生物學報 (2007) 42(1): 47-53
茯苓表現序列基因庫之建構與分析
吳宗勳 孫智雯
* 國立臺灣師範大學生命科學系 (收稿日期:2006.10.13,接受日期:2006.10.26) 摘 要 茯苓是一種在中藥方劑裡應用範圍最廣的真菌,因其具有抗腫瘤與增進免疫力的功能。然而, 其生物活性成份有關知識仍有限,且基因體研究尚在起步階段。因此,本研究欲自茯苓菌絲體建立 一個茯苓表現序列標誌(EST)基因庫,並從中尋找未來具有醫藥應用潛力的蛋白質。在分析液態 培養茯苓菌絲體 EST 基因庫的過程中,我們發現其 16%的基因參與代謝作用、21%的基因參與遺 傳訊息傳遞(其中 10 %為核醣體蛋白基因)、7%的基因參與環境訊息傳遞、3%的基因參與細胞程 序、53%為未知功能的基因。此外,有一個與樟芝免疫調節蛋白 Aca1 胺基酸序列相似度達 50%的 基因(WcFIP1),推測此蛋白質可能是一種茯苓免疫調節蛋白。 關鍵詞:茯苓、表現序列標誌基因庫、真菌免疫調節蛋白緒 言
世界上有多種蕈類(mushroom)可供食用或 應用於醫學。近代科學研究發現它們含有的固醇 類、三萜類、蛋白質、核酸、多醣等成分可能有 藥用活性或潛力,因而具有抗腫瘤、免疫調節、 調節血壓、調節血脂血糖、抗自由基、抑制血小 板凝集、抗病毒、抗細菌、抗寄生蟲、調節腎功 能、保肝、調節神經或緩和長期性支氣管炎等生 理功能,其中抗腫瘤與增進免疫力功能最受矚目 (Wasser and Weis, 1999)。蕈類萃取物可提高免疫細胞的活性,促進細 胞激素與干擾素的生成(Lee et al., 1995)。也可 以促進自體免疫反應與活化免疫系統,提高巨噬 細胞吞噬能力,活化嗜中性球、自然殺手細胞、 T 細胞及 B 細胞(Hibino et al., 1994)。所以能調 節免疫系統,殺死腫瘤細胞或抑制其生長。近年 研究發現猪苓(Polyporus umbellatus)萃取物能 抑制癌細胞增生、抗 B 型肝炎,並能活化小鼠腹 腔巨噬細胞(Ohsawa et al., 1992; Yan, 1988; Zhang et al., 1991)。而舞菇(Grifola frondosa) 多 醣 亦 被 發 現 具 有 抗 腫 瘤 與 免 疫 調 節 活 性 (Takeyama et al., 1987;Inoue et al., 2002)。另 外,雲芝(Trametes versicolor)萃取物有抗 HIV 病毒活性(Collins and Ng, 1997);而其免疫調節 與抗腫瘤活性更是受到重視(Liu et al., 1993; Kobayashi et al., 1994)。目前亞洲市場也已有 PSK ( polysaccharide K )、 PSP ( polysaccharo- peptide )等多種雲芝多 醣蛋白商品 問世( Ng, 1998)。 早期的研究認為,在蕈類的萃取物中含有大 量的多醣類,很可能是抗腫瘤與調節免疫功能的 主要活性成分(Adachi et al., 1987)。但是多醣結 構不易鑑定,科學上至今難以確認蕈類中具生理 活性多醣的確實結構、分子量、衍生結構等,且 鑑別不同蕈類的多醣成分亦十分困難。而且,早 期蕈類多醣製備,大部分是以有機溶劑(例如乙 醇)加入子實體萃取液或菌絲體發酵液,直接沉 澱出蕈類多醣。此一粗萃法也會將萃取液中的蛋 白質沉澱出來,得到多醣與蛋白質的混合物;加 上後續的層析分離步驟主要依據不同分子量的 多醣進行分離,很難破壞多醣體與蛋白質間的鍵 結。因此,早期研究探討蕈類抗腫瘤功能時所採 用的多醣樣品有部分可能是多醣與蛋白質的混 合物;也就是說,蕈類多醣的生理機能很可能有 部分是偶合在多醣上的蛋白質所貢獻,且多醣體 所偶合的免疫調節蛋白可能對多醣體的抗腫瘤 功能可能非常重要。 除了凝集素(lectin)之外,蕈類中的真菌免 疫調節蛋白亦能調節免疫功能。研究人員在靈芝 (Ganoderma lucidum)萃取液中發現一種分子量 約 17 kDa 的蛋白質,命名為 Ling Zhi-8(LZ-8),
