The Migratory Environmental History of Freshwater Resident Flathead
Mullet Mugil cephalus L. in the Tanshui River, Northern Taiwan
Chia-Hui Wang1, Chih-Chieh Hsu1,2, Chih-Wei Chang3,4, Chen-Feng You1, and Wann-Nian Tzeng2,* 1Earth Dynamic System Research Center, National Cheng Kung University, 1 University Rd., Tainan 701, Taiwan
2Institute of Fisheries Science, National Taiwan University, 1 Roosevelt Rd., Sec. 4, Taipei 106, Taiwan 3National Museum of Marine Biology and Aquarium, 2 Houwan Rd., Checheng, Pintung 944, Taiwan
4Institute of Marine Biodiversity and Evolution, National Donghwa University, 1 Da-Hsueh Rd., Sec. 2, Shoufeng, Hualien 974, Taiwan
(Accepted December 3, 2009)
Chia-Hui Wang, Chih-Chieh Hsu, Chih-Wei Chang, Chen-Feng You, and Wann-Nian Tzeng (2010) The
migratory environmental history of freshwater resident flathead mullet Mugil cephalus L. in the Tanshui River, northern Taiwan. Zoological Studies 49(4): 504-514. The diversity of the migratory environmental history of
freshwater resident flathead mullet Mugil cephalus was examined by analyzing the otolith elemental composition of mullet (n = 31) collected in midstream of the Tanshui River, northern Taiwan by laser-ablation inductively coupled plasma mass spectrometry. The mullet averaged 300 ± 33 mm in fork length and 410 ± 141 g in body weight, and all were sexually immature at 1+ and 2+ yr old. Strontium (Sr)/calcium (Ca) and barium (Ba)/Ca ratios in the otolith core indicated that the mullet in the river were from 2 different groups. Group 1 (18%) fish had higher Sr/Ca ratios and lower Ba/Ca ratios in the otolith core region. Group 2 fish (82%) were opposite to group 1, with higher Sr/Ca ratios and lower Ba/Ca ratios in the otolith core region, indicating that these mullet had resided in offshore waters in early life. Beyond the core region, the Sr/Ca and Ba/Ca ratios alternately varied, indicating that the mullet migrated between brackish and freshwater habitats after the juvenile stage. This study demonstrates that M. cephalus occupied diverse environments and migrated among marine, brackish, and freshwater habitats. More study is needed to determine whether the mullet in the Tanshui River of Taiwan is a local population and its relationship to the well-known offshore migratory population that migrates to southwestern Taiwan to spawn in Dec. http://zoolstud.sinica.edu.tw/Journals/49.4/504.pdf
Key words: Mullet, Freshwater fish, Otolith elemental composition, Migratory environmental history.
*To whom correspondence and reprint requests should be addressed. Chia-Hui Wang and Chih-Chieh Hsu contribute equally to this work. Tel and Fax: 886-2-23639570. E-mail:[email protected]
T
he flathead mullet (Mugil cephalus L.) is a cosmopolitan euryhaline fish, distributed in coastal, lagoon, and estuarine waters between 42°N and 42°S latitudes (Thomson 1966). It is economically important worldwide for both capture fishery and aquaculture. More attention has been focused on wild populations of mullet due to significant decrease since 1980 because of overharvesting for the roe (Chang and Tzeng 2006). The population of flathead mullet in waters adjacent to Taiwan migrates annually from the feeding grounds in coastal waters of China to offshore waters of both southwestern and northeastern Taiwan tospawn in Dec. at the age of 3-4 yr (Chen and Su 1986, Huang and Su 1989, Chang et al. 2004a, Hsu et al. 2007). After a process of passive dispersal for approximately 1 mo after spawning, they metamorphose from larvae to juveniles and are actively recruited to the estuarine nursery area (Blaber 1987, Chang et al. 2000, Hsu et al. 2009). Whether juveniles enter freshwater environments is controversial. Some hypothesized that all juveniles and post-spawning adults might migrate northward to coastal waters of the Chinese mainland to feed until the spawning migration (Tung 1981), while some proposed that a few juveniles
resided in coastal waters around Taiwan without a northward migration (Liu 1986). Immature mullet of various sizes are found year round in coastal waters, estuaries, and bays of Taiwan (Chen et al. 1989, Lee 1992, Huang et al. 2001). However, the detailed migrations and habitat movements of freshwater resident flathead mullet have never been investigated, nor has information on their connections and contributions to the known offshore migratory population, that is important for sustainable fisheries management for flathead mullet, been established.
