expression at the RNA level. In this article we further express and characterize the induction of a novel ribonucleotide reductase after WSSV infection of shrimp cells. A baculovirus/insect system was used to express the two recombinant protein subunits RR1 and RR2, and a DNA polymerase coupled RR activity assay showed a marked increase in ribonucleotide reductase activity when cell extracts containing recombinant RR1 and RR2 were combined. The same assay revealed that RR activity increased as infection advanced in the gills of experimentally infected shrimp. An increase in RR expression was also detected at the protein level in WSSV-infected shrimp cells. An immunocytochemistry assay by confocal laser scanning microscopy showed that in hemocytes collected from WSSV-infected shrimp, both of the subunit proteins (RR1 and RR2) were concentrated mainly around the nucleus, but only RR1 was detected inside it. All of these results suggest that WSSV RR is functionally involved during WSSV infection. © 2002 Elsevier Science (USA)
Key Words: Penaeus monodon; white spot syndrome virus; WSSV; ribonucleotide reductase activity.
INTRODUCTION
The complete sequence of the white spot syndrome virus (WSSV) genome has now been published for three different isolates (GenBank Accession Nos. AF332093, AF369029, AF440570 for viruses isolated from China, Thailand, and Taiwan, respectively). For two of these isolates (China and Thailand), an analysis of the com-plete WSSV genome has been published (Yang et al., 2001; van Hulten et al., 2001; Chen et al., 2002). However, most (⬃70%) of the 181 annotated WSSV ORFs encode proteins that have no homology to any known proteins or motifs (Yang et al., 2001), and the uniqueness of this virus makes it is difficult to directly apply other virus infection models to interpret the infection strategy of WSSV. This means that WSSV’s infection strategy must be investigated ab initio. Nevertheless, the WSSV ge-nome is already known to contain some genes/ORFs that encode homologues of cellular enzymes involved in nu-cleic acid metabolism. To date, the known enzymes are ribonucleotide reductase (RR) (Tsai et al., 2000a), dUT-Pase, thymidylate synthase, and chimeric polypeptide of thymidine kinase and thymidylate kinase (TK-TMK) (Tsai
et al., 2000b; Tseng et al., 2002). The presence of each of
these enzymes gives the virus specific advantages. For example, viruses with their own RRs are able to replicate in nondividing cells in which the cellular RRs are at very low levels (Yamada et al., 1991). However, in the case of WSSV, it must first be established that WSSV RR is in fact functionally active in infected shrimp cells.
In the present article we therefore focus on characteriz-ing WSSV’s RR activity. To this end, the genes of WSSV rr1 and rr2 were expressed by Autographa californica multiple
nucleopolyhedrovirus (AcMNPV)-based recombinant virus
for the production of recombinant WSSV RR1 and RR2 in insect cells, and we show here that the recombinant RR1 and RR2 associate to form a functional holoenzyme that exhibits catalytic activity. An RR activity assay and Western blotting analysis with antibodies against WSSV RR2 further show that in lysate from the stomach cells of WSSV-infected shrimp, the WSSV RR2 protein level increases as the infec-tion phase advances. We also demonstrate that RR activity increases as infection advances in the gills of experimen-tally infected shrimp. These results suggest that WSSV RR is functionally involved during WSSV infection.
RESULTS AND DISCUSSION
Overexpression of recombinant WSSV RR1 and RR2 in insect cells
To construct the two AcMNPV-based recombinant vi-ruses for overexpressing WSSV RR1 and RR2,
respec-1To whom correspondence and reprint requests should be addressed
at Department of Zoology, National Taiwan University, Taipei 106, Taiwan R.O.C. Fax: 886-2-23638179. E-mail: [email protected] or [email protected].
0042-6822/02 $35.00
© 2002 Elsevier Science (USA) All rights reserved.