並已完成蛋白質定序與基因選殖(Tanaka, 1989; Murasugi, 1991)。LZ-8 可凝集羊的紅血球,但對 人類紅血球無凝集反應,與凝集素的定義不盡相 符,另歸類為真菌免疫調節蛋白。體外試驗(in vitro)發現 LZ-8 能促進 T 細胞分裂及分泌細胞 激素 IL-2、IFN-γ、TNF-α、ICAM-1,並提高細 胞黏著分子CD2 與 CD11b 的表現,以及內皮細
胞IFN-γ 的表現(Kino et al., 1991)。
目前也已經找到蛋白質序列與 LZ-8 類似的 金針菇免疫調節蛋白(FIP-fve)、草菇免疫調節 蛋 白 (FIP-vvo ) 與 松 杉 靈 芝 免 疫 調 節 蛋 白 (FIP-gts),並已分別完成基因選殖。三種蛋白 質皆可凝集兔子的紅血球,並具有抑制過敏反 應、促進T 細胞增殖或分泌細胞激素、減緩由牛 血清白蛋白引起的全身性休克、降低發炎物誘發 的水腫等免疫調節功能(Ko et al., 1995; Hsu et al., 1997; Lin et al., 1997 )。 最 近 幾 年 , 樟 芝 (Taiwanofungus camphoratus)也發現具抗氧化 與 保 肝 功 能 、 及 調 節 小 鼠 免 疫 功 能 的 蛋 白 質 (Hseu et al., 2002),並選殖出一個具有免疫調節 功能的樟芝蛋白基因(錢, 2004)。至於其他蕈類 是否具有免疫調節功能的蛋白質,值得進一步研 究。 茯苓是一種在中藥方劑裡應用範圍最廣的 真菌,藥理研究亦證實其茯苓萃取物具有消炎、 免疫調節、止吐、抗氧化、抗腫瘤、鎮靜、利尿、 降血糖等作用(閔, 1996)。茯苓的三萜類可抑制 癌細胞增生(Kaminaga et al., 1996),多種茯苓菌 株產生之多醣均具有抗腫瘤活性,其衍生物 U-茯苓多糖或羥乙基茯苓多醣可抑制小鼠肉瘤細 胞生長、調節免疫系統功能(Jin et al., 2003)。但 是茯苓免疫調節蛋白質研究,直到最近才從茯苓 子 實 體 中 分 離 出 一 種 茯 苓 蛋 白 ( Poria cocos protein, PCP),能活化巨噬細胞 RAW264.7 分泌 TNF-α 及一氧化氮、誘導多種細胞激素 mRNA 表 現、及調節小鼠脾臟細胞的功能(張, 2005),這 顯示茯苓亦含有能調節哺乳類免疫調節功能的 蛋白質。然而,表現此茯苓蛋白的基因為何?與 其他已知的基因或蛋白質是否相似?這些問題 皆有待更深入的探討。因此在本研究中,我們建 構了一個茯苓表現序列標誌(EST)基因庫,除 了分析茯苓細胞內能被表現的基因類別外,並從 中探討未來具有醫藥應用潛力的蛋白質。
材料與方法
茯苓菌絲體 茯苓菌絲體購自食品工業發展研究所,編號 BCRC 36022 (ATCC 42588)。固態及液態培養 茯苓菌絲體則採用食品工業發展研究所建議之 MALT 培養基(財, 2000),在 26 ℃生長箱中恆 溫培養。液態培養的速率震盪為215 rpm 培養 1~4 週。茯苓菌絲體 total RNA/ poly (A+
) RNA 的萃取及 cDNA 的合成
Total RNA 是使用 TRIzol 試劑(Invitrogen) 萃取。然後以分光光度儀(Eppendorf)測定 OD260
及 OD280讀值(OD260/OD280比値低於 1.4 則丟
棄),以公式OD260×40×稀釋倍數計算出 total RNA
濃度。菌絲體 poly (A+) RNA 的萃取,是使用
QIAGEN 的 Oligotex® mRNA mini kit,萃取過程
乃根據廠商建議之程序。為期能獲得較長 cDNA
序列,利用BD SMARTTM PCR cDNA synthesis kit
