Original Report: Laboratory Investigation
Am J Nephrol 2009;29:43–53 DOI: 10.1159/000150600
Pentoxifylline Inhibits Transforming Growth
Factor-Beta Signaling and Renal Fibrosis in
Experimental Crescentic Glomerulonephritis
in Rats
Yee-Yung Ng
a
Yung-Ming Chen
b
Tun-Jun Tsai
b
Xiao-Ru Lan
c
Wu-Chang Yang
a
Hui Y. Lan
c
a Section of Nephrology, Department of Medicine, Institute of Clinical Medicine, Taipei Veterans General Hospital, National Yang Ming University, and b Department of Internal Medicine, National Taiwan University Hospital and National Taiwan University College of Medicine, Taipei, and c Department of Medicine, University of Hong Kong, Hong Kong , China
upregulation of renal TGF-  1 and activation of TGF-  /Smad signaling, which was blocked by treatment with PTX. Con-clusions: PTX may be an anti-fibrosis agent capable of in-hibiting renal fibrosis in a rat model of crescentic glomeru-lonephritis. Blockade of TGF-  1 expression and Smad2/3 activation may be a mechanism by which PTX inhibits renal fibrosis. Copyright © 2008 S. Karger AG, Basel
Introduction
Upregulation of pro-inflammatory cytokines and
ad-hesion molecules, excessive T cell and macrophage
infil-tration, and glomerular crescent formation with rapidly
progressive renal dysfunction are the features of
crescen-tic glomerulonephritis. In the past decades, the
patho-genic mechanisms of immune-mediated renal injury in
crescentic glomerulonephritis have been well elucidated
[1] ; however, little attention has been paid to the
develop-ment of renal fibrosis, another critical pathological
fea-ture in crescentic glomerulonephritis.
Pentoxifylline (PTX), a clinically available methyl
xanthine derivative, is a nonselective phosphodiesterase
Key Words
Pentoxifylline ⴢ Crescentic glomerulonephritis ⴢ
Transforming growth factor-beta ⴢ Epithelial-myofibroblast transition ⴢ Smads ⴢ Renal fibrosis
Abstract
Background/Aims: Pentoxifylline (PTX) has been shown to inhibit renal inflammation in a rat model of crescentic glo-merulonephritis. The present study investigated the role of PTX in renal fibrosis in rats with crescentic glomerulonephri-tis. Methods: A rat model of accelerated anti-glomerular basement membrane glomerulonephritis was induced and treated with PTX or vehicle control for 3, 7, 14 and 28 days. The therapeutic effect and mechanism of PTX on renal fibro-sis were examined by Northern blot and immunohisto-chemistry. Results: Diseased rats treated with vehicle con-trol developed a severe crescentic glomerulonephritis with progressive renal fibrosis identified by a marked accumula-tion of ␣ -SMA+ myofibroblasts and collagen matrix. This was associated with tubular epithelial-myofibroblast transition as evident by de novo expression of ␣ -SMA and a loss of E-cadherin on damaged tubular epithelial cells. Further stud-ies revealed that severe renal fibrosis was associated with
Received: April 16, 2008 Accepted: June 11, 2008 Published online: August 5, 2008
Nephrology
American Journal ofProf. Hui Y. Lan Department of Medicine © 2008 S. Karger AG, Basel
Ng /Chen /Tsai /Lan /Yang /Lan Am J Nephrol 2009;29:43–53
44
inhibitor with hemorheological activity and
anti-inflam-matory effects in vivo and in vitro [2] . It has been shown
that PTX exerts its anti-inflammatory activities in a
number of disease models including anti-GBM
crescen-tic glomerulonephritis [3–6] and human kidney diseases
[7–11] . However, it remains unclear whether the
thera-peutic effects of PTX on kidney diseases are also
associ-ated with inhibition of renal fibrosis and what
mecha-nisms may be involved.
In the current study, we found that tubular
epithelial-myofibroblast transition (EMT), a critical process in
re-nal fibrosis, occurred in a rat model of anti-GBM
cres-centic glomerulonephritis. Treatment with PTX was able
to block this process and inhibited renal fibrosis.
Block-ade of renal TGF-
 1 expression and inactivation of the
TGF-
 /Smad signaling pathway may be a mechanism by
which PTX blocks EMT and renal fibrosis in an
experi-mental model of rat crescentic glomerulonephritis.