Several tagging and marking techniques, such as satellite-transmitting archival tags, coded wire tags, chemical markers, radio telemetry, acoustic tags, and otolith elemental signatures can be used to study the migration history and population structure of fishes (Campana et al. 2000, Loher 2008). Otolith elemental signatures serve as a sensitive natural tag and can be used to track the entire life history of a fish. The otolith is located in the inner ear and functions for hearing and balance of the fish. It is a biomineralized aragonite crystalline structure mainly composed of CaCO3 with a minor organic substrate. Also,
a number of elemental impurities, e.g., sodium (Na), strontium (Sr), potassium (K), zinc (Zn), magnesium (Mg), manganese (Mn), lead (Pb), barium (Ba) etc., are co-precipitated in the otolith growth increments as the fish grows (Campana 1999). Elemental deposition in the otolith is influenced by both physiological and environmental factors, particularly the concentration of elements in the ambient water. The chemicals deposited represent a permanent record of the environmental conditions experienced by the fish at a particular time (Ruttenberg et al. 2005). In addition, otolith growth increments are periodically deposited in daily and annular rhythms which allow one to determinate the daily and annual age of a fish (Pannella 1971). Mullet otolith daily increment and annular periodity deposition rhythms were validated (Chang et al. 2000, Hsu and Tzeng 2009). Accordingly, the daily and annual age determination of mullet, together with the measurement of trace elements in the increment, can be used as a biological tracer to reconstruct the migratory environmental history and habitat use of the fish (Campana et al. 2000). Alkaline earth metals, such as Sr and Ba, have similar ionic radii as Ca, and may substitute for Ca in the orthorhombic aragonite lattice of fish otoliths (Campana 1999), so their concentrations are higher in otoliths compared to other trace metals.
Also, concentrations of Sr and Ba vary between fresh water and seawater, and are useful as natural indicators for understanding diadromous fish migration behavior. The Sr concentration is approximately 100 times higher in seawater than in fresh water (Campana 1999). Rearing experiments showed that otolith Sr/Ca ratios of M.
cephalus are positively correlated with the salinity
of the ambient water (Chang et al. 2004b). The Ba/Ca ratio in otoliths is another candidate which can be used to reconstruct the history of fish movements (Elsdon and Gillanders 2004 2005). Barium is sourced terrestrially in rock, soil, and aquatic suspended particles, and the dissolved Ba concentration is higher in river water than in seawater (Edmond et al. 1985, Chester 2000). Thus, elemental ratios such as Sr/Ca and Ba/Ca in otoliths, along with the fish age, are widely used to reconstruct the past migratory environmental history of fish (Fowler et al. 2005, Morales-Nin et al. 2005, Bradbury et al. 2008).
This study used the otolith elemental composition as a natural tag to study the migratory environmental history of the freshwater resident mullet M. cephalus in the Tanshui River, northern Taiwan. The otolith elemental composition was measured by laser-ablation inductively coupled plasma mass spectrometry (ICP-MS). Temporal changes in the elemental composition in otoliths permit reconstruction of the migratory environmental history of freshwater resident mullet and examination of the diversity of mullet migratory patterns in northern Taiwan.