合成cDNA,再建立茯苓 EST 基因庫。取 0.5 μg 的一週齡茯苓菌絲體poly (A+) RNA,並參照廠商 建議之程序,以反轉錄酶合成cDNA。再以專一 於茯苓肌動蛋白之引子wcActin-F2 (5´-GAAGAT GACGCAGATCATGTTC-3) 及 wcActin-R2(5´-T CTTAATCAGGAACTCGGTGAG-3´),進行聚合 酶連鎖反應(polymerase chain reaction,PCR),
經由PCR 產物的定量,來確認 cDNA 的品質。 茯苓表現序列標誌基因庫的建構 以BD PCR Primer IIA(5´-AAGCAGTGGTA TCAACGCAGAGT-3´)作引子、1 μL 上述 cDNA 作模版,進行PCR 反應(過程為 94 ℃ 15 秒、65 ℃ 30 秒、68 ℃ 6 分,進行 15 循環),擴增cDNA 量。PCR 產物經純化後,將其黏結至 pGEM-T 載 體(Promega),再以熱休克法轉形至大腸桿菌 DH5α 中(Ecos-101, Yeastern),再篩選單一菌落。 最後利用 SP6 及 T7 引子對(此二者為 pGEM-T 載體中可用來檢驗插入序列長度的引子),以 PCR 檢驗插入序列的 DNA 長度(過程為 94 ℃ 15 秒、55 ℃ 30 秒、72 ℃ 2 分,進行 25 循環)。DNA 長度大於500 bp 者,進行定序反應。 基因序列分析比對 所得定序結果中來自同一菌落所得序列,利 用Vector NTI 8 先拼接(contig)為單一序列、解 譯出可能的蛋白質序列,比對出重覆序列。其中
Wolfiporia EST Library BLAST 比對,若比對錯誤期望值(expect)小於 1×10-5或蛋白質序列相似度超過 50%,則試圖由 既存的資料庫中的已知蛋白質的功能,推測解譯 的 蛋 白 質 可 能 的 功 能 。 此 外 , 以 PSORT II ( http://psort.ims.u-tokyo.ac.jp/ ) 與 TargetP (http://www.cbs.dtu.dk/services/SignalP/)網站分 析蛋白質在細胞的區位,或利用 InterProScan (http://www.ebi.ac.uk/InterProScan/)網站分析蛋 白質胺基酸序列中是否存在與其他物種已知蛋 白質保守區(conserved domain)相似的序列,藉 以推測這些茯苓蛋白質的功能。最後參考 KEGG ( Kyoto Encyclopedia of Genes and Genomes, Kanehisa et al., 2004)的分類系統,並逐一整理 列表。
結果及討論
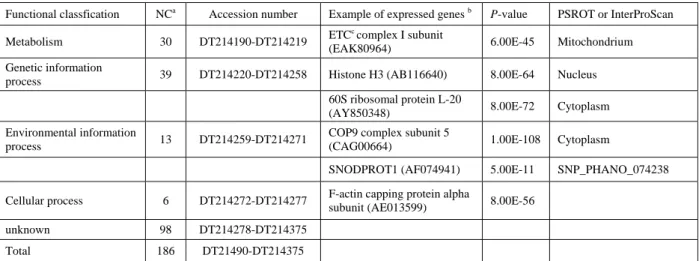
茯苓表現序列標誌基因庫的分析 從茯苓的 cDNA 基因庫中,我們挑選出 509 個單一菌落,以 PCR 及瓊脂凝膠電泳分析來檢驗 插入載體的 cDNA 的長度,發現其中 495 個菌落 含有插入的 cDNA 片段,而且 209 個菌落的插入 cDNA 序列長度大於 500 bp,此表示這個茯苓 cDNA 基因庫的有效菌落高達 42.2%。經定序反 應後,將此 209 個菌落的插入 cDNA 序列,送至 NCBI 網站上進行 BLAST 比對。扣除重複者,共 獲得 186 個不同的 cDNA 序列,並送至 dbEST 資料庫(Boguski et al., 1993),得到 186 條相對 應的序列登錄號碼(accession number)DT214190 至 DT214375。其中 53%為未知基因,另有一條 是 rRNA 序列。接著以 InterProScan 、PSORT II 分析蛋白質胺基酸序列保守區與區位,並推測這 些茯苓蛋白質可能的功能(表一) ,再利用 KEGG 網站的蛋白質功能性分類區分序列的功能(圖 一 )。 在 已 知 基 因 的 序 列 中 , 參 與 代 謝 (metabolism)作用的有 30 個(佔全部 16%)基 因,遺傳訊息傳遞(genetic information process) 功能有 39 個(21%)基因,環境訊息傳遞(environ- mental information process)功能有 13 個(7%)圖一、茯苓基因體之表現蛋白的分類及百分比。