Subjects and Methods
Immune Model of Crescentic Glomerulonephritis
Male Wistar rats (190–200 g) were obtained from the animal center of National Taiwan University Hospital. Experimental crescentic GN was induced as previously reported [5] . Briefly, an-imals were immunized by subcutaneous injection with 5 mg nor-mal rabbit IgG in Freund’s complete adjuvant. Five days later, groups of 6 animals were injected intravenously with 0.5 ml rabbit anti-rat GBM serum (2.5 ml/kg body weight, 12.5 mg IgG/ml), followed immediately by either PTX at a dose of 0.1 g/kg/day (Hoechst, Frankfurt, Germany) or vehicle (phosphate-buffered saline) via intravenous infusion over 1 h on a daily basis until be-ing killed at days 3, 7, 14 and 28. The experimental protocol was approved by the Animal Experimentation Committee, National Taiwan University.
Renal Histopathology
Kidney tissues for histological examination were fixed in 10% formalin and 4- m paraffin sections were stained with hema-toxylin and eosin or periodic acid-Schiff reagent (PAS). Glomeru-lar crescent formation and percentage of renal fibrosis in the cor-tical tubulointerstitium was scored on coded slides as described previously [5, 12] .
Antibodies
Antibodies used in this study included: 1A4, which specifi-cally recognized the smooth-muscle ␣ -actin isoform in rat, mouse and human (Sigma Chemical Co., St. Louis, Mo., USA); E-cad-herin (G-10), an epithelial marker on tubular epithelial cells (San-ta Cruz Biotechnology, Calif., USA); TGF-  1 and phosphorylated Smad 2/3 (Santa Cruz), and collagen III (Southern Biotech, Birmingham, Ala., USA). In addition, peroxidase-conjugated goat anti-mouse IgG, mouse peroxidase anti-peroxidase com-plexes, alkaline phosphatase-conjugated goat anti-mouse IgG and
mouse alkaline phosphatase anti-alkaline phosphatase complexes (APAAP) were purchased from Dakopatts (Glostrup, Denmark).
Immunohistochemistry
Double immunohistochemistry was performed using a micro-wave-based two-color immunostaining technique [12, 13] . Brief-ly, sections were dewaxed and treated with microwave oven heat-ing for 10 min in 10 m M sodium citrate pH 6.0 to retrieve antigens. Sections were then preincubated for 20 min in 10% fetal calf se-rum to block nonspecific binding, followed by the first primary antibody overnight at 4 ° C. After washing and inactivating en-dogenous peroxidase in 0.3% H 2 O 2 in methanol, sections were
incubated sequentially with the secondary antibody and then ter-tiary antibody for 1 h at room temperature and developed with diaminobenzidine to give a brown product. To block antibody cross-reactivity and inactivate endogenous alkaline phosphatase, sections were treated with microwave again and stained with the second primary antibody overnight at 4 ° C. Sections were then incubated with the alkaline phosphatase-conjugated goat anti-mouse, followed by a mouse APAAP for 1 h at room temperature, respectively. Finally, sections were developed with Fast Blue BB salt (Sigma) and examined under the microscope.
Semiquantitative Analysis of Immunohistochemical Staining
Total number of cells positive for ␣ -SMA and p-Smad2/3 within the tubulointerstitium were counted under 20 high-power field ( ! 40) by means of a 0.02-mm 2 graticule fitted in the eyepiece
of the microscope, and expressed as cells per mm 2 as previously
described [14] . For tubular epithelial cells that expressed TGF-  1 or were transitioning to a mesenchymal phenotype identified as double positive with ␣ -SMA and E-cadherin, at least 500 cortical tubules were scored and percent positive for TGF-  1 or double-positive for ␣ -SMA and E-cadherin was recorded as previously described [11] . All analyses were processed from the outer to inner cortex, avoiding large vessels and glomeruli. The large vessels, including the arterial walls, were excluded from the study. Be-cause of a diffuse nature in collagen III accumulation in the tu-bulointerstitium, the percent positive area was semiquantitative-ly scored as: score 1 ( ! 5%), score 2 (5–10%), score 3 (11–15%), score 4 (16–20%), score 5 (21–25%) and score 6 ( 1 25%). All analyses were performed on coded slides.
Renal RNA Extraction and Northern Blot Analysis
Renal RNA from the cortex was isolated using the guanidini-um thiocyanate/acid phenol method and analyzed by Northern blot as previously described [5] . The specific primer pairs for TGF-  1 were: upstream, 5 ⴕ -TCCACAGAGAAGAACTGCTG-3 ⴕ and downstream, 5 ⴕ -ACTTGCAGGAGCGCACAATC-3 ⴕ . The product was subsequently subcloned into the pGEM-dT vector (Promega, Madison, Wisc., USA). The cloned cDNAs were then linearized and used as templates for in vitro transcription of an-tisense digoxigenin-conjugated riboprobes, following the suppli-er’s instructions (Roche Molecular Biochemicals, Mannheim, Germany).
Statistical Analyses
Data were expressed as mean 8 SEM. All comparisons were done by one-way analyses of variance with Tukey’s comparison program from statistical package for social science (SPSS). p ! 0.05 was considered statistically significant.
