L-nw*, GST MI genetic polymorphism and hepatocellular carcinoma
risk among chronic hepatitis B carriers
ill ~,tttet nr ;tsscss ;tssoetations between the genetic polymorphism of I,-nr~ and glutathione S-transfcrase M 1 (GST Ml) .IIIJ Ihi. risk i)t hcpatoccllular carcinoma (IICC). :I total 01. 46 surgically treated HCC patients who were seropositive in hepa- ;,iis 1< htli-f:ll~i~ :nltigc‘n (Il1W.g) :rntJ XX tlBsAp-positive controls were recruited for this study. l.-rn~~~ and GST Ml genetic t~oiyttn>rpitisnt w;ts cx;rrnined using a polymcrase chain reaction-hased restriction fragment length polymorphism assay on !%‘A cx~r;tctc;i tram liver ,rnd pcriphcrai blood samples. There was no significant difference in GST M 1 genotypes between I I(‘(. p,~tt~~n:~ ;mtl nnnchcd L~ontrols. A gene dosage trend 01‘ association with I-ICC risk was observed for L-my genotype. ‘l‘tlc &xc-response relationship remained statistically signilicant in the multiple logistic regression analysis.
AAYI ,~rJr tlq~~itrs I3 intsction: I.-ttrw; (;lutathione S-transt’erase MI: Hepatocellular carcinoma
f fepatoccllular carcmomn (I-ICC) is the leading 13ust- ijt r2rlccr death in Taiwan, accounting for ap- proximately~ 3465 deaths per year [ I I]. This means 1 hat appr~~xtmately 2 I .3% of all cancer deaths in
iili\%m ;trLs due to IKYY Epidemiological studies ha1.c sttggcatcd that environmental factors play a ttutjor rofc in the dcvclopment of HCC [271. Such f’acft~ inilude hepatitis virus infection, exposure to i,hemica! carcinoperts such as aflatoxin and cigarette
smoking, and hormone levels. However, the molecu- lar mechanism of how these factors are related to the development of HCC remains to be elucidated.
The induction of cancers in humans and in ani- mals proceeds through a complex series of reactions and processes, subject to and controlled by a number of modifying factors. As a result, individual suscep- tibility to cancer is due to a combination of several factors. including differences in metabolism, DNA repair. altered proto-oncogene or tumor suppressor gene expression, hormonal, immunologic and nutri- tional status. Thus, susceptibility is probably the sin- Kyle most important factor relative to initiation of c
172 L.-L. Hsieh et al. /Cancer Letters 103 (1996) 171-l 76
tively small percentage of people who develop cancer in an exposed cohort.
Genetic susceptibility to chemically induced HCC determines the propensity to convert hepatocarcino- genie chemicals into active metabolites and to detox- ify active metabolites by glutathione conjugation. The glutathione S-transferases (GSTs) are a unique group of multifunctional isozymes which catalyze the conjugation of reduced glutathione (GSH) with a variety of electrophilic compounds, including car- cinogens and cytotoxic drugs 171. In humans, this supergene family is divided on the basis of chromo- somal location and sequence homology, into four classes named CL, ,u, JZ and 8 [16]. Within the GST p family, one member, the GST Ml enzyme has been shown to catalyze the conjugation of potentially cyto- and genotoxic epoxides such as aflatoxin B, (AFB,)- 8,9-epoxide [ 171. It has been shown that a deficiency of GST Ml enzyme activity is associated with a null genotype at this gene locus [20]. A series of reports suggested that the GST Ml null genotypelpheno- type is associated with susceptibility to cancer, espe- cially to cancer related to cigarette smoking [ 10,14,
19,211.
Activating proto-oncogenes, and turning them into oncogenes, is an important mechanism of tumor ini- tiation and progression [2]. A series of reports sug- gested that allelic variants of proto-oncogenes, such as H-rus and L-myc, might predispose individuals towards tumor development [3,18,23-251, although the hypothesized relationship between allelic variants of proto-oncogenes and cancer susceptibility does not yet have a mechanistic basis. Therefore, alleles of pro-oncogenes may serve as markers of genetic sus- ceptibility to cancer. Recently, a study, based on a small number of HCC cases, has suggested that per- sons with the SS genotype of L-myc may be pro- tected against HCC [25].