MATERIAL AND METHODS Sample collection
In total, 31 M. cephalus individuals were collected from midstream of the Tanshui River, the largest river in northern Taiwan, on 3rd-5th Oct. 2006 (Fig. 1). The sampling site is near the Fu-Ho Bridge, close to the conjunction of the Tanshui River and one of its major tributaries, the Shindien River, and is a freshwater environment (0 psu). After collection, the fish were immediately preserved on ice, and their fork length (FL, mm), body weight (BW, g), and gonad weight (GW, g) were measured fresh within 24 h (Table 1). Sexes of the fish were determined from the external features of the gonads (Hsu et al. 2007). The maturation stage of each fish was determined by the gonadosomatic index (GSI) which was
Fig. 1. Sampling location of Mugil cephalus (marked with “+”) midstream in the Tanshui River, northern Taiwan. 25°N 25°N 120°E 120°E (A) 5 km (B)
Table 1. Sampling date, sample size (F, female; M, male; -, undetermined), age (yr), mean (± SD) fork
length (FL, mm), body weight (BW, g), and gonadosomatic index (GSI, %) of Mugil cephalus collected from the Tanshui River, northern Taiwan. Otoliths that underwent LA-ICPMS analyses are marked with a V
No. Date FL BW GSI Sex Age LA-ICPMS
1 3rd Oct. 2006 380 833 3.09 M 2+ V 2 3rd Oct. 2006 312 439 2.81 M 1+ V 3 3rd Oct. 2006 303 452 0.67 F 2+ V 4 3rd Oct. 2006 262 281 - - 2+ -5 3rd Oct. 2006 237 189 - - 2+ V 6 3rd Oct. 2006 283 350 3.55 F 2+ V 7 3rd Oct. 2006 268 275 - - 1+ -8 4th Oct. 2006 371 734 2.57 F 2+ V 9 4th Oct. 2006 307 436 0.40 M 2+ V 10 4th Oct. 2006 322 470 2.05 F 1+ V 11 4th Oct. 2006 277 331 - - 1+ V 12 4th Oct. 2006 290 398 0.43 M 1+ V 13 4th Oct. 2006 283 325 0.17 F 1+ V 14 4th Oct. 2006 318 487 0.83 F 1+ V 15 4th Oct. 2006 294 356 0.11 F 1+ V 16 4th Oct. 2006 280 339 0.68 M 1+ V 17 4th Oct. 2006 281 344 0.25 F 1+ V 18 4th Oct. 2006 294 355 0.10 M 1+ -19 4th Oct. 2006 266 280 - - 1+ -20 5th Oct. 2006 287 322 - - 1+ V 21 5th Oct. 2006 356 541 1.31 F 2+ V 22 5th Oct. 2006 316 457 - - 1+ V 23 5th Oct. 2006 342 629 0.58 F 2+ V 24 5th Oct. 2006 297 365 2.44 M 1+ -25 5th Oct. 2006 353 692 0.69 F 2+ V 26 5th Oct. 2006 295 390 0.18 F 1+ V 27 5th Oct. 2006 292 351 - - 1+ -28 5th Oct. 2006 291 358 - - 1+ -29 5th Oct. 2006 273 296 - - - -30 5th Oct. 2006 296 381 0.43 M 1+ V 31 5th Oct. 2006 262 249 - - 1+
-calculated as follows:
GSI = [GW / (BW - GW)] × 100.
Otolith preparation
Sagittal otoliths of the mullet were removed, ultrasonically cleaned with distilled water, and air-dried. The otoliths were then embedded in epoxy resin (Epofix, Struers), and cut transversely with a low-speed diamond saw to obtain a section of approximately 300 μm thick, which was polished to expose the primordium. The polished otolith thin sections were cleaned with distilled water and then etched with a 5% HCl solution for 10 s to enhance the annuli in the otolith. The otolith was photographed with a digital camera under a light microscope (Nikon) with reflected light. The fish age was determined by counting the annuli, and the distance of each annulus to the otolith core was measured for comparison with the otolith elemental analyses so as to understand their temporal changes (Fig. 2). Nine of the mullet otoliths were slightly over-polished such that the primordium was missing. The annuli number could be counted from the edge but those samples were not included in further elemental composition analyses.
Otolith elemental composition measurement and analyses
An otolith section was re-polished, washed in 5% H2O2, triple-rinsed with distilled water, and
air-dried. The elemental composition of the otolith was measured along its dorsoventral axis by ICP-MS (Finnigan MAT ELEMENT 2, Thermo Electron Corp., Waltham, MA, USA) equipped with a laser
microprobe (Merchantek LUV 266 Nd: YAG UV, New Wave Research, Fremont, CA, USA) which was pulsed at 20 Hz with a spot size of 150 μm in diameter. The laser beam was moved along the line transect (Fig. 1) at a speed of 15 μm/s to continuously measure the change in the otolith elemental composition. Isotopes of 44Ca, 88Sr, and 138Ba were measured because the sensitivities of
these isotopes remained at least 10 times higher than background levels. Standard (National Institute of Standards and Technology 612 glass standard) counts were also measured before measurement of every 3 or 4 otoliths to calibrate the otolith elemental concentration and to monitor instrument analytical precision. At the start of each otolith analysis, background counts were collected for 30 s, and the average was subtracted from sample counts to correct for background levels. The ablation chamber was purged for 60 s after sampling each otolith. All counting data were expressed as ratios of an element to the Ca concentration (ppm/ppm) by estimating the relative response factor of the instrument to the known concentration in the standard (NIST 612). Counts of the elements measured along the line transect of the otolith were collected every 2.11 s at an interval of approximately 32 μm in spatial resolution.