Pentoxifylline Inhibits Renal Fibrosis Am J Nephrol 2009;29:43–53 45
Results
Effect of PTX Treatment on Fibrotic Changes in
Renal Histology
As reported previously [5] , rats injected with the
anti-rat GBM antiserum developed severe anti-GBM crescent
glomerulonephritis with massive inflammatory/immune
response, which is suppressed by PTX. The present study
focused on the therapeutic effect of PTX on renal fibrosis.
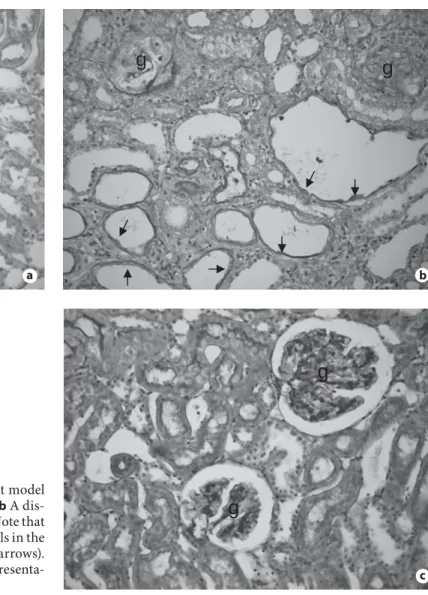
As shown in figure 1 , moderate-to-severe
glomeruloscle-rosis and tubulointerstitial fibglomeruloscle-rosis were developed in
dis-eased animals that had received the vehicle treatment,
which is inhibited by treatment with PTX. Interestingly,
severe tubulointerstitial fibrosis was associated with a
loss of normal tubular epithelial cell morphology,
becom-ing spindle-shaped cells laybecom-ing the highly dilated tubular
space ( fig. 1 b), which was also inhibited by PTX
treat-ment ( fig. 1 c). Immunohistochemically, severe renal
fi-brosis as identified by a marked accumulation of
␣ -SMA+
myofibroblasts and collagen type III was developed in
diseased rats treated with vehicle control, which was
sub-stantially blocked by the treatment with the PTX ( fig.
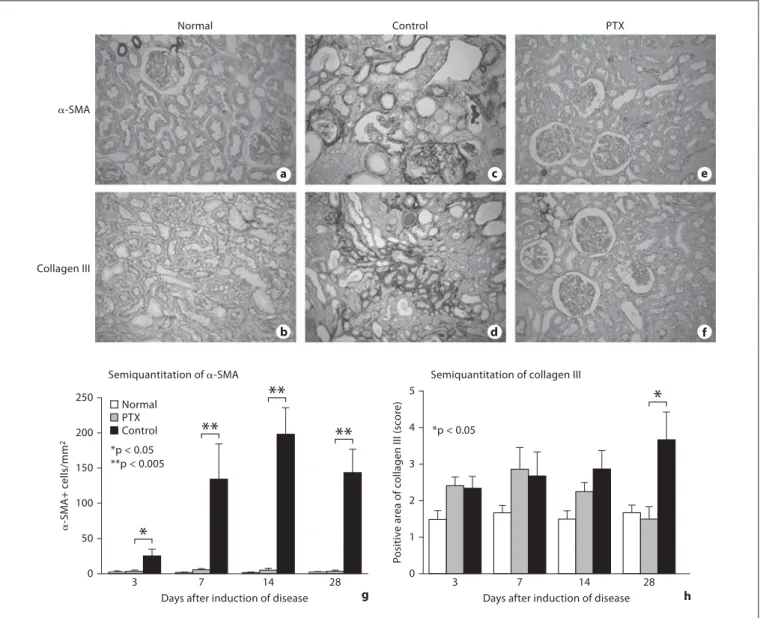
2 a–f). Semiquantitative analysis confirmed these
obser-vations ( fig. 2 g, h).
Effect of PTX Treatment on EMT and the
Development of Tubulointerstitial Fibrosis in
Anti-GBM Glomerulonephritis
It is now well recognized that EMT plays a role in
tu-bulointerstitial fibrosis [12, 15–17] . Thus, we examined
whether EMT occurs in tubulointerstitial fibrosis in the
anti-GBM glomerulonephritis model and if PTX
Fig. 1. Effect of PTX treatment on renal histology in a rat model of crescentic glomerulonephritis. a A normal rat kidney. b A dis-eased rat kidney treated with vehicle control for 28 days. Note that glomeruli are sclerosis (g) and many tubular epithelial cells in the dilated tubules become spindle-shaped in morphology (arrows).
c A diseased rat kidney treated with PTX for 28 days. Representa-tive pictures are stained with PAS. ! 200.
