Vol. 131: 87-96.1996 MARINE ECOLOGY PROGRESS SERIES
Mar Ecol Prog Ser Published February 8
Timing of metamorphosis and estuarine arrival
across the dispersal range of the Japanese eel
Anguilra japonica
P. W.
Cheng, W. N.
Tzeng*
Department of Zoology. College of Science, National Taiwan University, No. 1. Sec. 4, Roosevelt Rd, Taipei, Taiwan 10617. Republic of China
ABSTRACT: Elvers of the Japanese eel Anguilla japonica Temminck & Schlegel were collected from 6 estuaries in Taiwan, China and Japan. The fish were aged by counting otohth daily growth increments. The time of metamorphosis from leptocephali to glass eels was determined from the otolith increments where the width and strontium:calcium ratios changed drastically. Age upon arrival at the estuaries (mean i SDI was 154.71
+
10.69 to 182.06*
12.43 d , while age at metamorphosis was 115.8+
8-13 to 137.85+
11.28 d. Both ages showed a geographic cline that increased from south to north. The time from metamorphosis to arrival at the estuaries averaged 31.73*
7.58 to 45.02 +. 9.21 d. The lengthy duration of the leptocephalus stage and differences in the timing of metamorphosis are the principal factors in the long-distance dispersal of the eel.KEY WORDS: Otolith A g e ~ n g Metamorphos~s . Long-distance dispersal Elver . Japanese eel
INTRODUCTION
The Japanese eel Anguilla laponica Temminck & Schlegel is a temperate catadromous fish, widely dis- tributed in rivers of the northeastern Asian countries Taiwan, China, Korea and Japan (Tesch 1977). Devel- opment of the Japanese eel includes 5 stages, i.e. the leptocephalus, glass eel, elver, yellow eel and silver eel stages, similar to those of Anguilla anguilla (Bertin 1956). The Japanese eel was presumed to originate from the same panmictic population (Sang et al. 1994), and its spawning grounds were recently discovered in the North Equatorial Current west of the Mariana Islands, 15" N, 140" E (Tsukamoto 1992). The lepto- cephali drift with the North Equatorial Current from their spawning grounds. When they approach the con- tinental shelf of the Philippines, they turn northward into the Kuroshio Current conveyed by the mechanism of Ekman transport (Kimura et al. 1994). After reaching a fully grown size, leptocephali metamorphose into the glass eel stage and leave the strong Kuroshio Current
'Addressee for correspondence and reprint requests. E-mail: wntOccms.ntu.edu.tw
(Tsukamoto 1990). The glass eels become elvers upon arrival at estuaries. The elvers arrive at estuaries spread over an area of approxinlately 2000 km from south to north (Fig 1 ) . The mechanism determining the long-distance dispersal of the glass eel still remains open to discussion.
The body of a leptocephalus is shaped like a willow- leaf, suitable for long-distance dispersal by a current. Accordingly, the timing of metamorphosis seems to b e important in determining the geographic distribution of the eel. However, the collection of leptocephali is very difficult (Tanak 1975, Kajihara et al. 1988). The criterion for ascertaining the a g e of metamorphosis of a leptocephalus to a glass eel has been controversial (Tsukamoto & Umezawa 1990, Leomte-Finiger 1992), and there has been little use of otoliths in determining the early life history of the glass eel. Recent studies indicate that the a g e of the eel at metamorphosis can be determined by changes in otolith microstructures as well as by changes in the strontiun~:calcium (Sr:Ca) ratios in the daily growth increments of the otolith (Tzeng & Tsai 1992, 1994, Otake et al. 1994, Tzeng 1995). These findings suggest that it is possible to reconstruct early life-history events from otoliths. 0 Inter-Research 1996
88 Mar Ecol Prog Ser 131. 87-96, 1996
elver, were assessed according to the WN distribution of pigments on the body sur- faces (Strubberg 1913). Newly arrived elvers in coastal waters are in stage V,, and there a r e no external pigments on their body surfaces except the caudal spot. The skull spot appears at stage VB;
~ O ' N after this stage the eel abandons its purely pelagic existence. In stage VIB the Sirhm~picrrl Crrrmtcrt,lcrrt*trt eel loses its glass-like transparency (Tesch 1977). To precisely determine age
. ... upon arrival at a n estuary, elvers in the
20'N earlier stages of pigmentation were
selected for age determination.