Data analysis
The core region of the mullet otolith was deposited during the larval stage. The duration of the mullet larval stage was estimated to be approximately 14 d, and the otolith diameter at that age was about 150 μm (Chang 2003). Thus the average value of the 3 points taken from the core section (approximately 96 μm in radius around
Fig. 2. Thin transverse section of a Mugil cephalus otolith (2+ yr old) under reflected light after etching with HCl. The dotted line
indicates the analyzed LA-ICPMS transect. Black arrow, annulus; C, core; D, dorsal; E, external; I, internal; V, ventral. Scale bar = 500 μm. V E I C D
the primordium) (Fig. 2) of the otolith represented the habitat signature for each individual in the early larval stage. The otolith core values were examined by a principal component analysis (PCA) to test whether mullet collected in midstream of the Tanshui River were of a similar origin. The otolith element: Ca concentration ratios were examined by t-test for differences in origin using STATISTICA© vers. 6.1. Ba: Ca ratios were
log-transformed for all statistical analyses in order to meet the hypotheses for a normal distribution and homogenity of the variance.
RESULTS Size composition
Mugil cephalus specimens contained 12
females, 8 males, and 11 undetermined individuals, and the fork length and weight ranged 237-380 mm and 189-833 g, respectively. Fork lengths were
317 ± 32 (mean ± SD), 307 ± 31, and 276 ± 21 mm for female, male, and undetermined individuals, and total weights were 481 ± 141, 443 ± 162, and 308 ± 69 g, respectively. Fish ages were between 1+ and 2+ yr, and 65% of fish (n = 20) were 1+ yr old. The mean (± SD) GSI was 1.2 ± 1.1 with the highest GSI values at 3.1 (for males) and 3.6 (for females). The GSI and age composition indicated that all 31 fish collected from the freshwater habitat of the river were immature.
Elemental composition in the otolith edge
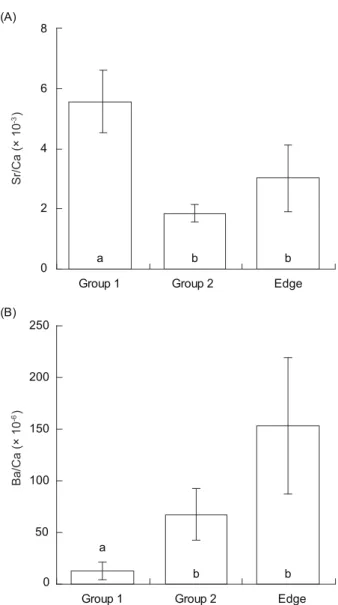
Sr/Ca and Ba/Ca ratios in the mullet otolith edges were used as the baseline of a freshwater habitat. The mean Sr/Ca and Ba/Ca ratios were (3.0 ± 1.1) × 10-3 and (153 ± 66) × 10-6, respectively
(Fig. 3). The otolith Sr/Ca ratio in freshwater-habitat mullet was in the same range at around 3.0 × 10-3 from a tank-rearing experiment (Chang
et al. 2004b), so this was set as the threshold value for discriminating fresh and brackish water.
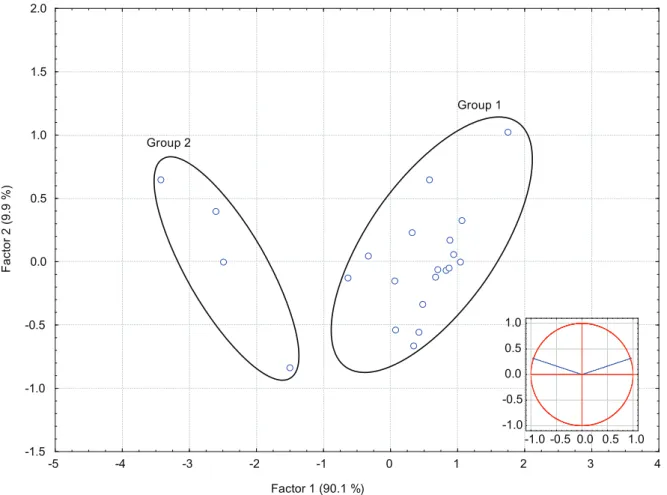
-5 -4 -3 -2 -1 0 1 2 3 4 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 Group 2 Group 1 Factor 1 (90.1 %) Fa ct or 2 (9 .9 % ) -1.0 -0.5 0.0 0.5 1.0 -1.0 -0.5 0.0 0.5 1.0
Fig. 3. The principle components analysis (PCA) grouping into 2 types of Mugil cephalus from the Tanshui River classified by the otolith
core elemental composition and relative contributions of each elemental ratio. The elliptical circle indicates the 95% confidence limit of each group.