tively, in insect cells, the full-length WSSV rr1 and rr2 coding regions were each cloned to the polyhedrin lo-cus-based single promoter transfer vector pAcSGHis NT-A (PharMingen, San Diego, CA) in-frame with the hexa-histidine-tag (6His-tag). The BacVector Transfection system (Novagen, Madison, WI) was used to produce the recom-binant viruses. The recomrecom-binant viruses containing rr1 and
rr2 were designated AcMNPV-rr1 and AcMNPV-rr2,
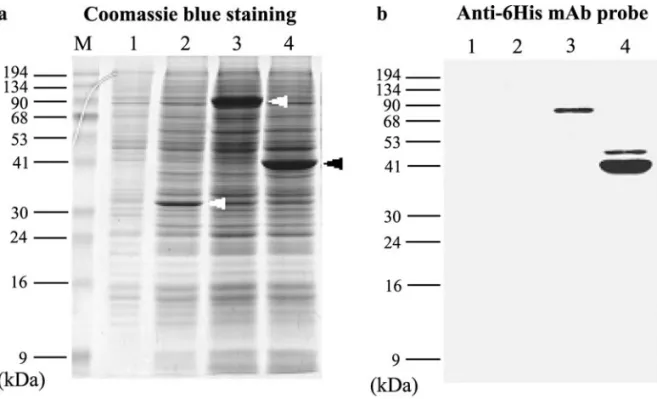
respec-tively. The recombinant viruses encoded 27 and 30 addi-tional amino acid residues for RR1 and RR2, including the 6His tag appended to the N-terminus of WSSV RR1 and RR2. As shown in Fig. 1, the apparent molecular mass of the recombinant proteins on sodium dodecyl sulfate–poly-acrylamide gel electrophoresis (SDS–PAGE) was 81.7 kDa for the RR1 fusion protein (rRR1) and around 43.4 kDa for the RR2 fusion protein rRR2 (the predicted molecular mass of RR1 and RR2 are 95.6 and 47.6 kDa). The intensity of the bands suggests that the yield of both rRR1 and rRR2 in insect cells was quite high (Fig. 1a).
Expression of rRR1 and rRR2 was confirmed by Western blot analysis using a 6His-tag monoclonal antibody (Fig. 1b). The fusion proteins were then purified using nickel beads and used to generate WSSV RR1 and RR2 anti-sera. In control immunoblot assays (data not shown) these antisera reacted only with the target proteins and did not show any cross-reactions with the other recombinant pro-teins or with any insect cell or baculovirus propro-teins. These polyclonal antisera were then used to monitor rRR1 and rRR2 expression levels in the insect cell expression system over time (Fig. 2). rRR1 was first detected at 36 h
postinfec-tion (h.p.i.); it increased over the next 36 h and then de-creased. The rRR1 was fairly stable, and only one band of protein was observed on the gel. rRR2 was also first de-tected at 36 h.p.i. and expression levels continued to in-crease to the end of the observation period (84 h.p.i.). Starting at 48 h.p.i., a second smaller band of protein ap-peared on the gel, with levels of this smaller form subse-quently increasing over time.
FIG. 1. Baculovirus expression of WSSV RR1 and RR2 in insect cells infected with corresponding recombinant viruses for 3 days. (a) Coomassie brilliant blue stained 12.5% SDS–PAGE gel with total lysates of Sf9 cells. Lane1: mock infection; Lane 2: infected with wild-type (wt) AcMNPV; Lane 3: infected with AcMNPV-rr1; Lane 4: infected with AcMNPV-rr2. Arrowheads show polyhedrin, RR1, and RR2 in Lanes 2, 3, and 4, respectively. (b) Western blot of a duplicate gel probed with anti-6His monoclonal antibody and developed by an ECL chemiluminescence system using goat anti-mouse IgGcoupled to horseradish peroxidase as secondary antibody.
FIG. 2. Temporal expression analysis of WSSV rRR1 and rRR2 in insect cells infected with AcMNPV-rr1 and AcMNPV-rr2, respectively, by Western blot assay using antibodies specific to (a) WSSV RR1 and (b) RR2 and an ECL chemiluminescence system with goat anti-rabbit IgGcoupled to horseradish peroxidase as secondary antibody. Lane headings show hours postinfection (h.p.i.).
WSSV rRR1/rRR2 interaction in whole-cell lysates as determined by co-immunoprecipitation
To discover if the interaction between WSSV rRR1 and rRR2 occurs in whole-cell lysates, crude extracts from Sf9 cells overexpressing WSSV rRR1 and rRR2, respec-tively, were combined. This combined mixture was first immunoprecipitated with WSSV rRR2 antibody and then probed with WSSV rRR1 antibody by Western immuno-blotting. Figure 3 shows that rRR1/rRR2 was co-immuno-precipitated by WSSV rRR2 antibody in whole-cell lysate, which suggests that the interaction between WSSV rRR1 and rRR2 occurred as predicted.
A reciprocal experiment was also performed in which the combined lysates were first immunoprecipitated with WSSV rRR1 antibody and then probed with WSSV rRR2 antibody. However, since the protein A band occurs at the same position on the gel as the rRR2 band, no clear results could be obtained (data not shown).
