Archives of Virology (2014) 000:000-000 Brief Report
Genetic and serological characterization of Chrysanthemum stem necrosis
virus, a species of the genus Tospovirus
Pei-Ru Wu1, Wan-Chu Chien1, Misturu Okuda2, Minoru Takeshita3, Shyi-Dong Yeh4,5, Yun-Chi
Wang1‚ and Tsung-Chi Chen1,6*
1 Department of Biotechnology, Asia University, Wufeng, Taichung 41354, Taiwan
2 Agricultural Research Center, National Agriculture and Food Research Organization, Kannondai,
Tsukuba, Ibaraki 305-8666, Japan
3 Laboratory of Plant Pathology, Faculty of Agriculture, Kyushu University, Hakozaki 6-10-1,
Higashi-ku, Fukuoka 812-8581, Japan
4 Department of Plant Pathology, National Chung Hsing University, Taichung 40227, Taiwan 5 Agricultural Biotechnology Center, National Chung Hsing University, Taichung 40227, Taiwan 6 Department of Medical Research, China Medical University Hospital, China Medical University,
Taichung 40402, Taiwan
*Corresponding author: T.-C. Chen
E-mail: [email protected] Tel: 886-4-23394362 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Abstract
Chrysanthemum stem necrosis virus (CSNV) is a tentative tospovirus species. In this study, the complete genomic sequence of the Japanese CSNV isolate TcCh07A was determined. The L RNA is 8960 nt long and encodes the 331.0 kDa RNA-dependent RNA polymerase. The M RNA is 4828 nt long and encodes the 34.1 kDa movement protein (NSm) and the 127.7 kDa glycoprotein precursor (Gn/Gc). The S RNA is 2949 nt long and encodes the 52.4 kDa silencing suppressor protein (NSs) and the 29.3 kDa nucleocapsid (N) protein. The N protein of CSNV-TcCh07A was purified from virus-infected plant tissues and used for production of a rabbit polyclonal antiserum (RAs) and a monoclonal antibody (MAb). Results of serological tests by indirect ELISA and western blotting using the prepared RAs and MAb, and a previously produced RAs against the N protein of Tomato spotted wilt virus (TSWV), indicate that CSNV-TcCh07A, TSWV, Tomato chlorotic spot virus, Groundnut ringspot virus, Alstroemeria necrotic streak virus and Impatiens necrotic spot virus are serologically related.
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Tospoviruses are persistently transmitted by thrips and cause significant losses in yield and quality of many economically important crops worldwide [21]. The genus Tospovirus, named from the type species Tomato spotted wilt virus (TSWV), is the only genus containing plant-infecting viruses in the family Bunyaviridae. The virion of tospoviruses is quasi-spherical in shape, 80-110 nm in diameter, with a characteristic lipid envelope and possesses a tripartite genome with large (L), medium (M) and small (S) ssRNAs. All three RNAs have a panhandle structure formed by base paring of complementary nucleotides (nt) at the 5'- and 3'-ends [13]. The L RNA is of negative sense and encodes an RNA-dependent RNA polymerase (RdRp) for
replication and transcription [27]. Both M and S RNAs are ambisense and contain two open reading frames (ORFs) in opposite directions that are flanked by an AU-rich intergenic region (IGR). The viral (v) sense of M RNA encodes a movement protein named NSm [14]. The viral complementary (vc) sense of M RNA encodes a precursor of two glycoproteins, Gn and Gc, which form spikes on the surface of the virion envelope [12], and which are also responsible for thrips transmission [29]. The v sense of S RNA encodes a nonstructural protein (NSs) that functions as a suppressor of RNA silencing [2, 25] and as an avirulence determinant against the natural resistance gene Tsw in Capsicum annuum [10]. The vc sense of S RNA encodes the nucleocapsid (N) protein that encapsidates genomic RNA molecules [8]. The sequence of the N gene is the most important criterion for demarcation of tospovirus species [13]. Classification of tospoviruses based on serological relationships of N proteins is practical in diagnosis and
inspection [5].