In this study, we have examined the relationship between the homozygous null genotype of GST Ml, hepatitis B virus infection and L-myc genotype in the development of HCC.
2. Materials and methods
2.1. Design of the case-control study
Seventy HCC patients at Lin-Kou Chang Gung
Memorial Hospital from September 1990 to Septem- ber 1992 were recruited as the case group. Most (54170 = 77%) of them were chronic carriers of hepatitis B surface antigen (HBsAg). Among them, 85.2% (46) of HBsAg-positive HCC cases were male. A total of 88 male HBsAg carriers who were frequency-matched with HCC cases on age were also recruited as the control group.
High molecular weight DNA was purified from surgically removed non-tumor tissues for the cases and from the peripheral blood samples of the healthy controls by digestion with proteinase K and extrac- tion with phenol/chloroform, as described [9,15].
2.2. PCR analysis of GST Ml genotype
A simple assay based on PCR technology has been developed [8] to determine the presence or ab- sence of GST Ml. The following approach was used in this study: briefly, genomic DNA (1 pug) was added to a PCR mix containing 200 ng of each primer for GST Ml (5’-CTGCCCTACTTGAm- GATGGG-3’ and 5’-CTGGATTGTAGCAGATCA- TGC-3’) and P-globin (5’-ACACAACTGTGTTC- ACTAGC-3’ and 5’-CAACTTCATCCACGTTC- ACC-3’) as the internal control, 80 mM dNTP, 1 unit of Taq polymerase (Promega Corp., Madison, WI, USA) and PCR buffer in a total volume of 25 ~1. The PCR products were electrophoresed on 8% polyacryl- amide gels, stained with ethidium bromide and pho- tographed under UV light.
2.3. PCR analysis of L-myc genotype
An EcoRI polymorphism of the L-myc proto- oncogene is located in the second intron [ 131. Genomic DNA was therefore amplified using primers that flank the polymorphic EcoRI site in the gene as described [25]. Genomic DNA (1 pug) was added to a PCR mix containing 200 ng of each primer (5’. AGCAGAGCTCACCCAATAGG-3’ and 5’-CCA- TTGTGTGGACAATCGCAT-3’), 40 mM dNTP,
1 unit of Taq polymerase and PCR buffer in 25~1. The PCR products were purified with ammonium acetate and isopropanol, digested with EcoRI at 37°C overnight, electrophoresed on 8% polyacrylamide gels, stained with ethidium bromide and photo- graphed under UV light.
2.4. Serological anul-ysis fbr heputitis virus infectiort Serological analysis for hepatitis B/C virus infec- tion was carried out as previously described 1221. HBsAg was analyzed using commercially available RIA kits (Abbott Laboratories, Chicago). Anti-HCV vt’as assayed using a commercial available EIA (Abbott HCV EIA). Initially, reactive serum speci- mens were retested in duplicate and repeatedly reac- tive samples were defined as possibly positive. Posi- tive samples were considered as true positive if con- firmed by the more specific second-generation assay using synthetic peptides from both core and non- structural regions of HCV (UBI HCV EIA; United Biomedical, New York).
Statistical analysis was done using x2 test with or without Yate’s correction, Fisher’s exact test, and logistic regression analysis when appropriate.
3. Results
I’he mean age it standard deviation were 47.3 -I 12.8 for HBsAg-positive HCC cases and 50.8 2 1 1.3 for age-matched healthy HBsAg carriers in the pres- ent study. Anti-HCV was detected in 6.5% (3/46) of HBsAg-positive HCC cases and in 0% (O/88) of healthy HBsAg carriers.
.i.2 GS7’MI genotype and NCC’
Fig. 1 shows the 273 bp DNA fragment amplified from the exon 4-5 region of GST Ml in subjects classified as GST M 1 positive. Examples of GST Ml null genotype, identified by the absence of this frag- ment, are also shown. Amplification of the 100 bp fragment from the p-globin gene that served as inter- nal control was observed in all subjects studied (Fig.