Figure 1. Types and percentage of proteins encoded
in the genome of Wolfiporia cocos. 表一、一週齡茯苓之 cDNA 基因庫的功能分類。
Table 1. Functional classification of 1-week-old cDNA library in Wolfiporia cocos.
Functional classfication NCa Accession number Example of expressed genes b P-value PSROT or InterProScan
Metabolism 30 DT214190-DT214219 ETC
c complex I subunit
(EAK80964) 6.00E-45 Mitochondrium
Genetic information
process 39 DT214220-DT214258 Histone H3 (AB116640) 8.00E-64 Nucleus
60S ribosomal protein L-20
(AY850348) 8.00E-72 Cytoplasm
Environmental information
process 13 DT214259-DT214271
COP9 complex subunit 5
(CAG00664) 1.00E-108 Cytoplasm
SNODPROT1 (AF074941) 5.00E-11 SNP_PHANO_074238
Cellular process 6 DT214272-DT214277 F-actin capping protein alpha
subunit (AE013599) 8.00E-56
unknown 98 DT214278-DT214375
Total 186 DT21490-DT214375
a
Number of clones.
b
Accession number of hit gene is shown in parentheses.
c
基因,細胞程序(cellular process)功能有 6 個(3%) 基因。代謝基因包含了參與一次代謝(primary metoblism)、二次代謝(second metoblism)、電 子 傳 遞 鏈 ( electron transfer chain )、 能 量 (energy)、及光合作用(photosynthesis)者。遺 傳 訊 息 傳 遞 基 因 包 含 了 參 與 染 色 絲 結 構 ( chromatin structure )、 DNA 複 製 ( DNA replication)、 DNA 修復( DNA repair)、轉錄 (transcription)、轉譯(translation)、及轉譯後修 飾(post-translational modification)者。19 個核 醣體蛋白(ribosomal protein)是包括在遺傳訊息 傳遞基因中。環境訊息傳遞基因包含了參與運輸 蛋 白 ( membrane transporter )、 胞 間 訊 息 傳 遞 (intracellular communication)、及細胞壁(cell wall)者。細胞程序基因包含了參與細胞骨骼 (cytoskeleton)、細胞週期(cell cycle)、及胞內 蛋白運輸(cellular sorting)者。 目前有許多物種的基因體已被解序,包含多 種病毒、細菌、酵母菌、果蠅、植物、及動物。 這些物種的基因體大小從數千到數百億核苷酸 鹼基,所含的基因數目從數百到數萬個,但對各 物種的基因作功能性分析時,結果皆極為類似。 