WSSV rRR activity in crude extracts of Sf9 cells infected with AcMNPV-rr recombinant viruses
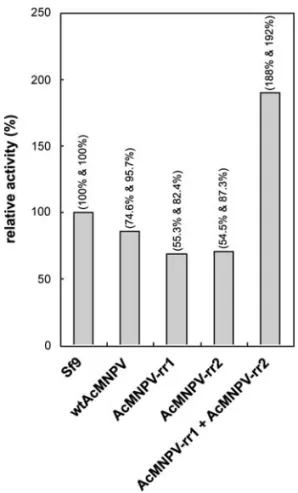
The formation of functionally active RR in infected cell lysates was further verified by the DNA polymerase-coupled RR activity assay, a simple and sensitive assay developed by Jong et al. (1998). This assay was used to measure RR enzymatic activity in crude desalted extracts from AcMNPV-rr1- and AcMNPV-rr2-infected Sf9 cells using a standard reaction mixture. Figure 4 shows that the RR activity levels, indicated by dCDP formation
monodon (Fig. 5b, Lane 1). Native WSSV RR2 was,
how-ever, detected in the same specimen (Fig. 5c, Lane 1). As expected, the native WSSV RR2 is about 5 kDa smaller than the fusion protein of the recombinant WSSV RR2 from the insect cell system (43.4 kDa vs 48.4 kDa). In a temporal analysis (Fig. 6), the native WSSV RR2 was de-tected at 18 and 60 h.p.i. The smaller form of WSSV RR2 was also observed at the late infection phase (60 h.p.i.).
FIG. 4. RR activity in desalted cell extracts (⬃60g) prepared from mock-infected Sf9 cells and Sf9 cells infected with the following: wild-type (wt) AcMNPV; AcMNPV-rr1; AcMNPV-rr2; and both AcMNPV-rr1 and AcMNPV-rr2 in combination. Data show the relative RR activity (i.e., the percentage RR activity with respect to the in Sf9 control cells) and are the combined (mean) results of two independent assays. Percent-ages in parentheses above each bar are the relative activities in the first and second independent assays, respectively.
FIG. 3. Co-immunoprecipitation of WSSV rRR1/rRR2 in cell lysates. Lysates were incubated with WSSV RR2-specific antibody in the pres-ence of protein A-coated Sepharose. The resulting immune complex was analyzed by SDS–PAGE in 12.5% acrylamide gel and then probed with WSSV rRR1 antibody, developed by an ECL chemiluminescence system using goat anti-rabbit IgGcoupled to horseradish peroxidase as secondary antibody. Lysates were from the following: Lane 1: Ac-MNPV-rr1-infected Sf9 cells; Lane 2: AcMNPV-rr2-infected Sf9 cells; Lane 3: mixed lysates from AcMNPV-rr1- and AcMNPV-rr2-infected Sf9 cells.
Expression and cellular localization analysis of WSSV RRs by light and confocal laser scanning microscopy (cLSM)
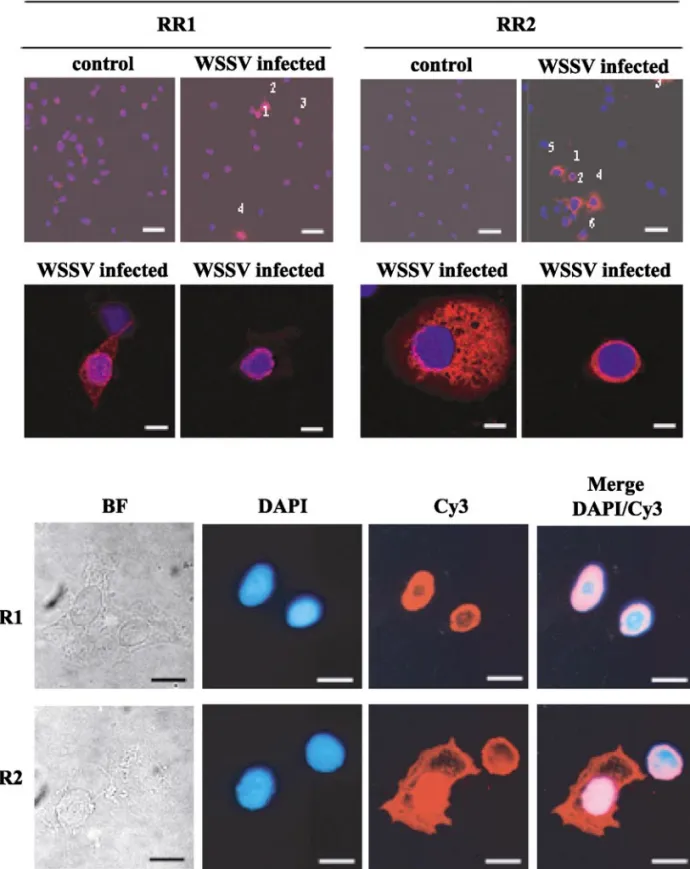
Shrimp hemocytes are WSSV target cells (Lo et al., 1997). In this study, we used hemocytes collected from experimentally infected shrimp to study the expression of WSSV RR1 and RR2 at the cellular level. The hemocytes were attached to cover slips, and the cells were labeled with antibodies specific against WSSV RR1 and RR2, respectively, for the immunocytochemistry assay. Obser-vation by cLSM showed no cross-reactions in the hemo-cytes from control (uninfected) shrimp, while both the anti-RR1 and the anti-RR2 antisera produced fluores-cence in hemocytes from WSSV-infected shrimp (Fig. 7, row 1). However, the fluorescence was much weaker in cells immunostained with anti-WSSV RR1 antisera and there were also fewer positive cells. This result is
con-sistent with our earlier failure to detect WSSV RR1 by Western blotting (see Fig. 5). It is possible that these observations were the results of differential expression of the two RR subunits, as has been reported in actively dividing mammalian cells (Engstrom and Rozell, 1988). However, other explanations, such as unequal antibody sensitivity, are also possible. Clearly this will require further investigation.