Chrysanthemum stem necrosis virus (CSNV), a tospovirus causing necrotic lesions
surrounded by yellow spots on leaves and necrosis on stems, peduncles and floral receptacles of chrysanthemum, was first found in Atibaia County, Brazil, in 1994 [1]. CSNV has also been reported in European and Asian countries, including the Netherlands [28], United Kingdom [18], Slovenia [22], Belgium [9], Japan [16] and China, and can also infect tomato [1], aster and Russell prairie gentian [17]. Frankliniella occidentalis and F. schultzei were identified as vectors of CSNV [20]. The sequences of the M and S RNAs of CSNV have been reported previously [19, 24, 26]. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
In this study, an isolate of CSNV denoted TcCh07A [26], collected from chrysanthemum in Tochigi Prefecture, Japan, was used for whole genome sequencing. Total RNA was extracted from leaf tissues of CSNV-TcCh07A-infected Nicotiana benthamiana plants using the Plant Total RNA Miniprep Purification kit (GMbiolab, Taichung, Taiwan) according to the manufacturer’s
instructions. Nucleotide sequences of tospovirus-degenerate and CSNV-specific primers used in this study are listed in Table S1. Reverse transcription (RT) was performed using 2 μg of total RNA mixed with 200 nM of individual primers and 25 U of Moloney murine leukemia virus (M-MLV) reverse transcriptase (GMbiolab). Reaction mixtures were incubated at 42°C for 60 min and then inactivated by incubation of 72°C for 15 min. Subsequently, cDNAs were mixed with 2.5 U of Ex Taq DNA polymerase (Takara, Shiga, Japan) and incubated at 94°C for 2 min for hot start. Polymerase chain reaction (PCR) was performed with 35 cycles of strand separation at 94°C for 30 sec, annealing at 50-60°C for 30 sec (depending on the Tm values of individual primers as indicated in Table S1), and synthesis at 72°C for 1 min. A final extension step at 72°C for 7 min was conducted. The amplified DNA fragments were analyzed by 1% agarose gel electrophoresis and then eluted from gels using the Micro-Elute DNA Clean/Extraction kit (GMbiolab) as per manufacturer’s instruction. All amplicons were cloned using the TOPO TA Cloning kit (Invitrogen, Carlsbad, CA) and the recombinant plasmids were transferred into E. coli DH5α competent cells. Three clones of each amplicon were selected for sequencing by an ABI3730XL DNA Analyzer (Perkin-Elmer Applied Biosystems, Foster City, CA) performed by Mission Biotech Company (Taipei, Taiwan).
The tospoviral 5'-terminal consensus sequence (5'-AGAGCAAU-3') and its complementary 3'-terminal sequence (5'-AUUGCUCU-3') of the L, M and S RNAs of CSNV-TcCh07A were verified by rapid amplification of cDNA ends (RACE) [15]. First strand cDNAs were synthesized by SuperScript® III reverse transcriptase (Invitrogen) and oligonucleotide PolyG(11g3a3g)
(5′-GGGGGGGGGGGAAAGGG-3′) was ligated at the 3'-end of cDNA molecules by terminal deoxynucleotidyl transferase (TdT). The tailed cDNA fragments were used as templates for PCR amplification by mixing with 2.5 U of Ex Taq DNA polymerase (Takara), the primer
PolyC(3c3t11c) (5′-CCCTTTCCCCCCCCCCC-3′) and another proper primer as shown in Fig. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
S1. Amplified fragments were cloned using the TOPO TA Cloning kit (Invitrogen) for sequencing.
The determined nt sequences of CSNV-TcCh07A were translated into amino acid (aa) sequences by the Sixframe program of Biology Workbench, San Diego Supercomputer Center (SDSC) (http://workbench.sdsc.edu/). The available sequences of individual tospoviruses (Table S2) were obtained from GenBank for sequence analyses. Percent identities of nt and aa sequences of CSNV-TcCh07A with those of other tospoviruses were calculated by the Gap program of SeqWeb (Accelrys Inc., San Diego, CA). Input data of multiple aa sequence alignments obtained from ClustalW (Biology Workbench, SDSC) were changed to the Phylip format for phylogenetic analyses conducted using the Phylip 3.68 package (University of Washington, Seattle, WA). Bootstrapping was performed from 1,000 repeats to generate multiple reassembled data sets by the Seqboot program. Distance matrices of aa sequences were produced by the Protdist program under PAM matrices of the Dayhoff model. Phylogenetic branches were set by the Neighbor program using the Neighbor-Joining method. Finally, phylogenetic trees were produced by the Consense program.