Ba/Ca ratios showed larger varations, and the value of 124 × 10-6, which was the lower limit of the
mean, was set as the threshold value.
Elemental composition within the otolith core
The PCA classified the elemental composition in the core region of mullet collected in fresh water in the Tanshui River into 2 groups (Fig. 4). The Sr/Ca and Ba/Ca ratios in the core regions of the otoliths significantly differed between the groups (t-test, t = -6.908, d.f. = 20, p < 0.001 for Sr/Ca;
t = 5.530, d.f. = 20, p < 0.001 for log Ba/Ca) (Fig.
3). For group 1, the core and edge Sr/Ca and
Ba/Ca ratios significantly differed (t-test, t = -7.429,
d.f. = 38, p < 0.001 for Sr/Ca; t = 13.692, d.f. = 38, p < 0.001 for log Ba/Ca); while for group 2, they
were similar (t-test, t = -2.071, d.f. = 24, p = 0.049 for Sr/Ca; t = -2.521, d.f. = 24, p = 0.018 for log Ba/Ca). Group 1 fish comprised 82% (n = 18) of the fish examined and were characterized by higher Sr/Ca ratios at (5.6 ± 1.0) × 10-3 and lower
Ba/Ca ratios of (12 ± 8) × 10-6 in the otolith core,
indicating that they inhabited a marine environment during the early larval stage. The otolith core of group 2, which comprised 18% (n = 4) of the mullet examined, was characterized by a significantly lower Sr/Ca ratio of (1.9 ± 0.3) × 10-3 (mean ± SD)
and a higher Ba/Ca ratio of (67 ± 25) × 10-6 (Fig. 3),
indicating that these fish (numbers 17, 20, 21, and 25) inhabited a different environment from group 1 during the early larval stage.
Temporal changes in otolith Sr/Ca and Ba/Ca ratios beyond the core region
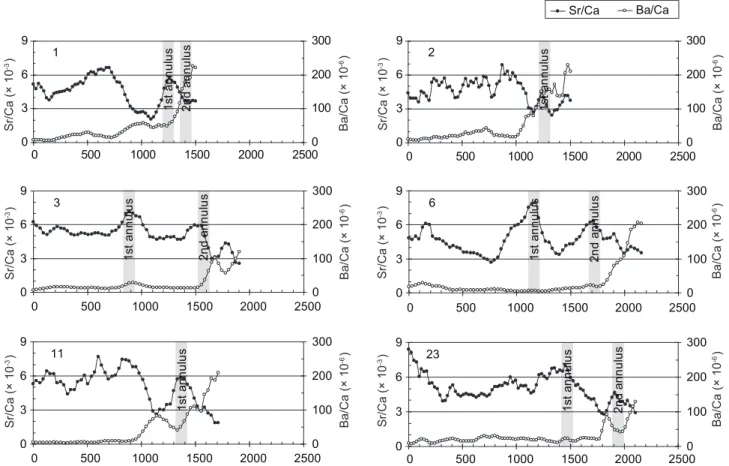
Temporal changes in the otolith Sr/Ca and Ba/Ca ratios of the mullet differed between individuals and were divided into 2 types. Temporal changes of 6 otoliths (type I) had higher Sr/Ca ratios (> 3 × 10-3), and the Ba/Ca ratios
were consistently < 20 × 10-6 before the annulus
near the otolith edges (Fig. 5). For example, fish no. 3 showed consistently high otolith Sr/Ca ratios beyond the core to the position of the 2nd annuli, then dropped to the lower brackish and freshwater level of < 3 × 10-3 after the 2nd annulus formed.
In some fish (nos. 3, 6, and 23), the Sr/Ca ratios were high at the 2 annuli and lower in between them. Otolith Ba/Ca ratios remained at a low level (< 20 × 10-6) until the 2nd annulus, after which
the Ba/Ca ratio suddenly increased and reached a peak at the otolith edge (> 120 × 10-6). The
otolith chemical signatures of high Sr/Ca and low Ba/Ca ratios indicated that those fish inhabited a marine environment during the juvenile and most of the subadult stages and entered fresh water just before being captured.