The cell localizations of RR subunits vary from organ-ism to organorgan-ism and may be exclusively cytoplasmic (e.g., mammalian RR small and large subunits) or mainly nuclear (e.g., Leishmania mexicana amazonensis RR small subunit) (Lye et al., 1999). In the case of WSSV, “optical sections” made by cLSM suggest that both pro-teins are concentrated mainly around the nucleus of the shrimp hemocytes and that RR1 also occurs inside it (Fig. 7, row 2). Fluorescent microscopy further confirmed that both proteins were present in the cytoplasm of the hemocytes (Fig. 7, last 2 rows). Strong RR2-positive sig-nals in the cytoplasm of some cells (Fig. 7, row 2, column 3; row 4) suggest that WSSV RR2 was diffused through-out the cytoplasm. WSSV RR1 may also be similarly diffused, but the RR1-positive signals in the cytoplasm are generally much weaker (Fig. 7, row 2, column 1). The molecular targeting mechanism of RR1 and RR2 still remains unclear and will require further investigation. RR activity in WSSV-infected shrimp organs
From six P. monodon shrimps (two uninfected speci-mens, two at 18 h.p.i., and two at 60 h.p.i.), the gills and hearts were separately removed, frozen, and stored in liquid nitrogen until used in this DNA polymerase-cou-pled RR activity assay. As shown in Fig. 8, by 60 h.p.i. RR activity increased markedly in the gill. By contrast, in the
FIG. 5. Detection of WSSV RR1 and RR2 in the stomach of infected shrimp. (a) Coomassie brilliant blue stained 12.5% SDS–PAGE gel. Lysates are from WSSV-infected shrimp cells at 60 h.p.i. (Lane 1) and from Sf9 cells infected for 72 h with AcMNPV-rr1 (Lane 2) and AcMNPV-rr2 (Lane 3). Western blots are of duplicate SDS–PAGE gels probed with (b) anti-rRR1 and (c) anti-rRR2 polyclonal antibodies and developed by an ECL chemiluminescence system with goat anti-rabbit IgGcoupled to horseradish peroxidase as secondary antibody.
FIG. 6. Temporal expression analysis of WSSV rr2 at the protein level in experimentally infected shrimp stomach by Western blot assay using WSSV RR2-specific antibodies and an ECL chemiluminescence system with goat anti-rabbit IgGcoupled to horseradish peroxidase as sec-ondary antibody. Recombinant WSSV RR2 from Sf9 cells infected with AcMNPV-rr2 at 72 h.p.i. is shown for reference.
heart, increases in RR activity over time were only very slight. This result is consistent with a previous study of experimentally infected P. monodon shrimp (Tzeng et al., 2002), in which a thymidine kinase activity assay
re-vealed that increases in TK activity in the heart were only very slight over time. In contrast to the gills, which are a major target organ of WSSV, the heart is not a main target and it therefore serves as a good “internal” control.
FIG. 7. Confocal laser scanning micrographs (rows 1 and 2) and light micrographs (rows 3 and 4) of hemocytes collected from uninfected (control) and WSSV-infected shrimp reacted first with antibodies specific to either WSSV RR1 or RR2, and then with Cy3-conjugated goat anti-rabbit IgG antibody. Rows 1 and 2 were counterstained with DAPI, and each photograph is a merged composite of the DAPI and Cy3 fluorescent signals. Row 1: Positive cells are indicated by Arabic numerals. Bar: 40m. Row 2: Higher magnification of selected RR1- and RR2-positive cells. Bar: 8 m. Rows 3 and 4: Light micrographs of two WSSV RR1 (row 3) and two RR2 (row 4) positive hemocytes from WSSV-infected shrimp. Column 1: bright field microscope; Column 2: nuclear (DAPI) fluorescent signals; Column 3: RR antigen-positive fluorescent signals; Column 4: merged DAPI and Cy3 images. Bar: 8m.