幾乎所有物種的表現序列,大約 50%是未知功能 基因,15~20%為代謝功能基因,15-20%為遺傳 訊息傳遞基因,5~10%為環境訊息傳遞基因,5% 為細胞程序基因。即使是已知功能的基因產物, 其調控以及相互作用的機制都尚不清楚。因此, 近年來基因體研究的重心已由定序逐漸轉變為 基因序列之解讀,透過生化、物理、化學、分子 生物、高速電腦計算等技術,探討基因的表現與 功能之「功能性基因體學」特定網站逐漸出現。 例如:病毒 (http://athena.bioc.uvic.ca/pbr/vgo)、 酵 母 菌 ( http://genome-www.stanford.edu/ Saccharomyces)、果蠅(http://flybase.bio.indiana. edu)、人類及老鼠(http://www.genome.gov 及 http://www.ncbi.nlm.nih.gov ) 、 阿 拉 伯 芥 ( http://www.arabidopsis.org ) 、 稻 米 (http://www.rice-research.org)。這些網站提供了 大量基因的資訊以供比對及推測許多其他物種 未知基因的功能。至於茯苓雖被廣泛應用為食物 及藥物,但在此研究前整個 NCBI 網站上被登錄 的茯苓的核苷酸序列只有 10 條,而且皆為與分 類 有 關的 細胞 質 型或 粒線 體型 rRNA 與 ITS (internal transcribed spacer)序列。因此,藉由 本研究所建立的基因庫可作為茯苓功能基因體 研究的初步基礎。
圖二、樟芝蛋白 Aca1 與茯苓免疫調節蛋白 WcFIP1 的胺基酸序列比對。
Figure 2. Alignment of Aca1 and WcFIP1 peptide
sequences. 茯苓免疫調節蛋白 WcFIP1 在茯苓液態培養一週齡菌絲體的 cDNA 基因 庫中,發現一個基因 WcFIP1 所解譯出的預測胺 基酸序列與樟芝免疫調節蛋白(Aca1)胺基酸序 列比對,相似度達 50%(圖二)。Aca1 為樟芝菌 絲體中的一種醣蛋白,分子量為 29 kDa,等電點 為 5.5。而 WcFIP1 在蛋白質分析軟體中預估的分 子量為 14.6 kDa,等電點為 4.0,似乎與 Aca1 不 同。但是,以西方免疫轉漬法分析 WcFIP1 與 Aca1 的表現,發現表現於大腸桿菌的 WcFIP1 與 Aca1 重組蛋白皆可被茯苓蛋白 PCP 單株抗體辨識 (data not shown)。由於 WcFIP1 與 Aca1 之間存 在特定序列或結構部份的相似性,而且 Aca1 具 有免疫調節活性,也許 WcFIP1 亦具有免疫調節 活性,但尚待進一步研究證實。另外,WcFIP1 基 因 與 Phaeosphaeria nodorum 的 蛋 白 質 SNODPROT1 胺 基 酸 相 似 度 達 55% 。 SNODPROT1 是感染小麥葉片時分泌的蛋白質, 與人類病原真菌 Coccidioides immilis 的抗原蛋白 以及 Aspergillus fumigatus 的過敏原蛋白相似 (Hall, et al., 1999)。也許 WcFIP1 會在茯苓感染 針葉樹根時也會被分泌出來,但亦尚待進一步研 究證實。
誌 謝
本報告之完成要感謝行政院科學委員會提供 經費(NSC92-3114-B-003-002)。感謝台大園藝系許 輔教授慨然提供本研究所使用所需之茯苓蛋白、 樟芝蛋白與茯苓蛋白抗體。Wolfiporia EST Library
參考文獻
Adachi K, Nanba H and Kuroda H. 1987. Potentiation of host-mediated antitumor activity in mice by beta-glucan obtained from
Grifola frondosa (maitake). Chem. Pharm.
Bull. (Tokyo) 35:262-270.