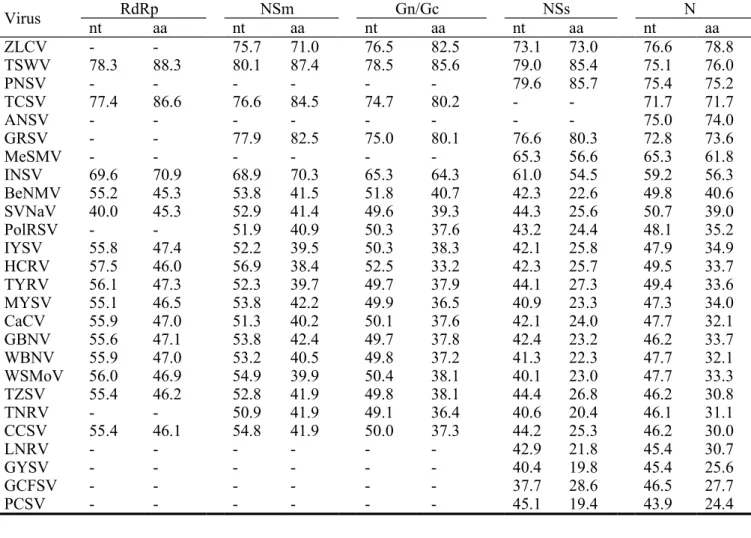
Our result showed that the full-length sequence of CSNV-TcCh07A L RNA contains 8960 nt in length (Accession No. KF493773), encoding a large ORF of 8625 nt for a RdRp of 2874 aa (331.0 kDa) (Fig. S1a). The 5'- and 3'-untranslatable regions (UTRs) contain 33 nt and 302 nt, respectively. The RdRp ORF of CSNV-TcCh07A shares higher nt and aa identities of 69.6-78.3% and 70.9-88.3%, respectively, with those of Impatiens necrotic spot virus (INSV), Tomato
chlorotic spot virus (TCSV) and TSWV; and nt and aa identities of 40.0-56.1% and 45.3-47.4%, respectively, with those of other reported tospoviruses (Table 1). Essential motifs, including motifs A (DxxKW, aa 1367-1371), B (QGxxxxxSS, aa 1455-1463), C (SDD, aa 1493-1495), D (K, aa 1540), E (ExxS, aa 1550-1553) and F (KxQxxxxxR, aa 1286-1294) are present in the deduced aa sequence (Fig. 1a). We notice that motifs A to E are identical within the RdRps of CSNV, TSWV, TCSV and INSV. The same is also observed for MYSV, CCSV, TZSV, CaCV, WSMoV, GBNV, WBNV, TYRV, IYSV and HCRV as well as in SVNaV and BeNMV. This suggests a co-evolutionary relationship within distinct tospovirus groups.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
The complete M RNA sequence of CSNV-TcCh07A is 4828 nt in length (Accession No. KF493772), consisting of two ORFs, NSm and Gn/Gc, in an opposite orientation separated by an AU-rich IGR (Fig. S1b). The sequences of the 5'-UTR, 3'-UTR and IGR of the M RNA consist of 101 nt, 83 nt and 324 nt, respectively. The NSm ORF is 912 nt in length, encoding a protein of 303 aa (34.1 kDa), and the Gn/Gc ORF is 3408 nt in length, encoding a glycoprotein precursor (GP) of 1135 aa (127.7 kDa). The NSm ORF shares 99.3% nt and aa identity with the original Brazilian isolate, and the Gn/Gc ORF shares 97.6% nt identity and 98.9% aa identity with the Brazilian Chry-1 isolate (Table S3). Additionally, the NSm ORF shares nt and aa identities of 68.9-80.1% and 70.3-87.4%, respectively, with those of Groundnut ringspot virus (GRSV), TCSV, TSWV, Zucchini lethal chlorosis virus (ZLCV) and INSV and relatively lower nt and aa identities of 50.9-54.9% and 38.2-42.4%, respectively, with those of other tospoviruses (Table 1). The Gn/Gc ORF shares nt and aa identities of 74.7-78.5% and 80.1-85.6%, respectively, with those of GRSV, TCSV, TSWV and ZLCV; 65.3% nt identity and 64.3% aa identity with that of INSV; and lower nt and aa identities of 38.7-51.8% and 32.0-40.7%, respectively, with those of other tospoviruses (Table 1).
The S RNA sequence of CSNV-TcCh07A was determined as being 2949 nt in length (Accession No. KF493771), consisting of NSs and N ORFs in opposite directions flanked by a 531 nt AU-rich IGR (Fig. S1c). The 5'- and 3'-UTRs of the S RNA are 79 nt and 152 nt long, respectively. The NSs ORF is 1404 nt in length encoding a 467 aa (52.4 kDa) protein, and the N ORF is 783 nt in length encoding a 260 aa (29.3 kDa) protein. The nt sequence of
CSNV-TcCh07A S RNA was compared to another Japanese isolate, HiCh06A L1 (Accession No.