This assay was repeated using the eight other shrimps from the same batch of experimentally infected shrimp at 0 (uninfected), 18, 36, and 60 h.p.i. (two shrimps for each time point; three separate cell extract from each sample of gill tissue). Results were similar to the previ-ous assay, with no increase in the RR activity at 18 h.p.i., and higher RR activity levels at 36 and 60 h.p.i. (Fig. 9).
RR is a key enzyme for DNA metabolism and DNA synthesis (Jordan and Reichard, 1998). RR is also tightly regulated in the cell cycle, closely coupled to cell DNA replication, and is absent or inactive in resting cells (Nordenskjo¨ld et al., 1970). Some large DNA viruses such as herpesviruses and poxviruses synthesize their own ribonucleotide reductases (Cohen, 1972; Lankinen et al., 1982; Slabaugh and Mathews, 1984), and although RR-deficient HSV-1 mutants can grow well in the still-devel-oping brains of newborn mice, they cannot grow in the brains of older mice (see, e.g., Yamada et al., 1991). Thus the viral RRs are thought to benefit the virus by enabling them to replicate in nondividing cells. The rapid onset and lethality of the disease caused by WSSV are well known (see, e.g., Chang et al., 1996). WSSV attacks almost all cells originating from the ectoderm and endoderm within a short period. In the present study, in which we have shown that functionally active WSSV RR exists in WSSV-infected shrimp, the presence of this functionally active enzyme may explain why WSSV rep-licates efficiently not only in young/actively proliferating cells but also in old/resting cells.
MATERIALS AND METHODS Plasmid constructions
The coding regions of WSSV rr1 and rr2 were am-plified by polymerase chain reaction (PCR) from total
WSSV DNA. The forward and reverse primers used for rr1 amplification were 5
⬘-TAACATCTCGAGGGTTCTA-ACCAGCAACAA-3⬘ and
5⬘-TGTCTTCCCGGGCTAGGA-AGAACACATTTC-3⬘. The primers used for rr2 were
5⬘-CCGAATTCATGGAGTCAATCAAAC-3⬘ and
5⬘-GACA-CAACCCGGGAACTA AAAATCGTCATAACT-3⬘. The
bases indicate the restriction sites. The 5⬘ primer for
rr1 was designed to insert at a XhoI site, and the 3⬘
primer was designed to insert at a SmaI site. The primers for rr2 were designed to insert at EcoRI and
SmaI sites. The reaction mixtures were subjected to
PCR and PCR products were purified using a high pure PCR product purification kit (Roche, Mannheim, Ger-many). The plasmid pAcSGHis NT-A (PharMingen) and purified PCR products were digested with restric-tion endonucleases. The oligonucleotides were sepa-rated by 1.5% agarose gel electrophoresis, and the rr1-and rr2-containing fragments rr1-and digested pAcSGHis NT-A were extracted with a QIAquick gel extraction kit (Qiagen, Germany). The rr1 and rr2 inserts were li-gated with digested pAcSGHis NT-A and inserted into the multiple cloning site downstream of the polyhedrin promoter. The resulting plasmids were named pAcSG His NTA-rr1 and pAcSGHis NTA-rr2.
Plasmids were purified by QIAprep spin miniprep kit (Qiagen) and then quantified by spectrophotometer at 260 nm.
Cells and virus
Insect cells (Sf9) were cultured in Sf900-II insect se-rum-free medium (Gibco-BRL, Life Technologies Inc., Rockville, MD). A. californica multicapsid
nucleopolyhe-FIG. 8. WSSV RR activity in desalted cell extracts from the heart and gills of uninfected shrimps (0 h.p.i.) and infected shrimp at 18 and 60 h.p.i. RR activity is indicated by the formation of dCDP (pmol/30 min/0.15 mg). Each time point shows data from two different shrimps.
FIG. 9. WSSV RR activity in desalted cell extracts from gills of uninfected shrimps (0 h.p.i.) and shrimps at 18, 36, and 60 h.p.i. RR activity is indicated by the formation of dCDP (pmol/30 min/0.10 mg). Each time point shows pooled data from three separate extracts taken from two different shrimp. Error bars show standard deviations.
and then the cells were harvested by centrifugation at 1000 g for 15 min at 4°C. The supernatants were dis-carded, and the cell pellets were stored at⫺20°C until used.