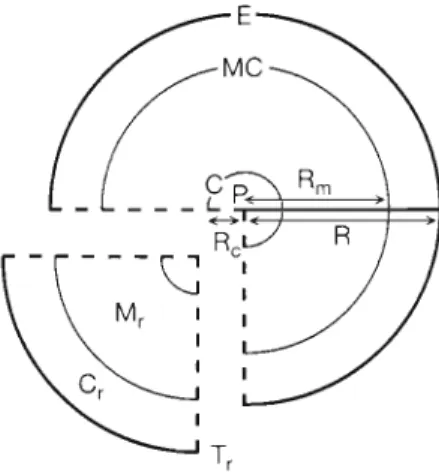
The otolith (sagitta) was extracted from the vestibular apparatus, its maximum
length was measured along the antero- IO'N posterior axis with a computer-aided Mr~rrh &c/rrnr~rr~rl Cidrren~ image analysis system (VIPRO 512), and it was then embedded in resin. The radii were measured from scanning electron 120"E 130'E 140°E 150'E
microscopy (SEM) micrographs at 300x Fig. 1. Map showing oceanic currents and 6 estuaries where elvers were magnification along the maximum axis of collected. C: Chyan-Tarng k v e r ; I: Ichinomiya River; M: Ming-Chiang the otolith sectioned on the sagittal plane River; S (S, & S,): Shuang-Shi River; T (T, & T2]: Tung-Kang River; Y. Ya- of the fish ( ~ i ~ . 2 ) . ~ ~growth i lin- ~
Lu River crements in otoliths were counted on SEM micrographs at a magnification of 2000x. Sr and Ca concentrations in the otoliths This study attempts to determine the age of Anguilla were measured from the primordium to the otolith japonica at metamorphosis from leptocephali to glass edge using an electron microprobe equipped with a
eels. These ages are used to understand the contribu- 4-channel wavelength-dispersive spectrometer (WDS; tion of the lengthy leptocephalus stage and time delay Shimadzu-ARL EMX-SM). Procedures for embedding, in metamorphosis to the long-distance dispersal of sectioning, polishing, coating and etching otoliths
Japanese eel larvae. for SEM and WDS followed those described by Tzeng
(1990) and Tzeng & Tsai (1992, 1994).
The age of an elver was determined according to MATERIALS AND METHODS the number of daily growth increments in the otolith (Tzeng 1990, Tzeng & Tsai 1992). The age a t rneta- A total of 240 elvers were collected from 6 estuaries: morphosis from a leptocephalus to a glass eel was esti- the Tung-Kang and Shuang-Shi rivers in Taiwan, the mated from the number of daily growth increments in Ming-Chiang a n d Chyan-Tarng rivers in China, the the otolith of the elver counted from the primordium to
Ichinomiya River in Japan and the Ya-Ru River the metamorphosis check, where otolith increment
between Korea and China (Fig 1 ) . The elvers were width suddenly became narrow and the Sr:Ca ratio collected using dip nets during nightime flood tides, suddenly decreased (Otake et al. 1994, Tzeng & Tsai The sampling dates are listed in Table 1 Total length 1994, Tzeng 1995). To estimate the actual age of meta-
(TL) of elvers was measured to the nearest 0.01 mm morphosis, 5 d were added to the number of the daily
after fixation in 95 % alcohol for 1 wk. Using 30 sam- growth increments, because no increment is deposited ples of various ages, the length relationship before ( Y ) in the core of the otolith during the yolk-sac stage for
a n d after (X) fixation is: 5 d (Tzeng 1990, Tzeng & Tsai 1992). The amount of time between metamorphosis and migration to the Y = -0.14
+
1.04X (r = 0.94). ( l ) estuaries was calculated by the number of daily growth This equation was used to adjust the length due to increments between the metamorphosis check and thealcohol fixation. edge of the otolith.
The pigmentation stages, which are important to Because counting daily growth increments in otoliths
C h e n g & T z e n g . metamorphosis a n d estuarine arnval of Anguilla japonlca 89
-- -p-
-Table 1 Anguilla]aponica. Total lengths a n d pigmentation stages of elvers collected from the 6 e s t u a n e s . Sampling sites shown In Flg 1
Sampllng Sampling Sample Total length ( m m ) Pigmentation stages ( % )
site d a t e s ~ z e Range M e a n + SD
v
,Av l ~ ~
v1,42 \ ' I A ~ T1 30 Dec 1992 3 0 53.54-60.49 56.96 1 1 9 95 5 T? 24 Mar 1993 3 0 51.29-60.14 56.07 i 2.4 83.3 16.7 S1 30 Dec 1992 3 0 50.99-60.85 56.84 k 2 3 1 4 5 55 S2 17 Feb 1993 3 0 51.77-59.19 55 98 t 2 15 33.3 5 6 6 10 M 1 Mar 1993 3 0 51.67-59.0 55 07 k 1 87 83 17 C 17 Feb 1993 30 5 1 16--59 23 5 5 6 k 1 8 7 70 30 Y 3 May 1993 30 55 07-60 68 5 8 3+
1.8 3.3 73 20 3 3 I 10 J a n 1994 3 0 51.31-62.17 57 4 1 t 2.3 19 4 66.7 9 7 3.2near the metamorphosis area were often diffusive and obscure, a subsample of otoliths with clear daily growth increments was selected for age-determination (Table 2) and their mean otolith growth increment widths on the corresponding radii were calculated. T h e n , the a g e at metanlorphosis (T,) was calculated from mean increment width a n d otolith radius by the following equation:
T , = M ,
+
t,, = ( R , - R,) I W R , + tyk (2) where A4, is the number of daily growth increments from the outer core to the metamorphosls check; R, a n d R, are the radii from the primordlum to the meta- morphosis check and to the outer core, respectively (Fig. 2 ) ; WRm is the mean otolith growth increment width on R,; and t,,k is the duration of the yolk-sac period before a growth increment was deposited in theFig. 2 Anguilla japonica. Schematic diagram of measure- ments of radii and counts of daily growth increments in otoliths of elvers P: p n m o r d ~ u m ; C : core; M C . metamorphosls check; E e d g e , R,, R, and R: radii from primordlum to the outer core, to the metamorphosis check a n d to t h e otolith e d g e , respectively, M,, T, a n d C, counts of daily growth in-
crements corresponding to the radli of R,,, a n d R and the section between M C a n d E , respectively
core of the otholith (approximately 5 d ) . Similarly, the amount of time between metamorphosis and m ~ g r a t i o n to the e s t u a r ~ e s , i.e. the number of daily growth incre- ments between the metamorphosis check a n d the e d g e of the otolith (C,), was estimated by the equation:
where R is the radius from the primordium to the e d g e of the otolith and W I R Y , , , is the mean increment width on the otolith from the metamorphosis check ( M C ) to the e d g e ( E ) (Fig. 2 ) . Then, the a g e of a n elver upon arrival at a n estuary (T,) equals T , plus
Cr.