g
g
ag
g
bg
g
cNg /Chen /Tsai /Lan /Yang /Lan Am J Nephrol 2009;29:43–53
46
ment could block this process. As shown in figure 3 a,
two-color immunohistochemistry revealed that all
tubu-lar epithelial cells in normal rat kidneys highly expressed
E-cadherin (an epithelial phenotype marker) with
com-plete absence of
␣ -SMA (a myofibroblast marker) and
tu-bulointerstitial fibrosis. Strikingly, severe
tubulointer-stitial fibrosis associated with EMT was observed in
ve-hicle-treated rats (
fig. 3
b). The transformed tubular
epithelial cells showed de novo expression of
␣ -SMA with
a partial loss of E-cadherin, contributing to focal
accu-mulation of numerous
␣ -SMA+ myofibroblasts within
the area of severe tubulointerstitial fibrosis (
fig. 3
b),
which was blocked by PTX ( fig. 3 c). As shown in
fig-ure 3 d, semiquantitative analysis showed that a
signifi-cant EMT occurred at day 7 and this was associated with
an increase in interstitial
␣ -SMA+ myofibroblast
mulation and the development of collagen matrix
accu-mulation as shown in figures 2 and 3 b.
␣-SMA
Collagen III
Normal Control PTX
Semiquantitation of ␣-SMA Semiquantitation of collagen III 250 200 100 150 *p < 0.05 **p < 0.005 50 0 ␣ -SM A+ c e lls/mm 2
*
3Days after induction of disease 7
**
14**
28**
Normal PTX Control 5 4 2 3 *p < 0.05 1 0 P o sitiv e ar ea of c o llagen III (sc o re )Days after induction of disease
3 7 14
*
28 a c e b d f g hFig. 2. Effect of PTX treatment on accumulation of ␣ -SMA+ myo-fibroblasts and collagen III in a rat model of crescentic glomeru-lonephritis at day 28. a , b Normal rat kidney tissues stained with ␣ -SMA ( a ) or collagen III ( b ). c , d A diseased kidney treated with vehicle control and stained with anti- ␣ -SMA ( c ) or collagen III ( d ) antibodies. e , f A diseased kidney with PTX treatment and
stain-ing with anti- ␣ -SMA ( e ) or collagen III ( f ) antibodies. g , h Semi-quantitative analysis for ␣ -SMA+ cells and collagen III accumula-tion within the tubulointerstitium. Each bar represents the mean 8 SEM for a group of 6 rats. * p ! 0.05, * * p ! 0.005 compared with the PTX treatment. ! 200.
Pentoxifylline Inhibits Renal Fibrosis Am J Nephrol 2009;29:43–53 47
Inhibition of TGF-

Expression and Smad2/3
Activation Is a Mechanism by Which PTX Attenuates
Renal Fibrosis in a Rat Model of Anti-GBM
Glomerulonephritis
We next examined the mechanisms whereby PTX
in-hibits renal fibrosis. Upregulation of renal TGF-
 and
activation of its signaling pathway Smad2/3 have been
shown to play a critical role in renal fibrosis [14] . Thus,
expression of renal TGF-
 1 and activation of Smad2/3
were examined. As shown in figure 4 , Northern blot
anal-ysis demonstrated that treatment with PTX showed a
sig-nificant reduction in renal TGF-
 mRNA expression
a
c
b
Fig. 3. Two-color immunohistochemical staining shows that treatment with PTX blocks tubular EMT in a rat model of cres-centic glomerulonephritis. Brown represents E-cadherin+ tubu-lar epithelial cells and blue represents ␣ -SMA myofibroblasts. The mixed color (purple) demonstrates EMT. a A normal rat kidney shows that all E-cadherin+ tubular epithelial cells are absent with
␣ -SMA staining. b A control-treated diseased kidney shows that most E-cadherin tubular epithelial cells in the area with massive
interstitial ␣ -SMA+ myofibroblast accumulation are spindle-shaped (EMT) with myofibroblast phenotype (E-cadherin+ ␣ -SMA+), which is further illustrated in the inserted area. c A dis-eased kidney treated with PTX shows that all E-cadherin+ tubu-lar epithelial cells are absent with ␣ -SMA and few ␣ -SMA+ myofibroblasts are found in tubulointerstitium. d Semiquantita-tive analysis. Each bar represents the mean 8 SEM for a group of 6 rats. * * p ! 0.005 compared to the control animals. ! 200.
PTX Control **p < 0.005 10 20 0 ␣ -SM A + E-cadherin+ tubules (%)
Days after induction of disease
**
3 7**
14**
28 d Quantitation of EMTNg /Chen /Tsai /Lan /Yang /Lan Am J Nephrol 2009;29:43–53
48
from day 3 to day 28. This was further confirmed by
im-munostaining with the anti-TGF-
 1 antibody and we
found that upregulation of renal TGF-
 1 in the diseased
kidney of control animals was significantly suppressed
by treatment with PTX ( fig. 5 ).