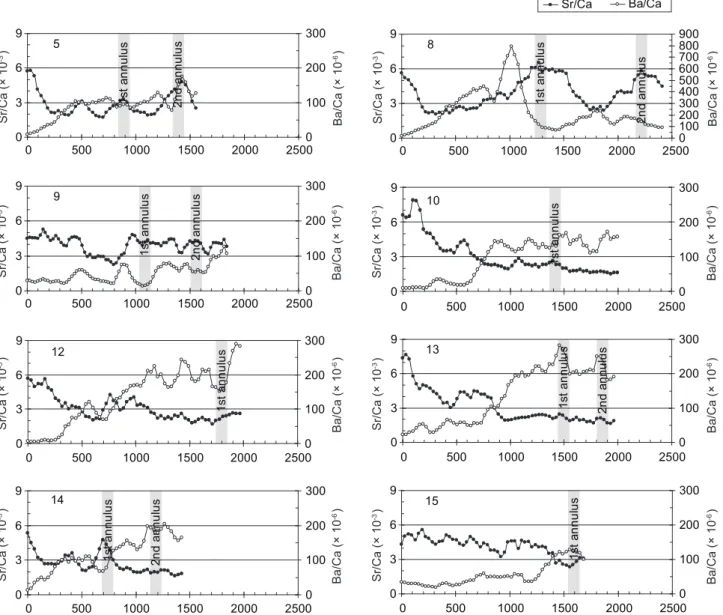
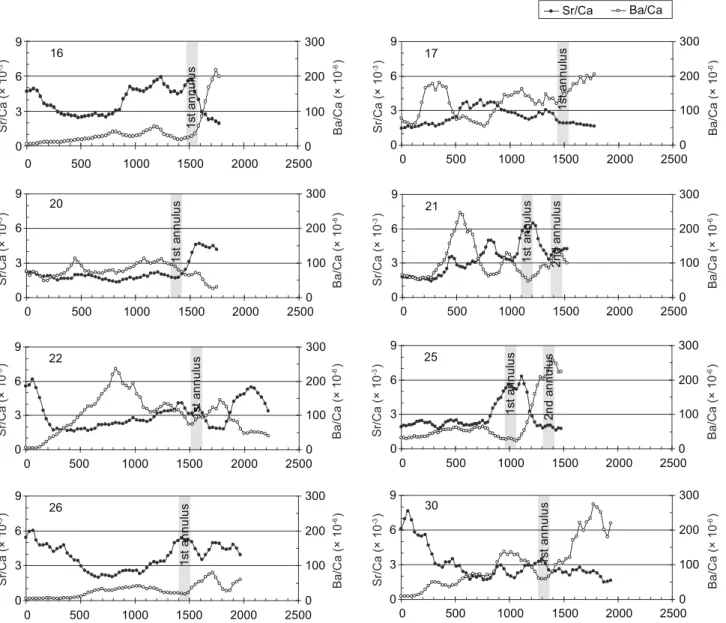
Type II fish had fluctuating Sr/Ca ratios beyond the core region (Fig. 6). In most of those individuals, the otolith Sr/Ca ratios decreased to a freshwater level of ≤ 3 × 10-3 beyond the
core region (Fig. 6). Some otolith Sr/Ca ratios fluctuated, but some stayed consistently low before deposition of the 1st annulus. The Ba/Ca ratios were low in the otolith core, then gradually increased and finally fluctuated between (100 and 200) × 10-6 (Fig. 6). Those individuals stayed 0
2 4 6 8
Group 1 Group 2 Edge
a b b (A) 0 50 100 150 200 250
Group 1 Group 2 Edge
a b b (B) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6)
Fig. 4. Otolith Sr/Ca (A) and Ba/Ca (B) ratios at the edge of
all individual mulletotolith cores (Edge) and the cores of the 2 groups of Mugil cephalus from the Tanshui River. The same letter indicates that otolith ratios are homogeneous among the groups.
in fresh- and brackish-water habitats and never returned to the marine environment again after being recruited into the estuary at the juvenile stage. Individuals with lower Sr/Ca ratios in the otolith core (group 2 in Fig. 4) were all classified into this type.