The hon~ogeneity of T,, T,, and C, among estuaries was tested with Scheffe's multiple range analysis (Sokal & Rohlf 1981). Correlation coefficients were cal- culated among fish lengths, pigmentation stages, otolith maximum lengths, otolith radii at metamorpho- sis, a g e at metamorphosis, a g e upon arrival a t the estu- aries, a n d the amount of time needed for migration to the estuaries after metamorphosis.
Table 2. Angullla laponica Counts of daily growth incre- ments ( m e a n t SD) in otollths of elvers collected from the 6 estuaries (T, S , M , C, Y and I ) shown In Fig. 1. M,, C, a n d T, counts of daily growth increments corresponding to the radil of R,, R-R,, a n d R as shown In Flg 2 Sample slzes
In parentheses
Sampling Otolith growth increment
site M , C, Tr - p T , 144 8 1 1 1 1 (7) 4 1 . 6 t 9 0 (7) 186.4
+
12.5 (7) T2 1 3 3 . 9 k 8.0 (7) 39.4+
2 4 (5) 172 0 & 5.9 (5) S , 147.1 1 15.7 (7) 4 2 . 2 t 6 0 (5) 1 8 9 . 0 k 16.4 (5) S, 134.3+ 17 7 (4) 3 5 . 0 5 5 7 (3) 175 3 + 11.1 (3) M 141 5+
9.8 (7) 34 2 1 6 8 (6) 1 7 7 . 6 + 12.6 (6) C 151 7 ? 10 6 ( 1 2 ) 4 4 . 3 k 8.0 (9) 198 3 k 6 9 ( 9 ) Y 1 5 9 . 7 2 8.3 (11) 41.8+ 9.0 (11) 201.5+ 11.9 (11) I 1 4 7 6 k 7 . 2 (8) 4 0 . 6 + 1 0 5 ( 7 ) 1 8 9 7 f 9 . 2 (7)Cheng & Tzeng: Metamorphosis and estuarine arrival of Anguilla japonica 91
Fig. 3. Angujlla japonica. S E M micrograph illustrating changes In ( a ) phase contrast a n d ( b ) incremental patterns In otoliths of elvers during n~etamorphosis. ( a ) 55.1 mm TL collected from the Tung-Kang River estuary on 24 February 1993; ( b ) 56.68 mm TL
from the same estuary on 30 December 1992. MC: metamorphosis check
RESULTS
Size a n d pigmentation stage of elvers
Total lengths of elvers arriving at the G estuaries ranged from 50.99 to 62.17 mm, with averages of 55.07 k 1.87 to 58.3 ? 1.8 mm (Table 1 ) . The elvers from the Ya-Lu River estuary in northern China were sig- nificantly longer than those from other estuaries (t-test, p c 0.01), but those from other estuaries showed no
significant difference (t-test, p > 0.05). Most newly recruited elvers in the estuaries were in the pigmenta- tion stages V, a n d V,. The occurrence of larger elvers in the Ya-Lu River estuary corresponded with their more advanced stages of pigmentation (VIA, a n d VIA?) (Table 1). There was a significant correlation between TL and the pigmentation stage of elvers (r = 0.32, df = 139, p < 0.0001).