I I
Table 1 shows the distribution of the GST Ml genotypes detected in these study groups. The fre- quency of GST Ml null genotype in the HBsAg-
positive HCC cases as compared with that in the healthy HBsAg carriers was not significantly differ-
273 bp
1OObp
Fig. 1. Representative PCR-RFLP analysis of the WT Ml gene on the lICC patients and controls. Samples of DNA were ampli-
fied using PCR, electrophorcsed in polyacrylamide gels, and
stained with cthidium bromide. The 273-bp DNA fragment corre- sponds to the GST M 1, while the IOO-bp DNA fragment corre- sponds to thep-globin gene as the internal control.
ent. The odds ratio was 1 .O (95% confidence interval (CI) 052.3).
3.3. L-nay genotype and HCC
The polyacrylamide gel band patterns for each L- rqc EcoRI genotype are shown in Fig. 2. Based on PCR and EcoRI digestion, two alleles are apparent: a 145 bp fragment corresponding to the ‘L’ allele which lacks the EcoRI site, and paired 104 bp and 41 bp fragments from the ‘S’ allele, produced from the cut at the EcoRI site. The three genotypes are LL (145 bp fragment only), LS (145 bp, 104 bp, and 41 bp fragments) and SS (104 bp and 41 fragments only).
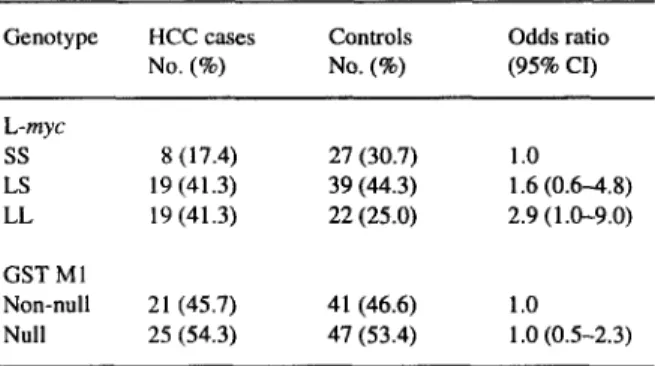
Among the healthy HBsAg carriers, the distribu- tion of L-myc genotype was LL 25.0% (22/88), LS 44.3% (39/88), and SS 30.7% (27188) (Table 1). Among HBsAg-positive HCC cases, the distribution of L-myc genotype was LL 41.3% (19/46), LS 41.3% (19/46), and SS 17.4% (g/46). Although this distri- bution does not differ significantly from that of the
174 L.-L. Hsieh et al. /Cuncer Letters 103 (1996) 171-176
Table 1 Table 2
The distribution of L-myc and GST Ml genotypes in HCC cases and controls
Odds ratios and 95% confidence interval for LS and LL genotype of L-myc gene according to age, and GST MI status
Genotype HCC cases No. (%) Controls No. (6) Odds ratio (95% CI) L-myc ss S(l7.4) 27 (30.7) 1.0 LS 19 (41.3) 39(44.3) 1.6(0.6-4.8) LL 19 (41.3) 22(25.0) 2.9(1.0-9.0) GSTMl Non-null 21 (45.7) 41 (46.6) 1.0 Null 25 (54.3) 47 (53.4) 1 .O (0.5-2.3)
controls (x2 = 4.7,
P = 0.096),
a gene dosage trend was observed(P
= 0.04 based on trend test). The odds ratio was 1.6 (95% CI = 0.6-4.8) for LS geno- type and 2.9 (95% CI = 1.0-9.0) for the LL geno- type, respectively. A similar gene dosage trend has only been observed for age groups younger than/ -145
bp
! -104
bp
-
41 bp
Fig. 2. Representative PCR-RFLP analysis of the L-myc gene on the HCC patients and controls. Samples of DNA were amplified using PCR, digested with EcoRI, electrophoresed in polyaclyla- mide gels, and stained with ethidium bromide. The 145-bp DNA fragment corresponds to the L allele which does not have an EcoRI site, while the paired 104-bp and 41-bp fragments corre- spond to the S allele which has the EcoRI site.