A close association between upregulation of TGF-
 1
and the development of EMT and tubulointerstitial
fibro-sis was further examined by two-color
immunohisto-chemistry with the anti-TGF-
 1 and ␣ -SMA or collagen
III antibodies. As shown in figure 6 , upregulation of
tu-bular TGF-
 1 was closely associated with the de novo
expression of
␣ -SMA and collagen III by tubular
epithe-lial cells (EMT), contributing to local accumulation of
␣ -SMA+ myofibroblasts and collagen III and the
devel-opment of severe tubulointerstitial fibrosis ( fig. 6 a, b),
which was inhibited by treatment with PTX ( fig. 6 c, d).
It is well known that TGF-
 1 signals through its
down-stream signaling proteins Smad2/3 to mediate fibrosis.
We thus examined the activation of Smad2/3 by their
nu-clear location using the anti-phosphorylated Smad2/3
antibody. As demonstrated in figure 7 , in normal kidney,
there was a weak activation of phospho-Smad2/3 in some
tubular epithelial cells and glomerular cells as identified
by the nuclear pattern of localization ( fig.7 a, d),
indicat-ing a low level of TGF-
 signaling. In contrast, diseased
rats treated with the vehicle control exhibited a marked
increase in a nuclear staining pattern of phospho-Smad2/3
in both glomerular and tubulointerstitial areas with
se-vere fibrosis ( fig. 7 b, d), which was largely inhibited by
PTX treatment ( fig. 7 c, d).
Discussion
The present study showed that PTX was also an
effec-tive agent capable of anti-fibrosis in an immunologically
induced glomerulonephritis, in addition to its
anti-in-flammatory properties as reported previously [2–9] . The
anti-fibrotic effect of PTX in a rat model of anti-GBM
crescentic glomerulonephritis may be associated with an
inhibition of tubular EMT and the production of collagen
matrix. Of significance, inhibition of renal TGF-
 1
ex-pression and Smad2/3 activation may be a critical
mech-anism by which PTX inhibits renal fibrosis.
Increasing evidence shows that tubular EMT is a
mechanism of renal tubulointerstitial fibrosis in both
an-imal models and human chronic kidney disease [12, 15–
20] . We have previously shown that EMT also
partici-pates in the formation and evolution of glomerular
cres-cents in rat anti-GBM glomerulonephritis [21] . In the
present study, we further showed that EMT was
associ-ated with progressive tubulointerstitial fibrosis in a rat
model of anti-GBM glomerulonephritis. Indeed, EMT
occurring around day 3–7 was associated with the
devel-opment of peritubular
␣ -SMA myofibroblast
accumula-tion and preceded the development of severe
tubuloin-terstitial fibrosis at day 28, suggesting that EMT is a key
process in producing extracellular matrix, leading to an
a b TGF- GAPDH TGF- GAPDH TGF- GAPDH TGF- GAPDH 3 days 1 week 2 weeks 4 weeks Normal Control PTX 0 1 2 +
* *
*
*
+ + + TG F - 1/GAPDH 3 3 7 14 Days of nephritis 28 Quantitative analysisFig. 4. Northern blot analysis shows that PTX treatment inhibits renal TGF-  1 mRNA expression in a rat model of crescentic glo-merulonephritis. a Each lane represents one rat kidney and data represents for a group of 6 rats. Representative Northern blots demonstrate that renal TGF-  1 mRNA is significantly upregu-lated in control animals, which is blocked in animals treated with PTX. b Quantitative analysis. Each line symbol represents the mean 8 SEM for a group of 6 rats. * p ! 0.05 PTX-treated vs. ve-hicle-treated nephritic rats; + p ! 0.05 vehicle-treated nephritic
rats vs normal control rats. y = Normal control rats; $ = vehicle-treated nephritic rats; ) = PTX-vehicle-treated nephritic rats.
Pentoxifylline Inhibits Renal Fibrosis Am J Nephrol 2009;29:43–53 49
advanced renal fibrosis. Furthermore, consistent with the
previous finding that PTX treatment attenuates
myofi-broblast and collagen matrix accumulation in
obstruc-tive kidney disease [22] , treatment with PTX was capable
of inhibiting the EMT process, thus preventing renal
fi-brosis. Therefore, results from the present study provided
new evidence for the antifibrotic effect of PTX.