AB600873) [26], indicating that these two CSNV isolates share a high degree of identity (98.2%). The NSs ORFs of both TcCh07A and HiCh06A L1 isolates share 97.9% and 91.3% nt and aa identity, respectively. In addition, the NSs protein of TcCh07A is 3-aa longer than that of HiCh06A L1. Different host response upon infection with TcCh07A and HiCh06A L1 has been previously reported [26]. Whether the NSs protein plays a role in host range determination remains to be further investigated. The N ORF of CSNV-TcCh07A shares 98.5-99.0% and 98.8-100% nt and aa identities, respectively, with those of different isolates of CSNV available in 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
GenBank (Table S3). The N ORF shares nt and aa identities of 71.7-76.6% and 71.7-78.8%, respectively, with those of ZLCV, TSWV, Pepper necrotic spot virus (PNSV) [6], Alstroemeria necrotic streak virus (ANSV), GRSV and TCSV; 59.2-65.3% and 56.3-61.8%, respectively, with those of Melon severe mosaic virus (MeSMV) and INSV; and 43.9-50.7% and 24.4-40.6%, respectively, with those of other known tospoviruses (Table 1). The NSs ORF shares 73.1-79.6% nt and 73.0-85.7% aa identities with those of PNSV, TSWV, GRSV and ZLCV; 61.0-65.3% nt and 54.5-56.6% aa identities with those of MeSMV and INSV; and 37.7-44.4% nt and 19.8-28.6% aa identities with those of other tospoviruses (Table 1).
Phylogenetic relationships based on the RdRp, Gn/Gc, NSm, NSs and N proteins showed that the current tospoviruses can be clustered into five evolutionary clades (Fig. 1b and Fig. S2). The first group is designated as the TSWV-related (T) clade. The second group is regarded as the WSMoV-related (W) clade. The third group is referred to as the GYSV-related (G) clade. The fourth group is denoted as the SVNaV-related (S) clade. Lisianthus necrotic ringspot virus (LNRV) [23] is distantly related to other tospoviruses and thus referred as the fifth (L) clade. Up to date, at least 27 tospovirus species have been characterized (Table 1) and most of them belong to the T and W clades.
The N protein of CSNV-TcCh07A was purified from virus-infected leaf tissues of Chenopodium quinoa plants following a previously described method [30]. The expected band corresponding to the N protein was visualized by preparative gel electrophoresis after the gel was submerged in cold (4°C) 0.25 M KCl solution. The N protein in sliced bands was eluted from the polyacrylamide gel using a Model 442 Electro-Eluter (Bio-Rad, Hercules, CA). The yield of purified N protein was estimated by the software Spot Density (AlphaInnotech Corporation, San Leandro, CA) by comparison with quantified bovine serum albumin (BSA).
One hundred and fifty micrograms of the purified N protein dissolved in 1 ml of phosphate buffered saline (PBS) were emulsified with an equal volume of Freund’s complete adjuvant (Difco Laboratories, BD, NJ) for subcutaneous injection of a New Zealand white rabbit.
Subsequently, the same amount of immunogen in 1 ml of PBS emulsified with an equal volume of Freund’s incomplete adjuvant (Difco Laboratories) was administered weekly into the immunized 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
rabbit for three weeks. The antiserum, denoted as RAs-CSNV-N, was collected weekly from the marginal veins of the ear for three months, starting one week after the fourth injection.
For preparation of a monoclonal antibody, 6 to 8-week-old BALB/cByJ mice were first immunized with 50 µg of the purified N protein (in 250 µl of PBS) emulsified with an equal volume of Freund’s complete adjuvant (Difco Laboratories) by intraperitoneal injection. The same amount of immunogen emulsified with Freund’s incomplete adjuvant (Difco Laboratories) was used for two additional intraperitoneal injections at weekly intervals. Mice were sacrificed three days after the fourth intraperitoneal injection without adding adjuvant to collect splenocytes that were then fused with Spll/0-ag/14 myeloma cells. Hybridoma cells were screened by indirect enzyme-linked immunosorbent assay (ELISA). The selected hybridoma cell lines were further cloned via limiting dilution. Finally, one stable hybridoma cell line (named S217D4C10) was obtained. The monoclonal antibody, denoted MAb-CSNV-N, was obtained from ascitic fluids of the Pristane-primed BALB/cByJ mice injected with S217D4C10 hybridoma cells.
The titer and sensitivity of RAs-CSNV-N and MAb-CSNV-N were determined by indirect ELISA as previously described [4]. Crude extracts of virus-infected plant tissues at a 1/50 dilution in coating buffer (50 mM Na2CO3, pH 9.6, containing 0.01% NaN3), or 10-fold serial dilutions (1
pg to 1 mg) of the purified N protein of CSNV-TcCh07A dissolved in coating buffer, were used to incubate with RAs-CSNV-N or MAb-CSNV-N at 37°C for 30 min. The alkaline phosphatase (AP)-conjugated goat anti-rabbit or anti-mouse IgGs (Jackson Immuno Research Laboratories, Inc., West Grove, PA) were diluted 1/5000 in conjugate buffer (PBST containing 2% PVP-40 and 0.2% ovalbumin) to react with RAs-CSNV-N and MAb-CSNV-N, respectively. The absorbance at 405 nm was recorded by a Model 680 microplate reader (Bio-Rad) 1-2 hr after the addition of enzyme substrate ρ-nitrophenyl phosphate disodium hexahydrate (ρ-NPP-Na) (GMbiolab). Results showed that the endpoint dilution of RAs-CSNV-N was 1/32000. When RAs-CSNV-N was used at a 10-3 dilution, the sensitivity was determined as 10 ng of purified N protein. The
endpoint dilution of MAb-CSNV-N was determined as 10-6, and 10-4-diluted MAb-CSNV-N was
used in further assays. One nanogram of purified CSNV N protein was detected by MAb-CSNV-N at the 10-4 dilution. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
Western blotting was also conducted for serological assays of RAs-CSNV-N and MAb-CSNV-N following previously published procedures [11]. Leaves of virus-infected N.