The cell pellets from one 25-cm2
T-flask were resus-pended in 0.4 ml lysis buffer (20 mM Tris–HCl, pH 8.0; 300 mM NaCl; 5 mM imidazole) and subjected to several bursts of sonication at 4°C. The lysate was centrifuged at 15,000 g for 20 min at 4°C, and the supernatant was mixed with 0.2 ml Ni-NTA resin (Qiagen) that had been equilibrated with binding buffer. The mixture was mixed gently for 30 min at 4°C, centrifuged for 10 s at 15,000 g to pellet the resin, and washed with 1 ml washing buffer (20 mM Tris–HCl, pH 8.0; 300 mM NaCl; 20 mM imida-zole) twice. The fusion protein was eluted with 0.3 ml elution buffer (20 mM Tris–HCl, pH 8.0; 300 mM NaCl; 250 mM imidazole). The concentration of protein was determined by Bio-Rad protein assay solution (Bio-Rad, Hercules, CA) with bovine serum albumin as standard. Preparation of antibodies
Polyclonal hyperimmune serum against RR1 or RR2 was prepared in New Zealand white rabbits. RR1 or RR2 (350g of each) was mixed well with Freund’s complete adjuvant and then injected subcutaneously at day 0. Subsequent booster injections of 250g of RR1 or RR2 in Freund’s incomplete adjuvant were given on days 13 and 27. The rabbits were exsanguinated on day 34. The whole blood was centrifuged at 1000 g for 30 min at 4°C. The cell pellets were discarded, and the sera were stored at ⫺20°C until used.
Co-immunoprecipitation of WSSV rRR1/RR2 in whole-cell lysates
The ability of WSSV rRR1 and rRR2 in whole-cell ly-sates to associate together to form a holoenzyme was analyzed by immunoprecipitation. Sf9 cells infected with AcMNPV-rr1 and AcMNPV-rr2, respectively, for 3 days were used to prepare whole-cell lysates using the fol-lowing procedure. Infected cells from 75-cm2
culture
The pellets were washed twice with 1 ml lysis buffer and once with ice-cold RIPA (150 mM NaCl; 1% NP-40; 0.1% SDS; 50 mM Tris, pH 8.0; 1 mM PMSF) and were then resuspended in 50 l SDS sample buffer. The immune complex was boiled for 5 min and centrifuged at 12,000
g for 30 s, and 10 l supernatant was removed and
analyzed by SDS–PAGE in 12.5% acrylamide gel. WSSV rRR1 in the immune precipitate was detected by Western blot using WSSV rRR1 antibody and the enhanced chemi-luminescent-light (ECL) detection kit (NEN Life Sciences, Boston, MA) as described below. AcMNPV-rr1-infected cell lysate and AcMNPV-rr2 infected cell lysate were used as sample controls. Mixed cell lysate incubated only with protein A–Sepharose was included as reagent control.
SDS–PAGE and immunoblotting
Insect cells infected with wild-type AcMNPV, and re-combinant AcMNPV viruses expressing heterologous proteins (RR1, and RR2), were analyzed in 12.5% SDS– PAGE according to the method of Laemmli (1970) as follows. After removal of the medium, the cells were resuspended with the lysis buffer [25 mM HEPES, pH 7.4; 1% Triton X-100; 0.1% SDS; 150 mM KCl; 1 mM EDTA; 1 mM DTT, and protease inhibitor cocktail (Roche)] and then mixed with the same volume of 2⫻ SDS sample buffer. After separation on SDS–PAGE, the proteins were transferred to nitrocellulose (NC) paper using a blotting buffer (48 mM Tris base; 39 mM glycine; 0.037% SDS; 20% methanol) at 2 mA/cm2 for 30 min by Trans-Blot SD cell (Bio-Rad).
After transfer, the NC paper was blocked for 1 h with 3% skimmed milk in TBS (200 mM NaCl; 50 mM Tris–HCl, pH 7.5). The NC paper was washed twice in TBS for 5 min and incubated with primary monoclonal anti-6 His antibody (Clontech, Palo Alto, CA) at a dilution of 1/3000, polyclonal rabbit anti-RR1 antibody at a dilution of 1/10,000, and anti-RR2 antibody at a dilution of 1/20,000 in blocking buffer for 1 h at room temperature. The membrane was then washed three times in TBS
contain-ing 0.5% Tween 20 (TBS/T) for 5 min each. Subsequently, anti-rabbit IgGantibody conjugated with horseradish peroxidase (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) was used at a concentration of 1/2500 in TBS. The membrane was washed three times with TBST and TBS for 5 min each. The detection was performed with the ECL detection kit (NEN Life Sciences).