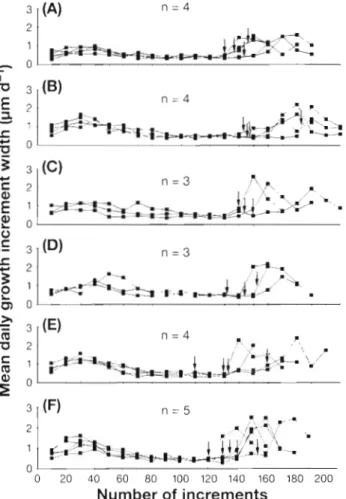
Otolith microstructure a n d a g e determination The microstructure of the otoliths of elvers showed obvious changes in phase contrast and crystal patterns (Fig. 3 ) . T h e brightness of the SEM otolith micrograph became darker beyond the MC (Fig. 3a), a n d the incre- mental pattern changed from a circular form to a radial form beyond the check (Fig. 3b). The increment widths gradually decreased beginning at approximately the 30th daily growth increment from the primordium and became smallest at the presumed MC, between approximately the 120th a n d the 150th daily growth increments from the primordium; beyond the check the increment widths drastically increased (Fig. 4 ) . These morphological changes corresponded with the drastic change in Sr contents or Sr:Ca ratios in the otolith a s described in a previous study (Tzeng & Tsai 1994), which associated the change with metamorpho- sis from a leptocephalus to a glass eel (Otake e t al. 1994). The leptocephalus is known to contain numer- ous gelatinous extracellular matrixes composed of sul- fated glycosaminoglycans (GAG), which a r e broken down during the process of metamorphosis (Pfeiler 1984, 1986, 1991). GAG exhibits an affinity to alkali earth elements and, in particular, to Sr (Nishizawa 1978). GAG breakdown may reduce the absorption of Sr and result in a drastic decrease in otolith Sr contents a n d , consequently, in Sr:Ca ratios. Thus, the number of daily growth increments in the otolith, the timing of the drastic changes in the otolith microstructural pattern,
a n d changes in Sr:Ca ratios in t h e otolith w e r e used to estimate the a g e of metamorphosis from leptocephali to glass eels. Fig. 5 shows daily growth increments in the otolith of a 56.5 mm TL elver collected from the Chyan-Tarng River estuary on 27 Febuary 1993. The flsh metamorphosed a t approximately 158 d after hatching and took approximately 33 d to migrate to the estuary after the metamorphosis. The elver arrived a t the estuary at approximately 191 d after hatching.
M e a n ( 5 SD) daily growth increments in otoliths from
a subsample of elvers from the 6 estuaries a r e shown in Table 2. Actual a g e a t metamorphosis a n d upon arrival a t the estuaries w a s based on the counts of daily growth increments plus t h e 5 d yolk-sac period.
0 1
0 20 40 60 80 100 120 140 160 180 200
Number of increments
Fig. 4 . Angullla japonica. Temporal changes in mean daily growth increment widths of otoliths of elvers collected from 6 estuaries: ( A ) Ichinom~ya K ~ v e r ; (B) Ya-Lu River; ( C ) Chyan- Tarng River; ( D ) Ming-Chiang R ~ v e r , (E) Shuang-Shi River; (F) Tung-Kang River. Arrows: l o c a t ~ o n s of metamorphosis
92 Mar Ecol Prog Ser 131: 87-96,1996
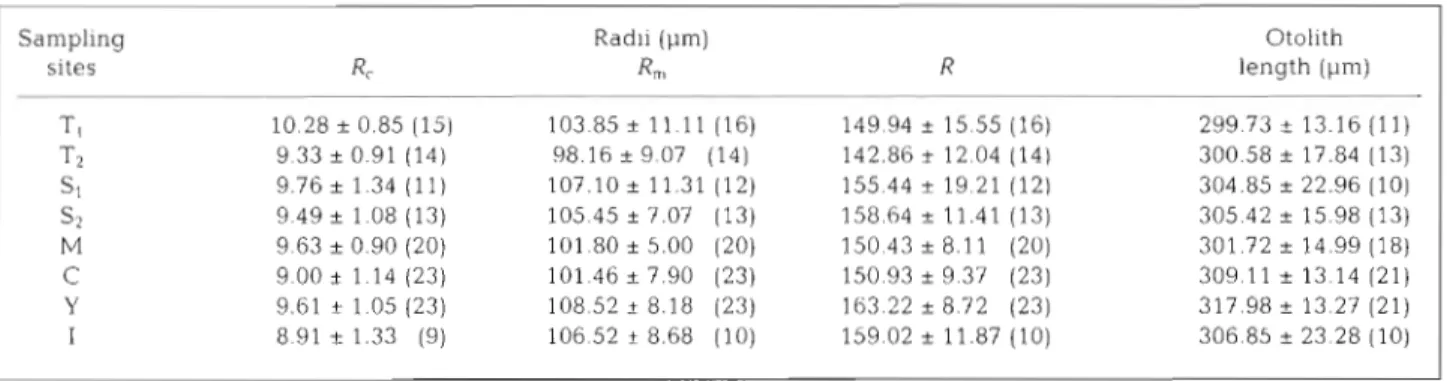
Otolith radius and increment width Otolith radii of elvers at different phases of their early life history (Fig. 2) are shown in Table 3. The radii were 8.91
+
1.33 pm (Sampling Site I in Japan) to 10.28+
0.85 pm (Site T1 in Taiwan) for the cores (R,), 98.16+
9.07 pm (Site T2 in Taiwan) to 108.52+
8.18 pm (Site Y in China) for the metamor- phosis checks (R,), and 142.86+
12.04 pm (Site T,) to 163.22*
8.72 pm (Site Y) for the maximum radii (R). The otolith maximum lengths were 299.73+
13.16 pm (Site T,) to 317.98+
13.27 pm (Site Y). The otoliths were larger for fish captured in estuaries farther from the spawning grounds, but the cores were smaller.Mean otolith growth increment widths of elvers at different phases of their early life history are shown in Table 4. Before meta- morphosis the mean otolith growth incre- ment widths showed a geographic cline, decreasing from south to north (Table 4B), but after metamorphosis the mean increment widths were homogenous among the 6 estu- aries (Table 4C).