benthamiana plants were ground in protein sample buffer (12.5 mM Tris-HCl, pH 6.8, 10% glycerol, 2% SDS, 2% β-mercaptoethanol and 0.001% bromophenol blue) at a 1/50 dilution. RAs-CSNV-N and MAb-RAs-CSNV-N were used at 10-3 and 10-4 dilutions, respectively. A rabbit antiserum
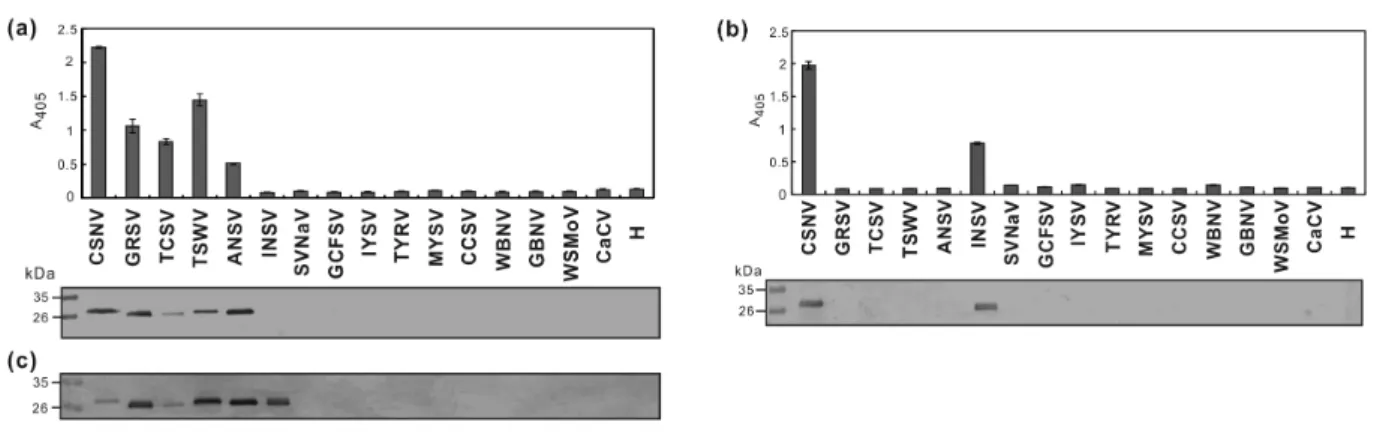
against the bacterial-expressed N protein of TSWV, denoted RAs-TSWV-N [3], was used at a 1/4000 dilution for comparison. The corresponding secondary antibodies were used at a 1/5000 dilution. Tospoviruses including ANSV, CSNV-TcCh07A, CaCV, CCSV, GBNV, GCFSV, GRSV, INSV, IYSV, MYSV, SVNaV, TCSV, TSWV, TYRV, WBNV and WSMoV were used to test the response of RAs-CSNV-N and MAb-CSNV-N. All tospoviruses were single lesion isolated and identified by RT-PCR using individual species-specific primer pairs to validate the serological results (data not shown). RAs-CSNV-N positively reacted with ANSV, GRSV, TCSV and TSWV as well as CSNV-TcCh07A in indirect ELISA and western blotting (Fig. 2a).
Similarly, RAs-TSWV-N [3] positively reacted with ANSV, CSNV-TcCh07A, GRSV, INSV and TCSV as well as TSWV in western blotting (Fig. 2a). Noteworthily, MAb-CSNV-N positively reacted not only with CSNV-TcCh07A but also with INSV in both indirect ELISA and western blotting (Fig. 2b). Sequence alignment revealed that a consensus sequence “YANILKAC” is present in the N proteins of CSNV, ZLCV and INSV. Whether this is the epitope recognized by MAb-CSNV-N will be further investigated.