Variables Odds ratio (95% confidence interval) P value ss LS LL (referent) Age 650 >50 GST Ml Null Non-null 1.0 2.5 (0.6-I 1.2) 4.5 (0.9-23.0) 0.045 1.0 0.8 (0.2-4.8) 1.8 (0.3-9.8) 0.530 1.0 0.9(0.211.2) 2.1 (0.5-9.3) 0.277 1.0 3.1 (0.6-17.0) 4.4 (0.7-30.0) 0.099
50 years but not in age groups over 50 years or either type of GST Ml group (Table 2).
To fully evaluate the factors affecting an individ- ual who has HCC, logistic regression was used. After correction for the effects of age and GST Ml geno- type, the LL genotype of L-myc remained signifi- cantly related to HCC with an odds ratio of 3.1 (95% CI 1.8-5.1) (Table 3). However, the GST Ml geno- type was not found to be associated with HCC.
4. Discussion
Chronic hepatitis virus infection has long been suggested to play a major role in the development of HCC [5]. In this study, 70 HCC patients at Lin-Kou Chang Gung Memorial Hospital from September
Table 3
Logistic regression analysis of multiple risk factors of HCC
Variables Odd ratios 95%CI
L-myc genotype ss 1.0 LS 1.8 1 .I-2.9 LL 3.1 1.X-5.1 GST M 1 genotype Null 1.0 Non-null 0.9 0.4-2.0 Age (years) >50 1.0 <50 1.8 0.8-3.7
1990 to September 1992 were recruited as the case group. Among them, 77. I % were carriers of HBsAg. This percentage is similar to the HBsAg carrier rate of HCC exposure in Taiwan (83.5%) reported previ- ously [4]. The male/female sex ratio was 4.8 (58/12). This ratio IS close to previous reports [27]. Anti-HCV was detected in 18.6% (13/70) of the HCC cases, which is also consistent with previous reports 16,261. A final of 46 male HBsAg-positive HCC cases were qclected for the present study. A total of 88 healthy male HBsAg carriers who were matched with HCC cases on age were also recruited as the control group.
Increasing attention has recently been paid to the role of individual susceptibility in the pathogenesis of cancer. Recent studies suggest that GST Ml and L- rnyc genotype may be important indicators of genetic i;usceptihility to cancer. This case-control study shows a gene dosage trend of L-my genotype asso- ciation with HCC. Compared to the healthy HBsAg carriers, HBsAg-positive HCC cases have 1.6 and 2.9 the odds of carrying the LS and LL genotype, rc- spectively. These findings are similar to other reports linking I,-wJY~ genotype with cancer susceptibility including HCC [25]. A similar gene dosage trend has only been found in age groups younger than 50 years groups but not in age groups over 50 years or either type 01‘ GST Ml group. These observations suggest that I,-m~c, genotype is a useful genetic susceptibility marker of HCC.
Logistic regression was used to fully evaluate the t’actors affecting an individual with HCC. After the effects of age and GST Ml genotype have been cor- rected for. the LL genotype of I,-myc was still sig- nificantly related to HCC with an odds ratio of 3.1 i%%> c*i 1 .x--s. Ii.
Two possible explanations can be proposed for :hese findings. The first is a selection bias as indi- vtduals who were not surgical candidates had been excluded from this study. An alternative explanation is that HBsAg-positive individuals with the 1-L genotype arc more susceptible to develop HCC. I,- rttvc. expressed both in normal liver tissue and in liver tumors ] 121, appears to be a plausible candidate as ;I susceptibility gene. However, a functional dif- ference between the proteins encoded by S and L alleles and the role of L,-mpc in HCC has not yet been described. It is also possible that the polymorphism (li’ I.-rn~(, might has no biological effect except as a
marker in linkage disequilibrium with an as yet un- known susceptibility gene.