The number of daily growth increments from the outer core to the metamorphosis check (Y) was negatively correlated with the mean increment width (X) ( Y = 224.759 - 110.77X; r = -0.71, n = 37). This indicates that fast-growing leptocephali metamorphosed earlier.
Geographic cline in age a t metamorphosis and arrival at estuaries
Age upon arrival at the estuaries differed among the 6 estuaries with the shortest period being 154.71 c 10.69 d in Taiwan (Site
SZ)
and the longest perlod 182.06 ? 12.43 d in Japan (Si.te I ) , a n average differ- ence of 21.35 d . Age at metamorphosis was 115.80+
8.13 to 137.85 k 11.28 d , a difference of 22 d (Table 5). The eel had a lengthy leptocephalus stage during oceanic passive migration, and there was a geographic clineFig. 5 . Angdlla japon~ca. Daily growth increments in otolith of a
56.5 mm TL elver collected from the Chyan-Tang River estuary on 27 February 1993 P. primordium; MC: metamorphosis check.
Cheng & Tzeng: hlletanlorphosis and estuarine al-rival of Anguilla j ~ ~ p o n i c a 93
Table 3. Anguilla japonica. Otolith length and radii (mean i SD) of elvers collected from the 6 estuaries (T, S, M , C , Y and I ) shown in Fig. 1. R,, R,, and R are defined in Fig. 2. Sample sizes in parentheses
Sampling sites - T , 7-2 S1 S2 M C Y Radli (pm) R", - Otolith length (pm)
in a g e at metamorphosis that increased from south to DISCUSSION north (Fig. 6B). The duration of the migration from the
time of metamorphosis to the time of arrival at the estu- The age of metamorphosis from a leptocephalus to a aries was 31.73
+ 7.58
to 45.02 + 9.21 d (Table 5). This glass eel for the Japanese eel was determined by duration also showed a geographic cline that increased Tsukamoto & Umezawa (1990) to be on the average from south to north except at Sites T, and T2 in south- between 85 and 97 d , which is less than the 116 to ern Taiwan (Fig. 6 C ) . 138 d estimated by our study (Table 5 ) . This discrep-Table 4 . Angullla japonica Homogeneity test for increment widths (mean
+
SDI on the radli of (A) R, ( B ) R, and ( C ) R-R,, in otoliths of elvers collectcd from the 6 estuaries. R, R, and R-R, are defined in Flg 2. The 6 estuaries (T, S, M , C, Y and I ) are shown in Fig. 1 Asterisks shared between samplingsites indicate they are the same homogeneous group
Table 5. Angujlla japonica. Homogeneity test for (A) age of elvers arriving at the 6 estuar~es, (B) age of leptocephali at metamorphosis, and ( C ) time between metamorphosis and arrival at the estuaries. The 6 estuaries (T, S, M. C, Y and I )
are shown in Flg. 1. Asterisks shared between sampling sites indicate they are the same homogeneous group Sampling site (A) R T2 C I Y M T I S1 S2 (8) R, C I M Y T2 S i T I S2 ( C ) R-R, T2 I 1 J C S2 M S1 Sample size - - - -5 4 5 8 5 5 6 2 7 5 5 8 7 7 5 3 5 5 5 8 4 2 5 6 Increment width (pm) Homogeneous group
Sampling Sample Age Homogeneous
site size (d) group
(A) Age of elvers
S2 13 154 71 T 1 0 6 9 T1 16 156.86 i 17 44 SI 12 157.61 i 18 78 M 20 162 87 -c 8.59 T2 14 1 6 4 2 9 t 1 4 1 7 C 23 176 49 t 11.34 Y 2 3 178 35 T 9.94 I 10 182 06
*
12 43 ( B ) Age of leptocephali S2 13 115.80 ? 8-13 T. 16 117.70 + 14.26 T2 14 121.39 12.04 5 1 1 'L 125.88+
14.7 M 20 128.38 -r 6 86 Y 23 135.52+
11.28 1 10 137.04 _c 12.89 C 23 137.85 t 11.28 ( C ) Timo from metamorphosis to estuariesS , 12 31 73
+
7.58 h4 2 0 34.49+
3.62 C 23 38.64+
5.72 S? 13 38.91 + 5.82 T I 16 39.16+
6 83 Y 23 42 83 + 7 36 T2 14 42.90+
6.20 l 10 45.02+
9.2194 M a r Ecol Prog Ser 131: 87-96, 1996
Fig. 6 Anguilla japonica. Geographic cline in ( A ] a g e of elvers arnvlng at 6 estuaries, ( B ] a g e of leptocephali at meta- morphosis, a n d (C) time between metamorphosis a n d arri\.dl at estuaries. T h e 6 estuaries (T, S, M. C , Y a n d I ) are sh.own
in Fig. 1
ancy is probably due to the different methods used to count the otolith daily growth increments around the metamorphosis zone. Tsukamoto & Umezawa (1990) indicated that otolith, increment widths became larger during the period when leptocephali metam.orphosed to glass eels. We found that increment widths became narrower in the area close to the metamorphosis check (Fig. 4 ) , where fine daily growth increments were pre- sent (Fig. 5). Therefore, we believe that during the metamorphosing period otolith growth decreases and increment width becomes narrow, and, thus, we counted more increments.