Light and laser scanning microscopy for cell
localization assay in hemocytes from WSSV-infected shrimp
Healthy (that is, two-step WSSV diagnostic PCR neg-ative; Lo et al., 1996) shrimp (P. monodon) and shrimp 72 h after infection with WSSV by intramuscular injection using the method described previously by Tsai et al. (1999) were used for this study. Hemolymphs were col-lected from the ventral part of the hemocoel of the sec-ond abdominal segment using a 25-gauge needle and a 1-ml syringe containing 0.5 ml cold modified Alsever’s solution (AS; 19.3 mM sodium citrate; 239.8 mM NaCl; 182.5 mM glucose; 6.2 mM EDTA, pH 7.0). Hemocyte monolayers were prepared by seeding hemocytes in 100 l hemolymph and allowing them to settle for 1 h onto a cover glass placed in each well of a 24-well microplate (Nunc Brand Products, Nalge Nunc International, Rosk-ide, Denmark). The monolayers were then washed twice with PBS. The hemocytes were fixed in paraformalde-hyde (4% in PBS) for 10 min at 4°C, treated with acetone (3 min at 4°C), and then washed thoroughly two times with PBS. After blocking in bovine serum albumin and normal goat serum (both diluted to 5% in PBS) for 16 h at 4°C, the hemocyte monolayers were treated with 1000⫻ PBS-diluted antisera (250 l) specific to WSSV RR1 or RR2 (4 h at room temperature). The cells were then washed with PBST (0.2% Tween 20 in PBS) and reacted with 1000⫻ PBST-diluted carboxymethylindocyanine (Cy3) dye-conjugated goat anti-rabbit IgGantibody (Sig-ma, St. Louis, MO) for 4 h at room temperature. Coun-terstaining of the nucleus was performed with 4 ⬘-6⬘-diamidino-2-phenylindole dihydrochloride (DAPI; Vector Laboratories, Burlingame, CA). After washing twice with PBST (15 min each time), the cover glasses were wet mounted with 60% glycerol. During all of the above re-actions, the monolayers were kept in darkness. Fluores-cent signals were observed with an Olympus Research Microscope (Model AHBT3) with a UV microscopy appa-ratus. The same specimens were also examined using a confocal laser-scanning microscope (Leica TCS SP2). Preparation of total soluble proteins from organs of WSSV-infected P. monodon shrimps for enzyme assay
Healthy uninfected P. monodon shrimp and shrimp at 18 and 60 h after infection with WSSV by intramuscular injection were used for the in vivo WSSV RR activity assay. The gills and heart (⬃0.25 g/each) were removed
from two uninfected and four WSSV-infected shrimp (two specimens for each time point) and washed with buffer I [500 mM HEPES, pH 7.2; 2 mM dithiothreitol (DTT)]. Each washed organ was then placed in a microcentrifuge tube and an aliquot (0.5 ml) of new buffer I was added. RNase A was added and the cells were homogenized. After homogenization with a wooden splint, the mixture was sonicated (three 10-s bursts) and centrifuged at 20,000 g for 20 min. The supernatant was loaded onto a Sephadex G-25 column (⬃4 ml) equilibrated with buffer II (50 mM HEPES, pH 7.2; 2 mM DTT), and then 2 ml of buffer I was added to elute the column. Fractions of 0.5 ml were collected from each tube. The concentration of total sol-uble proteins in each fraction was determined using Bio-Rad protein assay solution. The fraction containing 96 g of total soluble proteins was used for the RR activity assay.
Enzyme activity assay
The RR enzyme activity assay used here was based on the DNA polymerase coupling assay described by Jong
et al. (1998) with slight modifications. In this assay, RR
reduces tritium-labeled [3H]CDP to [3H]dCDP, a reaction which is coupled to subsequent phosphorylation to [3H]dCTP [catalyzed here by the endogenous nucleotide diphosphate (NDP) kinase]. [3
H]dCTP is then incorpo-rated into newly synthesized DNA by the DNA polymer-ase. The resultant [3H]-labeled DNA, which can be easily separated from the ribonucleotides, is then subjected to scintillation counting (Jong et al., 1998). The infected insect cells were collected, washed twice with buffer I, and resuspended in 0.5 ml buffer I. RNase A (final con-centration: 0.2 mg/ml) was added and the cells were homogenized. All steps were carried out on ice. The homogenate was centrifuged at 12,000 g at 4°C for 20 min, and the endogenous nucleotides were removed with a G-25 column. Aliquots of between 25 and 100g of cell proteins were used in each RR assay.