Moreover, the frequency of GST Ml null geno- type in the HBsAg-positive HCC cases compared with that found for the healthy HBsAg carriers was not significantly different. The percentage of GST Ml null genotype in this control group is similar to that reported in a previous study on the Taiwanese population [I]. Since the major function of the GST Ml enzyme is to catalyze the conjugation of epox- ides of polycyclic aromatic hydrocarbons, aflatoxins and other compounds. This enzyme can serve as a valid susceptibility marker only if the exposure to environmental carcinogens is known. However, we did not have this information available for this study. Further epidemiological study designs are needed to elucidate the role of GST Ml on the development of HCC.
Acknowledgements
This study was supported by NSC grant NSC 84- 2331-B182-066 and Chang Gung Medical Research Grant CMRP360. References III 121 131 I41 151 I61 [71
Hell, D.A., Thompson, C.L., Taylor, J., Miller, CR., Perera, F., Hsieh, L.L. and Lucier, G.W. (1992) Genetic monitoring of human polymorphic cancer susceptibility genes by po- lymerase chain reaction: application to glutathione trans- fcrase m. Environ. Health Perspect., 98, 113-I 17.
Bishop, J.M. (1991) Molecular themes in oncogenesis. Cell, 64,235-248.
Champcme, M.H., Hieche, I., Latil, A., Hacene, K. and Lidereau, R. (1992) Association between restriction frag- ment length polymorphism of the I,-nryc gene and lung me- tastasis in human breast cancer. Int. J. Cancer, SO, 6-9. Chen. C.J.. Liang, K.Y., Chang, AS., Chang, Y.C., Lu, S.N.. Liaw, Y.F., Chang, W.Y., Sheen, M.C. and Lin, T.M. (1991) Effects of hepatitis H virus, alcohol drinking, ciga- rette smoking, and familial tendency on hepatocellular car- cinoma. Hepatology, 13, 398-406.
Chcn, D.S. (I 993) From hepatitis to hcpatoma: lessons from type B viral hepatitis. Science, 262, 369- 370.
Chcn. D.S., Kuo. G., Sung, J.I.., Lai, M.Y., Shcu, J.C., Chen, P.J., Yang, P.M., IIsu, H.M., Chang, M.H., Chen, C.J., Hahn, L.C., Choo, Q.L., Wang, T.11. and Houghton, M. (1990) Hepatitis C virus infection in an area hyperendemic of hepatitis 13 and chronic liver disease: the Taiwan expcri- encc. J. Infect. Dis., 162, 817-822.
176 L.-L. Hsieh et al. /Cuncer Letters 103 (1996) 171-176
glutathione transferases in chemical carcinogenesis. Crit. Rev. Biochem. Mol. Biol., 25,47-70.
[8] Comstock, K.E., Sanderson, B.J.S., Claflin, G. and Henner, W.D. (1990) GSTI gene deletion determined by polymerase chain reaction. Nucleic Acids Res., 18, 3670.
[9] Dubeau, L., Chandler, L.A., Gralow, J.R., Nichols, P.W. and Jones, P.A. (1986) Southern blot analysis of DNA extracted from formalin-fixed pathology specimen. Cancer Res., 46, 2964-2969.
[IO] Hayashi, S., Watanabe, J. and Kawajiri, K. (1992) High susceptibility to lung cancer analyzed in terms of combined genotypes of P4501AI and Mu-class glutathione S- transferase genes. Jpn. J. Cancer Res., 83,866-870.
[I I] Health and Vital Statistics: 2. Vital Statistics 1992 Republic of China. (1993) Department of Health, the Executive Yuan, Taiwan, ROC.
[12] Hernandez, L., Petropoulos, C.J., Hughes, S.H. and Luinsky, W. (199 I) DNA methylation and oncogene expression in methapyrilene-induced rat liver tumors and in treated hepa- tocytes in culture. Mol. Carcinogenesis, 4, 203-209. [ 131 Kaye, F., Battey, J.F., Nau, M.M., Brooks, B., Seifter, E.,
Degreve, J., Birrer, M.J., Sausville, E. and Minna. J. (1988) Structure and expression of the human L-rnyc gene reveal a complex pattern of alternative mRNA processing. Mol. Cell Biol., 8, 186-195.