CSNV seriously damages chrysanthemum, which is one of the most economically important ornamentals worldwide. Development of serological tools for virus diagnosis and detection allows researchers and farmers to inspect and prevent the disease caused by CSNV. Both RAs-CSNV-N and MAb-CSNV-N were used to detect CSNV infection of diseased chrysanthemum samples collected from the field in Japan. Results showed that positive reactions can be obtained when both RAs-CSNV-N and MAb-CSNV-N are tested against chlorotic tissues, but not against asymptomatic tissues. Virus particles present in a 100-fold dilution of the sap from symptomatic chrysanthemum tissue can be easily detected by RAs-CSNV-N. The use of MAb-CSNV-N increases the detection sensitivity up to a 1000-fold dilution of the sap (Fig. S3). Thus, sampling 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
from chlorotic tissues coupled with detection using MAb-CSNV-N is recommended for CSNV detection. RAs-CSNV-N and MAb-CSNV-N were also used for a field survey in the
chrysanthemum growing farms of central Taiwan, and no CSNV-infected sample was detected. Although CSNV has not yet been found in Taiwan, one of its insect vectors, F. schultzei, is present. In addition, Taiwan neighbors with Japan and China where CSNV occurs, implying a high risk of introduction. Since chrysanthemum is an important cut flower in Taiwan for exporting to Japan, prevention of CSNV occurrence is highly relevant.
Here, we report for the first time the complete genomic sequence of a CSNV isolate, and propose that tospoviruses can be divided into five evolutionary groups. Moreover, our findings confirm previous results of CSNV antibodies cross reacting with TSWV, TCSV and MeSMV [7], and extends the serological relationship of CSNV with GRSV, ANSV and INSV, as reciprocally verified by RAs-TSWV-N [3] (Fig. 2). Serological relationships among tospoviruses generally
match with phylogenetic grouping. Verification of serological relatedness among tospoviruses is helpful for prompt detection of tospovirus infections in field.
Acknowledgments
We thank the virus providers: Dr. Dennis Gonsalves for GRSV; Dr. James W. Moyer for INSV; Dr. Richard Kormelink for ANSV, IYSV and TYRV; Dr. Prem A. Rajagopalan for GBNV and WBNV; Dr. Ioannis E. Tzanetakis for SVNaV; the DSMZ Plant Virus Collection, Germany for TCSV; and the permission of Bureau of Animal and Plant Health Inspection and Quarantine, Council of Agriculture, Executive Yuan for virus import. We also thank Dr. Lei Wan for
providing myeloma cells. Funding was provided by the National Science Council (NSC92-2317-B-005-024, NSC-101-2911-I-005-301 and NSC-102-2911-I-005-301) and the Ministry of Education, Taiwan, R.O.C. under the ATU plan for study.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
References
1. Bezerra IC, Resende RdeO, Pozzer L, Nagata T, Kormelink R, de Avila AC (1999) Increase of tospoviral diversity in Brazil with the identification of two new tospovirus species, one from chrysanthemum and one from zucchini. Phytopathology 89:823-830
2. Bucher E, Sijen T, de Haan P, Goldbach R, Prins M (2003) Negative-strand tospoviruses and tenuiviruses carry a gene for a suppressor of gene silencing at analogous genomic positions. J Virol 77:1329-1336
3. Chen SM, Wang YC, Wu PR, Chen TC (2014) Production of antiserum against the nucleocapsid protein of Tomato spotted wilt virus and investigation of its serological relationship with other tospoviruses. Plant Prot Bull 56:55-74
4. Chen TC, Hsu HT, Jain RK, Huang CW, Lin CH, Liu FL, Yeh SD (2005) Purification and serological analyses of tospoviral nucleocapsid proteins expressed by Zucchini yellow mosaic virus vector in squash. J Virol Methods 129:113-124
5. Chen TC, Lu YY, Cheng YH, Li JT, Yeh YC, Kang YC, Chang CP, Huang LH, Peng JC, Yeh SD (2010) Serological relationship between Melon yellow spot virus and Watermelon silver mottle virus and differential detection of the two viruses in cucurbits. Arch Virol 155:1085-1095
6. Cheng YH, Zheng YX, Tai CH, Yen JH, Chen YK, Jan, FJ (2014) Identification,
characterisation and detection of a new tospovirus on sweet pepper. Ann Appl Biol 164:107-115
7. Ciuffo M, Kurowski C, Vivoda E, Copes B, Masenga V, Falk BW, Turina M (2009) A new Tospovirus sp. in cucurbit crops in Mexico. Plant Dis 93:467-474
8. de Haan P, Wagemakers L, Peters D, Goldbach R (1990) The S RNA segment of tomato spotted wilt virus has an ambisense character. J Gen Virol 71:1001-1007