It has been shown that PTX is able to inhibit fibroblast
growth factor-2-dependent EMT and collagen matrix
a b
c
synthesis in vitro and attenuates tubulointerstitial
fibro-sis in a rat model of obstructive kidney by blocking
Smad3/4-dependent transcription [22, 23] . It is now well
accepted that TGF-
 /Smad signaling plays a critical role
in the development of renal fibrosis [24] . Inhibition of
re-nal fibrosis in a mouse model of autoimmune
glomerulo-nephritis by a gene transfer of Smad7 demonstrates a role
for the TGF-
 /Smad pathway in the development of renal
fibrosis in crescentic glomerulonephritis [25] . In the
Fig. 5. Immunohistochemistry shows that PTX treatment inhibits renal TGF-  1 protein expression in a rat model of crescentic glo-merulonephritis. a A normal rat kidney. b A diseased rat kidney treated with vehicle control for 28 days. c A diseased rat kidney
treated with PTX for 28 days. d Semiquantitative analysis. Each bar represents the mean 8 SEM for a group of 6 rats. * p ! 0.05, * * p ! 0.005 compared with the control animals. ! 200.
d Normal PTX Control 60 40 20 0 TG F - + tubules (%)
Days after induction of disease Semiquantitation *p < 0.05 **p < 0.005 3
*
7**
14**
28**
Ng /Chen /Tsai /Lan /Yang /Lan Am J Nephrol 2009;29:43–53
50
ent study, we added new data that PTX exhibited its
an-tifibrotic effect on immunologically mediated crescentic
glomerulonephritis by suppressing renal TGF-

expres-sion and inactivating the downstream of TGF-

signal-ing pathway, Smad2/3. Indeed, marked upregulation of
TGF-
 1 mRNA and protein was evident in the diseased
kidney with strong activation of Smad2/3 and severe
re-nal fibrosis. Upregulation of TGF-
 1 by tubular
epithe-lial cells was associated with EMT as evident by
co-ex-pression of TGF-
 1 and ␣ -SMA or collagen III.
Treat-*
*
*
*
*
*
a b c d Control PTXFig. 6. Two-color immunohistochemical staining shows a close localization of tubular TGF-  1 expression and ␣ -SMA+ myofi-broblast and collagen III accumulation in a rat model of crescen-tic glomerulonephritis at day 28. Brown represents ␣ -SMA+ or collagen III+ staining and blue represents TGF-  1+ cells. a A rep-resentative diseased kidney treated with vehicle control and stained with ␣ -SMA and TGF-  1. b A representative diseased kidney treated with vehicle control and stained with collagen III and TGF-  1. c A representative diseased kidney treated with PTX and stained with ␣ -SMA and TGF-  1. d A representative diseased
kidney treated with PTX and stained with collagen III and
 1. Two-color immunostaining shows that TGF-  1 (blue) is strongly expressed by tubular epithelial cells in the area with mas-sive interstitial ␣ -SMA+ myofibroblast accumulation ( a ) and col-lagen III deposition ( b ). Note that some tubular epithelial cells with strong TGF-  1 expression are also ␣ -SMA+ with myofibro-blast phenotype ( * ) and produce collagen III ( * ), which is enlarged in the inserted pictures. In contrast, treatment with PTX blocks TGF-  1 expression and accumulation of ␣ -SMA+ myofibroblasts and collagen III ( c , d ). ! 200.
Pentoxifylline Inhibits Renal Fibrosis Am J Nephrol 2009;29:43–53 51
ment with PTX was able to significantly inhibit TGF-
 1
mRNA and protein expression, thereby preventing EMT
and collagen matrix expression. Further study showed
that the anti-fibrotic effect of PTX was associated with
blockade of the TGF-
 1 signaling pathway because a
marked Smad2/3 activation was significantly inhibited in
the diseased kidney treated with PTX. Since TGF-
 /
Smad signaling is a major pathway leading to EMT and
renal fibrosis [15, 24, 26, 27] , blockade of this pathway is
able to suppress TGF-
 1-induced EMT and renal fibrosis
in vivo and in vitro [14, 27] . Thus, inhibition of TGF-

1-dependent Smad2/3 activation could be a central
mecha-nism by which PTX attenuates renal fibrosis in a rat
mod-el of anti-GBM crescentic glomerulonephritis.
It is also possible that inhibition of renal TGF-
 1 and
its downstream mediator CTGF expression by PTX may
be a consequence of the anti-inflammatory effect of PTX
since this is associated with the anti-inflammatory effect
a b
c
Fig. 7. Immunohistochemistry shows that PTX treatment inhibits Smad2/3 activation in a rat model of crescentic glomerulonephri-tis. Activation of TGF-  signaling is identified by phosphorylated Smad2/3 nuclear location. a A normal rat kidney. b A diseased rat
kidney treated with vehicle control for 28 days. c A diseased rat kidney treated with PTX for 28 days. d Semiquantitative analysis. Each bar represents the mean 8 SEM for a group of 6 rats. * * p ! 0.005 compared with the control animals. ! 200.
