physiol., 69 (1982) 1348-13449. 12 C.Y. Wang and C.H. Kao, Senescence of rice leaves
VIII. The regulation of proline accumulation during senescence. Bot. Bull. Acad. Sin., 24 (1983) 1 4 5 - 1 5 4 . 13 C.T. Chen, I.T. Chou and C.H. Kao, Senescence of rice
leaves XX. Changes of proton secretion during senesc- ence. Plant Sci., 66 (1990) 29-- 34.
14 W.P. Hurng, T.L. Lin, S.S. Ren, J.C. Chen, R.R. Chen and C.H. Kao, Senescence of rice leaves XVIII. Changes of stomatal a p e r t u r e during senescence. Plant Cell Physiol., 29 (1988) 27 - 3 1 .
15 L.S. Bates, Rapid determination of free proline for water s t r e s s studies. Plant Soil, 29 (1973) 205-- 207. 16 R.M. Spanswick, Electrogenic ion pumps. Annu. Rev.
Plant Physiol., 32 (1981) 267--289.
17 N. Beffagna and G. Romani, Effects of two plasma- lemma A T P a s e inhibitors on H" extrusion and intracel- lular pH in Elodea densa leaves. J. Exp. Bot., 39 (1988) 1033-- 1043.
18 E. Marre, Fusicoccin: a tool in plant physiology. Annu. Rev. Plant Physiol., 30 (1979) 273--288.
19 20
21
22
C.T. Chen and C.H. Kao, Proton secretion in rice leaves. Bot. Bull. Acad. Sin., 29 (1988) 315--320. A. Bertl and H. Felle, Cytoplasmic pH of root hair cells of Sinapis alba recorded by a pH sensitive microelec- trode. Does fusicoccin stimulate the proton pump by cytoplasmic acidification? J. Exp. Bot., 36 (1985) 1142- 1149.
B. Brummer, A. Bertl, I. Potrykus, H. Felle and R.W. Parish, Evidence that fusicoccin and indole-3-acetic acid induced cytosolic acidification of Zea mays cells. Fed. Eur. Biochem. Soc., 189 (1985) 109-114.
A. Hager and I. Moser, Acetic acid esters and permea-
23
24
hie weak acids induce active proton extrusion and extension growth of coleoptile segments by lowering the cytoplasmic pH. Planta, 163 (1985) 391 - 400. J.K.M. Roberts, P.M. Ray, N. Wade-Jardetzky and O. Jardetzky, Extent of intracellular pH changes during H ÷ extrusion by maize root-tip cells. Planta, 152 (1981) 7 6 - 78.
S.F. Boggess and C.R. Stewart, The relationship between water stress induced proline accumulation and inhibition of protein synthesis in tobacco leaves. Plant Sci. Lett., 17 (1980~ 2 4 5 - 252.