9. De Jonghe K, Morio S, Maes M (2013) First outbreak of Chrysanthemum stem necrosis virus (CSNV) on potted Chrysanthemum in Belgium. New Disease Reports 28:14
10. de Ronde D, Butterbach P, Lohuis D, Hedil M, van Lent JWM, Kormelink R (2013) Tsw 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
gene-based resistance is triggered by a functional RNA silencing suppressor protein of the Tomato spotted wilt virus. Mol Plant Pathol 14:405-415
11. Gooderham K (1984) Transfer techniques in protein blotting. In: Walker JM (ed) Methods in Molecular Biology. Vol. 1. Proteins. Humana Press, Clifton, NJ, pp 165-178
12. Kikkert M, van Lent J, Storms M, Bodegom P, Kormelink R, Goldbach R (1999) Tomato spotted wilt virus particle morphogenesis in plant cells. J Virol 73:2288-2297
13. King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (2012) Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses. Elsevier Academic Press,
Amsterdam, pp 725-741
14. Lewandowski DJ, Adkins S (2005) The tubule-forming NSm protein from Tomato spotted wilt virus complements cell-to-cell and long-distance movement of Tobacco mosaic virus hybrids. Virology 342:26-37
15. Li JT, Yeh YC, Yeh SD, Raja JAJ, Rajagopalan PA, Liu LY, Chen TC (2011) Complete genomic sequence of watermelon bud necrosis virus. Arch Virol 156:359-362
16. Matsuura S, Kubota K, Okuda M (2007) First report of Chrysanthemum stem necrosis virus on chrysanthemum in Japan. Plant Dis 91:468
17. Momonoi K, Moriwaki J, Morikawa T (2011) Stem necrosis of aster and Russell prairie gentian caused by Chrysanthemum stem necrosis virus. J Gen Plant Pathol 77:142-146 18. Mumford RA, Jarvis B, Morris J, Blockley A (2003) First report of Chrysanthemum stem
necrosis virus (CSNV) in the UK. Plant Pathol 52:779
19. Nagata T, Carvalho KR, Sodré RdeA, Dutra LS, Oliveira PA, Noronha EF, Lovato FA, Resende RdeO, Oliveira de Avila AC, Inoue-Nagata AK (2007) The glycoprotein gene of Chrysanthemum stem necrosis virus and Zucchini lethal chlorosis virus and molecular relationship with other tospoviruses. Virus Genes 35:785-793
20. Okuda S, Okuda M, Matsuura S, Okazaki S, Iwai H (2013) Competence of Frankliniella occidentalis and Frankliniella intonsa strains as vectors for Chrysanthemum stem necrosis virus. Eur J Plant Pathol 136:355-362
21. Pappu HR, Jones RA, Jain RK (2009) Global status of tospovirus epidemics in diverse 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
cropping systems: Successes gained and challenges that lie ahead. Virus Res 141:219-236 22. Ravnikar M, Vozelj N, Mavrie I, Svigelj SD, Zupaneie M, Petrovie N (2003) Detection of
Chrysanthemum stem necrosis virus and Tomato spotted wilt virus in chrysanthemum. Abstracts, 8th International Congress of Plant Pathology (ICPP), Christchurch, New Zealand 23. Shimomoto Y, Kobayashi K, Okuda M (2014) Identification and characterization of
Lisianthus necrotic ringspot virus, a novel distinct tospovirus species causing necrotic disease of lisianthus (Eustoma grandiflorum). J Gen Plant Pathol 80:169-175
24. Silva MS, Martins CR, Bezerra IC, Nagata T, de Avila AC, Resende RO (2001) Sequence diversity of NS(m) movement protein of tospoviruses. Arch Virol 146:1267-1281
25. Takeda A, Sugiyama K, Nagano H, Mori M, Kaido M, Mise K, Tsuda S, Okuno T (2002) Identification of a novel RNA silencing suppressor, NSs protein of Tomato spotted wilt virus. FEBS Lett 532:75-79
26. Takeshita M, Nagai N, Okuda M, Matsuura S, Okuda S, Furuya N, Tsuchiya K (2011) Molecular and biological characterization of Chrysanthemum stem necrosis virus isolates from distinct regions in Japan. Eur J Plant Pathol 131:9-14
27. van Knippenberg I, Goldbach R, Kormelink R (2002) Purified Tomato spotted wilt virus particles support both genome replication and transcription in vitro. Virology 303:278-286 28. Verhoeven JThJ, Roenhorst JW, Cortes I, Peters D (1996) Detection of a novel tospovirus in
chrysanthemum. Acta Hortic 432:44-51
29. Whitfield AE, Kumar NKK, Rotenberg D, Ullman DE, Wyman EA, Zietlow C, Willis DK, German TL (2008) A soluble form of the Tomato spotted wilt virus (TSWV) glycoprotein GN
(GN-S) inhibits transmission of TSWV by Frankliniella occidentalis. Phytopathology
98:45-50
30. Yeh SD, Chao CH, Cheng YH, Chen CC (1996) Serological comparison of four distinct tospoviruses by polyclonal antibodies to purified nucleocapsid proteins. Acta Hortic 431:122-134 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Table 1. Nucleotide (nt) and amino acid (aa) identities (%) of the individual coding
sequences of Chrysanthemum stem necrosis virus (CSNV) TcCh07A isolate compared with those of other tospoviruses.