Boguski MS, Todd MJ, Lowe TMJ and Tolstoshev CM. 1993. The dbEST-database for “expressed sequence tags”. Nature Genetics 4:332-333.
Collins RA and Ng TB. 1997. Polysaccharopeptide from Coriolus versicolor has potential for use against human immunodeficiency virus type 1 infection. Life Sci. 60:PL383-7.
Hall N, Keon JPR and Hargreaves JA. 1999. A homologue of a gene implicated in the virulence of human fungal diseases is present in a plant fungal pathogen and is expressed during infection. Physiol. Mol. Plant Pathol. 55:69-73.
Hibino Y, Konishi Y, Koike J, Tabata T, Ohashi Y and Sugano N. 1994. Productions of interferon-gamma and nitrite are induced in mouse splenic cells by a heteroglycan-protein fraction from culture medium of Lentinus
edodes mycelia. Immunopharm. 28:77-85.
Hseu YC, Chang WC, Hseu YT, Lee CY, Yech YJ, Chen PC, Chen JY and Yang HL. 2002. Protection of oxidative damage by aqueous extract from Antrodia camphorata mycelia in normal human erythrocytes. Life Sci. 71:469-82.
Hsu HC, Hsu CI, Lin RH, Kao CL and Lin LY. 1997. Fip-vvo, a new fungal immunomodulatory protein isolated from
Volvariella volvacea. Biochem. J. 323:557-
565.
Inoue A, Kodama N, and Nanba H. 2002. Effect of maitake (Grifola frondosa) D-fraction on the control of the T lymph node Th-1/Th-2 proportion. Biol Pharm Bull. 25:536-40.
Jin Y, Zhang L, Zhang M, Chen L, Cheung PC, Oi VE and Lin Y. 2003. Antitumor activities of heteropolysaccharides of Poria cocos mycelia from different strains and culture media. Carbohydr. Res. 338:1517-21.
Kaminaga T, Yasukawa K, Kanno H, Tai T, Nunoura Y and Takido M. 1996. Inhibitory effects of lanostane-type triterpene acids, the components of Poria cocos, on tumor
promotion by 12-O-tetradecanoylphorbol- 13-acetate in two-stage carcinogenesis in mouse skin. Oncology 53:382-5.
Kanehisa M, Goto S, Kawashima S, Okuno Y and Hattori M. 2004. The KEGG resources for deciphering the genome. Nucleic Acids Res. 32:D277-D280.
Kino K, Sone T, Watanabe J, Yamashita A, Tsuboi H, Miyajima H and Tsunoo H. 1991. Immunomodulator, LZ-8, prevents antibody production in mice. Int. J. Immunopharmacol. 13:1109-1115.
Ko JL, Hsu CI, Lin RH, Kao CK and Lin LY. 1995. A new fungal immunomodulatory protein, FIP-fve isolated from the edible mushroom,
Flammulina velutipes and its complete amino
acid sequence. Eur. J. Biochem. 228:244-249. Kobayashi Y, Kariya K, Saigenji K and Nakamura
K. 1994. Suppression of cancer cell growth in vitro by the protein-bound polysaccharide of
Coriolus versicolor QUEL (PS-K) with SOD
mimicking activity. Cancer Biother. 9:63-9. Lee SS, Wei YH, Chen CF, Wang SY and Chen KY.
1995. Antitumor effects of Ganoderma
lucidum. J. Chinese Med. 6:1-12.
Lin WH, Hung CH, Hsu CI and Lin JY. 1997. Dimerization of the N-terminal amphipathic alpha-helix domain of the fungal immunomodulatory protein from Ganoderma
tsugae (Fip-gts) defined by a yeast two-hybrid
system and site-directed mutagenesis. J. Biol. Chem. 272:20044-20048.
Liu WK, Ng TB, Sze SF and Tsui KW. 1993. Activation of peritoneal macrophages by polysaccharopeptide from the mushroom,
Coriolus versicolor. Immunopharmacol.
26:139-46.