The duration of the leptocephalus stage was 116 to 138 d. A prolonged larval stage has also been fou.nd In other species, for example, 110 to 170 d for I n g George whiting Sillaginous punctata (Jenkins & hldy 1994). Variation in the duration of the larval stage is a n important factor in determining the distribution of a fish. In the absence of a suitable habitat for settlement,
larvae may delay metamorphosis (Jackson & Strath- mann 1981, Victor 1986), a delay which may last for months in several species of invertebrates (Scheltema 1971, Hadfield 1978, Domanski 1984, Pechenik et al. 1984, Richmond 1985). This flexibility in time of settle- ment and metamorphosis allows the larvae enough time to disperse over great distances in order to find a suitable habitat (Scheltema & Williams 1983, Christo- pher & Leggett 1987) Interspecific ~~ariations in the length of the larval pc,riod and tho time at which meta- morphosis takes place depends on the early life history strategy of a species and on the hydrodynamic pro- cesses operating at the time of the dispersal (Jenkins & May 1994).
The timing of the metamorphosis of a leptocephalus and transportation by the Kuroshio Current were con- sidered to be important factors determining the ulti- mate destination of eels (Tsukamoto & Umezawa 1994). The velocity of the Kuroshio Current averages 2.5 knots (range 2 to 3 knots), i.e. 96 km d-' (77 to 116 km d-l) (Nitani 1972). The difference in mean ages of leptocephali at metamorphosis to glass eels esti- mated from the elvers collected at the 6 estuaries in this study was approximately 22 d (Table 5 ) . This time lag allows the Kuroshio Current to disperse the lepto- cephali to a distance of about 2100 km at a velocity of 2.5 knots. This distance was very close to the estimated distance of approximately 2000 km between the north- ernmost and southernmost limits of the 6 estuaries studied (Fig. 1). This evidence suggests that the timing of the metamorphosis of the leptocephali and the Kuroshio Current play important roles in the long- distance dispersal of the Japanese eel.
The mean increment widths of the otoliths from the outer core to the metamorphosis check for the elvers collected at the 6 estuaries showed a geographic cline that decreased from south to north, but age at meta- morphosis increased from south to north. These oppo- site trends in the geographic clines of otolith growth rate and age a t metamorphosis indicated that the fast- growing leptocephall m.etamorphosed earlier and migrated to the southern estuaries of the distribution range, while the slow-growing ones metamorphosed later and migrated to the northern estuaries of the range. Apparently, a g e at metamorphosis is a n impor- tant factor in determining the southern limit of the dis- tribution of the Japanese eel.
The Japanese eel is seldom found in Philippine waters (Tabeta et al. 1975, Tabeta et al. 2976), although the larvae migrate in that direction. The eel spawn in waters west of the Mariana Islands (1.5'
N,
140" E), and their larvae drift with the North Equatorial Current at a speed of 20 to 32 cm S-' (Kimura et al. 1994). Depending upon the speed of the current a n d the distance involved, larvae arrive in PhilippineChcsng & Tzeng: \letamorphosis a n d e s t u a r ~ n e arrival of Anguilla japonica 9 5
waters approxin~ately 55 to 87 d after hatching. The a g e of the elvers at ~netamorphosis a t the 6 estuaries studied in Taiwan, China a n d J a p a n averaged 116 to 138 d (Table 5). T h e leptocephali which arrive in Philippine waters a r e apparently too young to meta- morphose and migrate toward estuaries. This may explain why the Japanese eel is seldom found in the Philippines.