The RR assay solution contained 50 mM HEPES, pH 7.2, 6 mM DTT, 4 mM Mg acetate, 2 mM ATP, and 50 pmol [3H]CDP. The mixture was incubated at 37°C for 30 min and boiled for 5 min. Then the mixture was centrifuged for 5 min to remove the precipitates. The labeling mixture contained 90 mM HEPES (pH 6.6), 10 mM MgCl2, 0.2 mM dNTP, 1 g/l herring DNA, 150 ng/l 6-mer random primer (Roche) and 5 units Klenow fragment. The RR reaction mixture was added and after incubation at room temperature for 30 min, the mixture was spotted onto a piece of 1.5-cm2 Whatman DE-81 filter paper. After air-drying, the filter was washed for 5 min with 5% Na2HPO4 twice, once with methanol (5 ml/filter), and once with acetone (5 ml/filter). The washed filters were air-dried and inserted into vials containing 3 ml of the scintillation solution FluoranSafe 2 Scintran (BDH, Poole, U.K.).
protein (VP35) gene of shrimp white spot syndrome virus and char-acterization of the motif important for targeting vp35 to nuclei of transfected insect cells. Virology 293, 44–53, doi:10.1006/viro. 2001.1273.
Cohen, G. H. (1972). Ribonucleotide reductase activity of synchronized KB cells infected with herpes simplex virus. J. Virol. 9, 408–418. Engstrom, Y., and Rozell, B. (1988). Immunocytochemical evidence for
the cytoplasmic localization and differential expression during the cell cycle of the M1 and M2 subunits of mammalian ribonucleotide reductase. EMBO J. 7, 1615–1620.
Jong, A. Y., Yu, K., Zhou, B., Frgala, T., Reynolds, C. P., and Yen, Y. J. (1998). A simple and sensitive ribonucleotide reductase assay.
Biomed. Sci. 5, 62–68.
Jordan, A., and Reichard, P. (1998). Ribonucleotide reductases. Annu.
Rev. Biochem. 67, 71–98.
Laemmli, U. K. (1970). Cleavage of structural proteins during the as-sembly of the head of bacteriophage T4. Nature 227, 680–685. Lankinen, H., Graslund, A., and Thelander, L. (1982). Induction of a new
ribonucleotide reductase after infection of mouse L cells with pseu-dorabies virus. J. Virol. 41, 893–900.
Lo, C. F., Ho, C. H., Chen, C. H., Liu, K. F., Chiu, Y. L., Yeh, P. Y., Peng, S. E., Hsu, H. C., Liu, H. C., Chang, C. F., Su, M. S., Wang, C. H., and Kou, G. H. (1997). Detection and tissue tropism of white spot syn-drome baculovirus (WSBV) in captured brooders of Penaeus
mon-odon with a special emphasis on reproductive organs. Dis. Aquat. Org. 30, 53–72.
Lo, C. F., Leu, J. H., Ho, C. H., Chen, C. H., Peng, S. E., Chen, Y. T., Chou,
viro.2000.0956.
Tsai, M. F., Yu, H. T., Tzeng, H. F., Leu, J. H., Chou, C. M., Huang, C. J., Wang, C. H., Lin, J. Y., Kou, G. H., and Lo, C. F. (2000b). Identification and characterization of a shrimp white spot syndrome virus (WSSV) gene that encodes a novel chimeric polypeptide of cellular-type thymidine kinase and thymidylate kinase. Virology 277, 100–110, dio:10.1006/viro.2000.0957.
Tsai, M. F., Kou, G. H., Liu, H. C., Liu, K. F., Chang, C. F., Peng, S. E., Hsu, H. C., Wang, C. H., and Lo, C. F. (1999). Long-term presence of white spot syndrome virus (WSSV) in a cultured shrimp population without disease outbreaks. Dis. Aquat. Org. 38, 107–114.
Tzeng, H. F., Chang, Z. F., Peng, S. E., Wang, C. H., Lin, J. Y., Kou, G. H., and Lo, C. F. (2002). Chimeric polypeptide of thymidine kinase, and thymidylate kinase of shrimp white spot syndrome virus (WSSV): Thymidine kinase enzyme activity of the recombinant protein ex-pressed in baculovirus/insect cell system. Virology, 299, 248–255. van Hulten, M. C., Witteveldt, J., Peters, S., Kloosterboer, N., Tarchini, R.,
Fiers, M., Sandbrink, H., Lankhorst, R. K., and Vlak, J. M. (2001). The white spot syndrome virus DNA genome sequence. Virology 286, 7–22, doi:10.1006/viro.2001.1002.
Yang, F., He, J., Lin, X., Li, Q., Pan, D., Zhang, X., and Xu, X. (2001). Complete genome sequence of the shrimp white spot bacilliform virus. J. Virol. 75, 11811–11820.
Yamada, Y., Kimura, H., Morishima, T., Daikoku, T., Maeno, K., and Nishiyama, Y. (1991). The pathogenicity of ribonucleotide reductase-null mutants of herpes simplex virus type 1 in mice. J. Infect. Dis. 164, 1091–1097.