Murasugi A, Tanaka S, Komiyama N, Iwata N, Kino K, Tsunoo H and Sakuma S. 1991. Molecular cloning of a cDNA and a gene encoding an immunomodulatory protein, Ling Zhi-8, from a fungus, Ganoderma lucidum. J. Biol. Chem. 266:2486-2493.
Ng TB. 1998. A review of research on the protein-bound polysaccharide (polysaccharo- peptide, PSP) from the mushroom Coriolus
versicolor (Basidiomycetes: Polyporaceae).
Gen Pharmacol. 30:1-4.
Ohsawa T, Yukawa M, Takao C, Murayama M and Bando H. 1992. Studies on constituents of fruit body of Polyporus umbellatus and their
cytotoxic activity. Chem. Pharm. Bull. (Tokyo) 40:143-7.
Takeyama T, Suzuki I, Ohno N, Oikawa S, Sato K, Ohsawa M and Yadomae T. 1987. Host-mediated antitumor effect of grifolan NMF-5N, a polysaccharide obtained from
Grifola frondosa. J. Pharmacobiodyn.
10:644-51.
Tanaka S, Ko K, Kino K, Tsuchiya K, Yamashita A, Murasugi A, Sakuma S and Tsunoo H. 1989. Complete amino acid sequence of an immunomodulatory protein, ling zhi-8 (LZ-8). An immunomodulator from a fungus,
Ganoderma lucidium, having similarity to
immunoglobulin variable regions. J. Biol. Chem. 264:16372-16377.
Wasser SP. and Weis AL. 1999. Therapeutic effects of substances occurring in higher
Basidiomycetes mushroom: a modern
perspective. Crit. Rev. Immuno. 19:65-96.
Yan SC. 1988. Linical and experimental research on
Polyporus umbellatus polysaccharide in the
treatment of chronic viral hepatitis. Zhong Xi Yi Jie He Za Zhi. 8:141-3.
Zhang YH, Liu YL and Yan SC. 1991. Effect of
Polyporus umbellatus polysaccharide on
function of macrophages in the peritoneal cavities of mice with liver lesions. Zhong Xi Yi Jie He Za Zhi. 11:225-6. 財團法人食品工業發展研究所. 2000. 真菌目錄. 財團法人食品工業發展研究所 pp. 161-182. 閔 三 弟, 1996. 真菌 的藥 用 價值 . 食 用 菌學報 3:55-64. 張慧欣. 2005. 茯苓免疫調節蛋白的純化、機能活 性及生理角色之探討. 國立台灣大學, 園藝 學研究所碩士論文. 錢家樂. 2004. 樟芝免疫調節蛋白基因選殖及表 現之研究. 國立台灣大學, 園藝學研究所碩 士論文.
BioFormosa(2007)42(1): 47-53
Construction and Analysis of EST Library in Wolfiporia cocos
Tsung-Hsun Wu, Chih-Wen Sun*
Department of Life Sciences, National Taiwan Normal University Taipei, Taiwan
(Received: 13 October 2006, accepted: 26 October 2006)
ABSTRACT
Wolfiporia cocos is the most popular fungal ingredient in Chinese prescription because it functions in arresting the growth of tumer cells or stimulating immune systems. Nevertheless, knowledge regarding to its bioactive compounds and genomic information are limited. Thus, we would like to establish an expressed sequence tag (EST) library from Wolfiporia cocos and explore candidates with pharmaceutical potentials in this study. Characterization of Wolfiporia EST library reveal that 16%, 21%, 7% and 3% of genes are participating in metabolism, genetic information process, environmental information process, cellular process, respectively. 53% of those are unknown genes. Furthermore, a gene encodes a protein sharing 50% sequence similarity to Taiwanofungus camphorate fungal immunomodulatory protein (Aca1), and is therefore named as WcFIP1. We hypothesize that the newly isolated WcFIP1 might be one of the fungal immunomodulatory proteins in Wolfiporia cocos.
Key words: Wolfiporia cocos, expressed sequence tag (EST) library, fungal immunomodulatory protein