Three fully groct7n leptocephali of Japanese eel were collected in the southeastern waters of Taiwan, be- tween approximately 20" and 22" N . O n e of them had a TL of 55.80 a n d was 84 d old (Tanaka 1975, Tabeta & Takai 1975a,b, Tsukamoto e t al. 1992). Based on the a g e at metamorphosis (116 to 138 d ) estimated in this study a n d the location of the fully grown leptocephali, the latitude where leptocephali may metamorphose to become glass eels was estimated to b e in a n area further north from Taiwan. If this is true, the migratory distance covered from the time of metamorphosis to the tlme of arrival at estuaries should b e greater for elvers in the southern portion of Taiwan than for those in the northern portion (Fig. 1). This may explain why the time from metamorphosis to arrival a t estuaries for the elvers collected at Site T (T, &
T,)
in southern Tai- wan was longer than for those collected at Site S(S,
&S2) in northern Taiwan (Fig. 6 C ) . This also supports the hypothesis that elvers along the southwestern coasts of Taiwan a r e being carried by the China Coastal Current from the north (Tzeng 1985, 1986, Tzeng & Tsai 1992).
In conclusion, a g e at metan~orphosis a n d the lengthy duration of the leptocephalus stage a r e important factors determining the long-distance dispersal of the Japanese eel.
Acknowledgements. This study w a s financially supported by the National Science Council, Republic of China (Project No. NSC 852311B002032). T h e authors a r e grateful to Mr Y. H. Shieh and Dr K . Tsukamoto for providing the fish specimens from Chlna a n d J a p a n , to Miss C . H. Wang, Miss C . E Wu and Mr H. P Wei for pi-epanng the manuscript, and to Dr T F. Tsai, Dr K. Severin a n d 3 anonymous reviewers for helpful comments.
LITERATURE CITED
Bertin L (1956) Eels - a biolog~cal study. Cleaver-Hume Press, London
Christopher RC, Leggett WC (1987) Size a n d a g e at metamor- phosis in marine flshes: a n a n a l y s ~ s of laboratory-reared w ~ n t e r flounder (Pseudopleuronectes an1erlcarus) with a review of varlatlon in other species. C a n J Fish Aquat SCI 44:1936-1947
Domanski PA (1984) Giant larvae: prolonged planktonic lar- val phase in the asteroid Luldia sarsi. Mar Biol 80~189-195 Hadfield LIG (1978) Growth and metamorphosis of plank-
tonic larvae of Ptychodera flava (Hemichordata Enterop-
neusta). In: Chit1 FS, Rice ME (eds) Settlement and meta- morphosis of marlne invertebrate larvae. Elsevier, N C ~ Y York, p 247-254
Jackson GA, Strathmann RR (1981) Larval mortality from off- shore mixing a s a link betlveen precompetent and compe- tent periods of developn~ent. Am Nat 118:16-26
J e n k ~ n s GP, May HMA (1994) Variation in settlement a n d larval duratlon of King George whiting, Slllaginocles p u n c - tata (.ailldginldae), in Swan Bay. Victoria, Australia. Bull mar Sci 54(1):281-296
Kajihiira T, Tsukamoto K , O t a k e T. 'Lloritaka K, Hasumoto H, Oya hl, Tabeta 0 (1988) Sampling Ieptocephal~ with reference to the die1 vertical migrahon and the gears. N ~ p p o n Suisan Gakkaishi 54(6):94 1-946
Kimura S, Tsukamoto K, Sugimoto T (1994) A model for the larval migration of the J a p a n e s e eel: roles of the trade winds and s a l i n ~ t y front. Mar Biol 119:185-190
Lecomte-Finiger R (1992) Growth hlstory a n d a g e at recruit- ment of European glass eels (Anguilla anguilla) as revealed by otolith microstructure. Mar Biol 114.205-210 Nishizawa K (1978) Marine algae from a v ~ e w p o i n t of phar-
maceutical studies. J a p J Phycol 26:73-78
Nitani H (1972) Beginning of the Kuroshio. In: Stommel H, Yoshida K (eds) Kuroshio - its physical aspects. Univ Tokyo Press, p 129-163
Otake T, Ishii T, Nakahara M, Nakamura R (1994) Drastic changes In otolith strontium/calcium ratios in leptocephali a n d glass eels of J a p a n e s e eel Anguilla japonica. ]Mar Ecol Prog Ser 112:189-193
Pechenik JA, Scheltema RS, Eyeter LS (1984) Growth stasis a n d limitc~d shell calcification in larvae of Cymatium p a r t h e n o p e u n ~ during trans-Atlantic transport. Sclence 224: 1097-1099
Pfeiler E (1984) Glycosaminoglycan breakdown during meta- morphosis of larval bone fish Albula. Mar Biol Lett 5: 241-249
Pfeiler E (1986) Towards an explanation of the developmental strategy in leptocephalus larvae of marine teleost fishes Environ Biol Fish 15:3-13