DISCUSSION
Validation of otolith Sr/Ca and Ba/Ca ratios as an indicator of mullet habitat
The mullet were collected in the middle of the river where the salinity at the bottom was close to 0, although the water level was influenced by tidal fluctuations. The chemical signatures with low Sr/Ca (3.0 × 10-3) and high Ba/Ca (124 × 10-6)
ratios at the otolith edge indicated that the mullet had recently inhabited fresh water, as indicated by their location of capture. This conclusion was supported by previous studies that otolith Sr/Ca ratios are < 3 × 10-3 when mullet are in fresh
water (at salinities of 0-5 psu) and are > 3 × 10-3
in saline water (5-35 psu) (Chang et al. 2004a b). Accordingly, as for other diadromous fishes, the otolith Sr/Ca ratios can be used to indicate fish movement between fresh and saline (estuarine and seawater) environments (Tzeng et al. 2002). Otolith Ba/Ca ratios for mullet in relation to the ambient water salinity have not been validated. However, otolith Ba/Ca ratios were reported to be higher in fresh than in seawater for other fishes (Elsdon and Gillanders 2004 2005). In this study, we found that Ba/Ca ratios in otoliths were higher when the mullet were in fresh water (124 × 10-6)
and lower in seawater (~20 × 10-6, the higher
limit of the mean of the core in group 1 (Fig. 4). Therefore, by combining otolith Sr/Ca and Ba/Ca ratios, we could reconstruct the mullet migratory environmental history in greater detail. Low otolith Sr/Ca ratios (< 3.0 × 10-3) can be used to
discriminate freshwater from marine habitats, while low otolith Ba/Ca ratios (< 20 × 10-6) can be used
to discriminate marine from freshwater habitats. When the chemical signatures fall between the 2 criteria, then an estuarine habitat is suggested.
0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 1 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 3 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 11 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 23 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 6 1s ta nn ul us 2n d an nu lu s Sr/Ca (× 10 -3) Ba /C a (× 1 0 -6) Sr/Ca Ba/Ca Sr/Ca (× 10 -3) Ba /C a (× 1 0 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 2 1s ta nn ul us Sr/Ca (× 10 -3) Ba /C a (× 1 0 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba /C a (× 1 0 -6)
Identification of habitat use during the larval stage by Sr/Ca and Ba/Ca ratios
A f t e r s p a w n i n g o ff s o u t h w e s t e r n o r northeastern Taiwan (Chen and Su 1986, Huang and Su 1989, Chang et al. 2004a, Hsu et al. 2007), larvae of M. cephalus passively disperse to estuarine nursery areas (Chang et al. 2000, Hsu et al. 2009). Larvae younger than 14 d old do not actively swim (Tung 1967, Liao 1975); therefore, these larvae would still be resident in the water mass where they hatched. Accordingly, the elemental composition of the 1st 3 measurement points in the otolith (approx. 48 μm from the primordium) was regarded as characteristic of the ambient water of the spawning
ground. In the present study, 2 different signals near the otolith core were found (Fig. 4). Group 1 was characterized by higher Sr/Ca ratios and comprised larger percentages (82%) of freshwater resident mullet, while Sr/Ca ratios in the core of group 2 fish were < 3 × 10-3. This indicated that
group 2 fish spent their early life in a water mass that differed from that of group 1. The lower Sr/Ca ratios in group 2 otolith cores suggest that their parents spawned in a low-salinity water mass, or the distance for larval dispersal to the estuarine nursery area was short, thus making the average core Sr/Ca value significantly lower than that of the other group. It is not known whether mullet spawn outside the known spawning grounds of southwestern and northeastern Taiwan. A 3rd
Fig. 6. Temporal changes in otolith Sr/Ca and Ba/Ca ratios for migratiory type II Mugil cephalus collected from the Tanshui River. 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 5 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 400 500 600 700 800 900 8 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 9 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 12 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 14 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 10 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 13 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 15 1s ta nn ul us Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca Ba/Ca
spawning population of M. cephalus may explain the appearance of mullet juveniles in estuaries between autumn and the following spring, while juveniles of the traditional spawning population are only recruited during midwinter in northern and southern waters of Taiwan (Chang et al. 2000, Hsu et al. 2007). Our new discovery of different otolith core signatures implies that another spawning population of M. cephalus, with different life history traits, exists in the waters of Taiwan and East Asia. Elliott and Dewailly (1995) indicated that M.
cephalus spend their entire lives in the estuary and
do not enter offshore waters. Our results indicated that M. cephalus has more-diverse spawning and nursery ground behaviors than expected.