[ 141 Lafuente, A., Pujol, F., Carretero, P., Villa, J.P. and Cuchi, A. (1993) Human glutathione S-transferase ,U (GST p) defi- ciency as a marker for the susceptibility to bladder and lar- ynx cancer among smokers. Cancer Len., 68,49954. [IS] Lu, S.H., Hsieh, L.L., Luo, F.C. and Weinstein, LB. (1988)
Amplification of the EGF receptor and c-myc genes in hu- man esophageal cancers. Int. J. Cancer, 42,502-505. [16] Mannervik, B., Awasthi, Y.C., Board, P.G., Hayes, J.D., Di
Rho, C., Ketterer, B., Listowsky, I., Morgenster, R., Mu- ramatsu, M., Pearson, W.R., Pickett, C.B., Sato, K., Wider- sten, M. and Wolf, C.F. (1992) Nomenclature for human glutathione transferase. Biochem. J., 282, 305-306.
[ 171 Raney, K.D., Meyer, D.J., Ketterer, B., Harris, T.M. and Guengetich, F.P. (1992) Glutathione conjugation of afla- toxin Bt exe- and endo-epoxides by rat and human glu- tathione S-transferases. Chem. Res. Toxicol., 5,470-478.
[18] Ryberg, D., Heimdal, K., Fossa, S.D., Borresen, A. and Haugen, A. (1993) Rare Ha-rus I alleles and predisposition to testicular cancer. Int. J. Cancer, 53,938-940.
[I91 Seidegard, J., Pero, R.W., Miller, D.G. and Beattie, E.J. (1986) Glutathione transferase in human leukocytes as a marker for the susceptibility to lung cancer. Carcinogenesis, 7,751-753.
[20] Seidegard, J., Vorachek, W.R., Pero. R.W. and Pearson, W.R. (1988) Hereditary differences in the expression of the human glutathione transferase active on fruns-stilbene oxide are due to a gene deletion. Proc. Natl. Acad. Sci. USA, 85, 7293-7297.
[21] Seidegard, J., Pero, R.W., Markowitz, M.M., Roush, G., Miller, D.G. and Beattie, E.J. (1990) Isozymes(s) of glu- tathione transferase (class mu) as a marker for the suscep- tibility to lung cancer: a follow up study. Carcinogenesis,
1 I, 33-36.
[22] Sheen, IS., Liaw, Y.F., Chu, C.M. and Pao, CC. (1992) Role of hepatitis C virus in spontaneous hepatitis B surface antigen clearance during chronic hepatitis B virus infection. J. Infect. Dis., 165, 831-834.
[23] Sugimura, H., Caporaso, N.E., Modali, R.V., Hoover, R.N., Resau, J.H., Trump, B.F., Lonergan, J.A., Krontiris, T.G., Mann, D.L., Weston, A. and Harris, C.C. (1990) Association of rare alleles of the Harvey t-as protooncogene locus with lung cancer. Cancer Res., SO, 1857-1862.
1241 Tamai, S., Sugimura, H., Caporaso, N.E.. Resau, J.H., Trump, B.F., Weston, A. and Harris, C.C. (1990) Restriction fragment length polymorphism analysis of the L-myc gene locus in a case-control study of lung cancer. Int. 3. Cancer, 46,411--115.
[25] Taylor, J.A., Bell, D.A. and Nagomey, D. (1993) L-myc proto-oncogene alleles and susceptibility to hepatocellular carcinoma. Int. J. Cancer, 54,927-930.
[26] Tsai, J.F., Jeng, J.E., Ho, M.S., Chang, W.Y., Lin, Z.Y. and Tsai, J.H. (1994) Hepatitis B and C virus infection as risk factors for hepatocellular carcinoma in Chinese: a case- control study. Int. J. Cancer, 56, 619-621.
[27] Yu, M.W. and Chen, C.J. (1992) Epidemiologic characteris- tics and risk factors of hepatocellular carcinoma. J. Natl. Publ. Health Assoc. (ROC), 1 I, 165-187.