d Normal PTX Control 400 300 200 100 0 p -Smad2/3+ c e lls/mm 2
Days after induction of disease **p < 0.005 3
**
7**
14**
28**
Phospho-Smad2/3+ cellsNg /Chen /Tsai /Lan /Yang /Lan Am J Nephrol 2009;29:43–53
52
References
1 Tipping PG, Kitching AR, Cunningham MA, Holdsworth SR: Immunopathogenesis of crescentic glomerulonephritis. Curr Opin Nephrol Hypertens 1999; 8: 281–286. 2 Lin SL, Chen YM, Chiang WC, Tsai TJ, Chen
WY: Pentoxifylline: a potential therapy for chronic kidney disease. Nephrology (Carl-ton) 2004; 9: 198–204.
3 Chen YM, Chien CT, Hu-Tsai MI, Wu KD, Tsai CC, Wu MS, Tsai TJ: Pentoxifylline at-tenuates experimental mesangial prolifera-tive glomerulonephritis. Kidney Int 1999; 56: 932–943.
4 Wang W, Zolty E, Falk S, Basava V, Reznikov L, Schrier R: Pentoxifylline protects against endotoxin-induced acute renal failure in mice. Am J Physiol Renal Physiol 2006; 291: F1090–F1095.
5 Chen YM, Ng YY, Lin SL, Chiang WC, Lan HY, Tsai TJ: Pentoxifylline suppresses renal tumour necrosis factor-alpha and amelio-rates experimental crescentic glomerulone-phritis in rats. Nephrol Dial Transplant 2004; 19: 1106–1115.
6 Lin SL, Chen YM, Chien CT, Chiang WC, Tsai CC, Tsai TJ: Pentoxifylline attenuated the renal disease progression in rats with remnant kidney. J Am Soc Nephrol 2002; 13: 2916–2929.
7 Demir E, Paydas S, Balal M, Kurt C, Sert-demir Y, Erken U: Effects of pentoxifylline on the cytokines that may play a role in rejec-tion and resistive index in renal transplant recipients. Transplant Proc 2006; 38: 2883– 2886.
8 Navarro JF, Mora C, Muros M, Garcia J: Ad-ditive antiproteinuric effect of pentoxifyl-line in patients with type 2 diabetes under angiotensin II receptor blockade: a short-term, randomized, controlled trial. J Am Soc Nephrol 2005; 16: 2119–2126.
9 Chen YM, Lin SL, Chiang WC, Wu KD, Tsai TJ: Pentoxifylline ameliorates proteinuria through suppression of renal monocyte che-moattractant protein-1 in patients with pro-teinuric primary glomerular diseases. Kid-ney Int 2006; 69: 1410–1415.
10 Rodriguez-Moran M, Gonzalez-Gonzalez G, Bermudez-Barba MV, Medina de la Garza CE, Tamez-Perez HE, Martinez-Martinez FJ, Guerrero-Romero F: Effects of pentoxi-fylline on the urinary protein excretion pro-file of type 2 diabetic patients with micro-proteinuria: a double-blind, placebo-con-trolled randomized trial. Clin Nephrol 2006; 66: 3–10.
11 Ducloux D, Bresson-Vautrin C, Chalopin J: Use of pentoxifylline in membranous ne-phropathy. Lancet 2001; 357: 1672–1673. 12 Ng YY, Huang TP, Yang WC, Chen ZP, Yang
AH, Mu W, Nikolic-Paterson DJ, Atkins RC, Lan HY: Tubular epithelial-myofibroblast transdifferentiation in progressive tubuloin-terstitial fibrosis in 5/6 nephrectomized rats. Kid Int 1998; 54: 864–876.
13 Lan HY, Mu W, Nikolic-Paterson DJ, Atkins RC: A novel, simple, reliable, and sensitive method for multiple immunoenzymed stain-ing: use of microwave oven heating to block antibody crossreactivity and retrieve anti-gens. J Histochem Cytochem 1995; 43: 97– 102.
14 Lan HY, Mu W, Tomita N, Huang XR, Li JH, Zhu HJ, Morishita R, Johnson RJ: Inhibition of renal fibrosis by gene transfer of inducible Smad7 using ultrasound-microbubble sys-tem in rat UUO model. J Am Soc Nephrol 2003; 14: 1535–1548.
15 Lan HY: Tubular epithelial-myofibroblast transdifferentiation mechanisms in proxi-mal tubule cells. Curr Opin Nephrol Hyper-tens 2003; 12: 25–29.
16 Liu Y: Epithelial to mesenchymal transition in renal fibrogenesis: pathologic signifi-cance, molecular mechanism, and therapeu-tic intervention. J Am Soc Nephrol 2004; 15: 1–12.