Virus nt RdRpaa nt NSmaa nt Gn/Gcaa nt NSsaa nt N aa ZLCV - - 75.7 71.0 76.5 82.5 73.1 73.0 76.6 78.8 TSWV 78.3 88.3 80.1 87.4 78.5 85.6 79.0 85.4 75.1 76.0 PNSV - - - 79.6 85.7 75.4 75.2 TCSV 77.4 86.6 76.6 84.5 74.7 80.2 - - 71.7 71.7 ANSV - - - 75.0 74.0 GRSV - - 77.9 82.5 75.0 80.1 76.6 80.3 72.8 73.6 MeSMV - - - 65.3 56.6 65.3 61.8 INSV 69.6 70.9 68.9 70.3 65.3 64.3 61.0 54.5 59.2 56.3 BeNMV 55.2 45.3 53.8 41.5 51.8 40.7 42.3 22.6 49.8 40.6 SVNaV 40.0 45.3 52.9 41.4 49.6 39.3 44.3 25.6 50.7 39.0 PolRSV - - 51.9 40.9 50.3 37.6 43.2 24.4 48.1 35.2 IYSV 55.8 47.4 52.2 39.5 50.3 38.3 42.1 25.8 47.9 34.9 HCRV 57.5 46.0 56.9 38.4 52.5 33.2 42.3 25.7 49.5 33.7 TYRV 56.1 47.3 52.3 39.7 49.7 37.9 44.1 27.3 49.4 33.6 MYSV 55.1 46.5 53.8 42.2 49.9 36.5 40.9 23.3 47.3 34.0 CaCV 55.9 47.0 51.3 40.2 50.1 37.6 42.1 24.0 47.7 32.1 GBNV 55.6 47.1 53.8 42.4 49.7 37.8 42.4 23.2 46.2 33.7 WBNV 55.9 47.0 53.2 40.5 49.8 37.2 41.3 22.3 47.7 32.1 WSMoV 56.0 46.9 54.9 39.9 50.4 38.1 40.1 23.0 47.7 33.3 TZSV 55.4 46.2 52.8 41.9 49.8 38.1 44.4 26.8 46.2 30.8 TNRV - - 50.9 41.9 49.1 36.4 40.6 20.4 46.1 31.1 CCSV 55.4 46.1 54.8 41.9 50.0 37.3 44.2 25.3 46.2 30.0 LNRV - - - 42.9 21.8 45.4 30.7 GYSV - - - 40.4 19.8 45.4 25.6 GCFSV - - - 37.7 28.6 46.5 27.7 PCSV - - - 45.1 19.4 43.9 24.4 1 2 3 4 5 6
Fig. 1 Analyses of the RNA-encoded RNA-dependent RNA polymerases (RdRps)
and nucleocapsid (N) proteins of tospoviruses. (a) Conserved motifs of RdRps of
tospoviruses are aligned. Consensus sequences within individual motifs among
members of the family Bunyaviridae are indicated. Accession codes of the analyzed
sequences of tospoviruses are listed in Table S2. The phleboviruses Uukuniemi virus
2 1 2 3 4 5 6
(Uukuniemi, D10759) and Rift Valley fever virus (RVFV, X564664), the bunyavirus
Bunyamwera virus (Bunyamwera, X14383), and the hantavirus Hantaan virus (Hanta, X55901) were used for comparison. (b) Phylogenetic dendrogram of tospoviruses
based on the N protein using the Neghbour-Joining algorithm with 1000 bootstrap
replicates. Viruses with a close relationship are circled.
2 1 2 3 4 5
Fig. 2 Serological reactions of rabbit polyclonal antiserum RAs-CSNV-N (a) and
monoclonal antibody MAb-CSNV-N (b) with different tospoviruses. Crude leaf
extracts of Nicotiana benthamiana plants infected with individual tospoviruses
(CSNV-TcCh07A, GRSV, TCSV, TSWV, ANSV, INSV, SVNaV, GCFSV, IYSV,
TYRV, MYSV, CCSV, WBNV, GBNV, WSMoV and CaCV; full virus names are
listed in Table S2) were used for indirect enzyme-linked immunosorbent assays (top
panel) and western blotting (lower panel). (c) Antiserum against the N protein of
TSWV (RAs-TSWV-N) [3] was used for comparison in western blotting. An
uninfected N. benthamiana plant (H) was used as negative control.
2 1 2 3 4 5 6 7 8 9 10 11