Pfeiler E (1991) Glycosaminoglycan composition of anguilli- form a n d elopiform leptocephali. J Fish Biol38:533-540 Richmond RH (1985) Revers~ble metamorphosis in coral
planula larvae. Mar Ecol Prog Ser 22:181-185
Sang TK, Chang HY, Chen CT, Hui C F (1994) Population structure of the J a p a n e s e eel, Anguilla japonica. Mol Biol Evol 11(2):250-260
Sheltema RS (1971) Larval dispersal a s a means of genetic exchdnge between geographically separated populations of shallo~v-water benthic marine gastropods Biol Bull 140 284-322
Scheltema RS, \rVilllams IP (1983) Long-distance dispersal of planktonic larvae a n d the biogeography a n d rvolution of some Polynesian a n d Western Paclfic mollusks. Bull mar Sci 33:545-565
Sokal RR, Rohlf FJ (1981) B~onietry, 2nd e d n WH Freeman, San Francisco
Strubberg AC (1913) T h e metamorphosis of elvers a s influ- enced by outward conditions. Meddr Kommn Danm Flsk- l-lavunders 4:l-11
Tabeta 0, Takzil T (1975aj Leptocephalus of Anguilla japon- ica found in the waters south of Taiwan. J a p J lchthyol 22:lOO-103
Tabeta 0 , Takai T (1975b) Leptocephali of Anguilla japonica found in the waters south of the Okinawa Island. Nippon Suisan Gakkaishi 4 1: 137- 145
Tabeta 0 , Takai T, Matsun I 1 1975) Record of the Japanese eel from the Phlllppines. Nippon Suisan Gakkaishi 4 1:64 1-644
Mar Ecol Prog Ser 131: 87-96, 1996
Tabeta 0 , Tanimoto T, Tanaka T. Matsui I, lmamura T (1976) Seasonal occurrence of angulllid elvers In Cagayan River, Luzon Island, the Philippines. Nippon Suisan Gakkaishi 42:421-426
Tanaka S (1975) Collection of leptocephali of the Japanese eel in waters south of the Okina\ra Island. Nippon Suisan Gakkaishi 41:129-136
Tesch FW (1977) The eel - biology and management of anguillid eels. Chapman & Hall, London
Tsukamoto K (1990) Recruitment mechanism of the eel Anguilla japonica to the Japanese coast. J F ~ s h Biol 36: 659-671
Tsukamoto K (1992) Discovery of the spawning area for Japanese eel. Nature 356:789-791
Tsukamoto K, Umezawa A (1990) Early life history and oceanic migration of the eel Anguilla japonica. La Mer 28 188-198
Tsukamoto K , Umezawa A (1994) Metamorphosis: a key factor of larval migration determining geographic distri- bution and speciation of eels. In: Proceedings of 4th Indo- Pacific fish conference. Bangkok, Thailand, 28 Nov-4 Dec 1993. Faculty of Fisheries, Kasetart University, p 231-248 Tsukamoto Y , Umezawa A, Tsukamoto K, Okiyama M (1992)
A fully grown leptocephalus of Japanese eel collected from the western north Pacific. Nippon Suisan Gakkaishi 58.2209
This a r t ~ c l e was submitted to the editor
Tzeng WN (1985) Imm~gration timing and activity rhythms of the eel, AngsliUn japonica, elvers in the estuary of north- ern Taiwan, with emphasis on environmental influences Bull Jap Soc Frsh Oceanogr 47/48:11-28
Tzeng WN (1986) Resources and ecology of the Japanese eel Angujlla japonica elvers in the coastal waters of Taiwan. China Fisheries Monthly 404:19-24 (in Chinese)
Tzeng WN (1990) Relationship between growth rate and age at recruitment of Anguilla japonica elvers in a Taiwan estuary as inferred from otolith growth increments Mar Biol 107:75-81
Tzeng WN (19951 Migratory history recorded in otollths of the Japanese eel, Angnilla laponica, elvers as revealed from SEM and WDS analysis. Zoo1 Stud 34 (Suppl) 1: 234-236
Tzeng WN, Tsai Y C (1992) Otolith microstructure and daily age of Angujlla japonica Temminck & Schlegel elvers from the estuaries of Taiwan with reference to unit stock and larval migration. J Fish Biol 40:845-857
Tzeng WN, Tsai YC (1994) Changes in otolith microchemistry of the Japanese eel, Anguilla japonica, during its migra- tion from the ocean to the rivers of Taiwan. J Fish Biol 45: 67 1-684
Victor BC (1986) Delayed metamorphosis with reduced larval growth in a coral reef fish (Thalassoma bifasciatum) Can J Fish Aquat Sci 43:1208-1213
Manuscript first received: May 26, 1995 Revised version accepted: Septem ber 26, 1995

![Fig. 6 Anguilla japonica. Geographic cline in ( A ] a g e of elvers arnvlng at 6 estuaries, ( B ] a g e of leptocephali at meta- morphosis, a n d (C) time between metamorphosis a n d arri\.dl at estuaries](https://thumb-ap.123doks.com/thumbv2/9libinfo/8851246.242266/8.873.97.445.116.633/anguilla-japonica-geographic-estuaries-leptocephali-morphosis-metamorphosis-estuaries.webp)