17 Kalluri R, Neilson EG: Epithelial-mesenchy-mal transition and its implications for fibro-sis. J Clin Invest 2003; 112: 1776–1784. 18 Nishitani Y, Iwano M, Yamaguchi Y, Harada
K, Nakatani K, Akai Y, Nishino T, Shiiki H, Kanauchi M, Saito Y, Neilson EG: Fibroblast-specific protein 1 is a Fibroblast-specific prognostic marker for renal survival in patients with IgAN. Kidney Int 2005; 68: 1078–1085. 19 Aresu L, Rastaldi MP, Scanziani E, Baily J,
Radaelli E, Pregel P, Valenza F: Epithelial-mesenchymal transition (EMT) of renal tu-bular cells in canine glomerulonephritis. Virchows Arch 2007; 45: 1937–1942.
20 Jinde K, Nikolic-Paterson DJ, Huang XR, Sakai H, Kurokawa K, Atkins RC, Lan HY: Tubular phenotypic change in progressive tubulointerstitial fibrosis in human glomer-ulonephritis. Am J Kidney Dis 2001; 38: 761– 769.
of PTX in kidney diseases [5, 9, 22] . Indeed, an increase
in pro-inflammatory cytokines (IL-1, TNF
␣ ) within
im-mune-mediated disease kidney is able to stimulate
in-flammatory cells (T cells and macrophages) and resident
kidney cells (tubular cells and fibroblasts) to produce
growth factors such as TGF-
 1, which, in turn, can
stim-ulate tubular epithelial cells to undergo the EMT process
and activate resident renal fibroblasts to produce
extra-cellular matrix. Our previous finding of IL-1-induced
EMT via the TGF-
 -dependent mechanism supports this
notion [28] . Once renal inflammation is inhibited, as a
result of PTX treatment, upregulation of renal TGF-
 1
and activation of its downstream Smad signaling
path-way could be suppressed. Therefore, PTX may exert its
renoprotective effect by blocking both renal
inflamma-tion and fibrosis.
Acknowledgements
This study was supported by grants from the Research Grant Council of Hong Kong (GRF 759206 and 768207), Research and Conference Grant of Hong Kong University (200511159199), and research grant of Taiwan (NSC94-2314-B-075-051).
Conflict of Interest Statement
The results presented in this paper have not been published previously in whole or part, except in abstract format.
Pentoxifylline Inhibits Renal Fibrosis Am J Nephrol 2009;29:43–53 53 21 Ng YY, Fan JM, Mu W, Nikolic-Paterson DJ,
Yang WC, Huang TP, Atkins RC, Lan HY: Glomerular epithelial-myofibroblast trans-differentiation in the evolution of glomeru-lar crescent formation. Nephrol Dial Trans-plant 1999; 14: 1860–1872.
22 Lin SL, Chen RH, Chen YM, Chiang WC, Lai CF, Wu KD, Tsai TJ: Pentoxifylline attenu-ates tubulointerstitial fibrosis by blocking Smad3/4-activated transcription and profi-brogenic effects of connective tissue growth factor. J Am Soc Nephrol 2005; 16: 2702– 2713.
23 Strutz F, Heeg M, Kochsiek T, Siemers G, Zeisberg M, Müller GA: Effects of pentoxi-fylline, pentifylline and gamma-interferon on proliferation, differentiation, and ma-trix synthesis of human renal fibroblasts. Nephrol Dial Transplant 2000; 15: 1535– 1546.
24 Wang W, Koka V, Lan HY: Transforming growth factor-beta and Smad signalling in kidney diseases. Nephrology (Carlton) 2005; 10: 48–56.
25 Ka SM, Huang XR, Lan HY, Tsai PY, Yang SM, Shui HA, Chen A: Smad7 gene therapy ameliorates an autoimmune crescentic glo-merulonephritis in mice. J Am Soc Nephrol 2007; 18: 1777–1788.
26 Fan JM, Ng YY, Hill PA, Nikolic-Paterson DJ, Mu W, Atkins RC, Lan HY: Transform-ing growth factor-beta regulates tubular epi-thelial-myofibroblast transdifferentiation in vitro. Kidney Int 1999; 56: 1455–1467. 27 Li JH, Zhu HJ, Huang XR, Lai KN, Johnson
RJ, Lan HY: Smad7 inhibits fibrotic effect of TGF-Beta on renal tubular epithelial cells by blocking Smad2 activation. J Am Soc Nephrol 2002; 13: 1464–1472.
28 Fan JM, Huang XR, Ng YY, Nikolic-Paterson DJ, Mu W, Atkins RC, Lan HY: Interleukin-1 induces tubular epithelial-myofibroblast transdifferentiation through a transforming growth factor-beta1-dependent mechanism in vitro. Am J Kidney Dis 2001; 37: 820–831.