The migratory environmental history of freshwater resident mullet
By analyzing temporal changes in the otolith Sr/Ca and Ba/Ca ratios of freshwater resident mullet, 2 migratory types were found (Figs. 5, 6). Type I mullet (27%) contained high Sr/Ca and low Ba/Ca ratios, indicating that those fish mostly inhabited marine environments. Type II mullet (73%) had fluctuating Sr/Ca and Ba/Ca ratios beyond the core region, indicating that individuals stayed in freshwater and estuarine habitats and never returned to the marine environment again after the juvenile stage. Migratory environmental histories of mullet caught in estuarine, nearshore, and offshore waters were studied by Chang et
Fig. 6. (continued) 0 3 6 9 0 500 1000 1500 2000 2500 0 100 200 300 16 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 17 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 20 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 22 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 26 1s ta nn ul us 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 21 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 25 1s ta nn ul us 2n d an nu lu s 0 3 6 9 0 500 1000 1500 2000 25000 100 200 300 30 1s ta nn ul us Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca (× 10 -3) Ba/Ca (× 10 -6) Sr/Ca Ba/Ca
al. (2004a), and similar migratory types were identified. However, mullet from high-salinity water comprised 67% of type I and only 33% of type II. If only the traditional spawning population caught during midwinter was considered, an even lower percentage of type II remained (11.1%) (Chang et al. 2004a). In our study, mullet collected in fresh water were predominantly type II (estuarine and freshwater), and some individuals showed a different pattern in the otolith core (group 2 in figure 4). This indicates that there is a mixture of migratory and resident mullet near Taiwan. Although most individuals recruited from the southeastern Taiwan spawning area may return to the sea after the juvenile phase, a resident mullet population may exist that inhabits estuarine and freshwater habitats and seldom returns to the sea. Juveniles may reside in estuaries and freshwater rivers around Taiwan and not join the northerly maturation migration. Thus the migratory patterns of freshwater mullet may differ from those of nearshore and offshore areas.
Acknowledgments: This study was financially
supported by the National Science Council of Taiwan (NSC93-2313-B-002-054). The authors would like to thank Mr. B.M. Jessop for helpful comments on an early draft of the manuscript.
REFERENCES
Blaber SJM. 1987. Factors affecting recruitment and survival of mugilids in estuaries and coastal waters of southeastern Africa. Am. Fish. Soc. Symp. 1: 507-518.
Bradbury IR, SE Campana, P Bentzen. 2008. Otolith elemental composition and adult tagging reveal spawning site fidelity and estuarine dependency in rainbow smelt. Mar. Ecol.-Prog. Ser. 368: 255-268.
Campana SE. 1999. Chemistry and composition of fish otolith: pathways, mechanisms and application. Mar. Ecol.-Prog. Ser. 188: 263-297.
Campana SE, GA Chouinard, JM Hanson, A Fréchet, J Brattey. 2000. Otolith elemental fingerprints as biological tracers of fish stocks. Fish. Res. 43: 129-139.
Chang CW. 2003. Use of otolith microstructure and microchemistry to study life history and migratory environment of grey mullet Mugil cephalus in the coastal waters of Taiwan. PhD dissertation, National Taiwan University, Taipei, Taiwan.
Chang CW, Y Iizuka, WN Tzeng. 2004a. Migratory environmental history of the grey mullet Mugil cephalus as revealed by otolith Sr: Ca ratios. Mar. Ecol.-Prog. Ser.
269: 277-288.
Chang CW, SH Lin, Y Iizuka, WN Tzeng. 2004b. Relationship between Sr: Ca ratios in otoliths of grey mullet Mugil cephalus and ambient salinity: validation, mechanisms, and applications. Zool. Stud. 43: 74-85.
Chang CW, WN Tzeng. 2006. Spatial and temporal variations of grey mullet resource in the coast of Taiwan: species identification, production and aquaculture. Fish. Extens. Natl. Taiwan Univ. 18: 25-42. (in Chinese)
Chang CW, WN Tzeng, YC Lee. 2000. Recruitment and hatching dates of grey mullet (Mugil cephalus L.) juveniles in the Tanshui estuary of northwest Taiwan. Zool. Stud.
39: 99-106.
Chen WY, WC Su. 1986. Reproductive biology of the grey mullet, Mugil cephalus L. of Taiwan. In CR Lee, ed. Study on the resource of grey mullet in Taiwan, 1983-1985. Kaohsiung, Taiwan: Kaohsiung Branch of the Taiwan Fisheries Research Institute, pp. 73-80.
Chen WY, WC Su, KT Shao, CP Lin. 1989. Morphometric studies of the grey mullet (Mugil cephalus) from the waters around Taiwan. J. Fish. Soc. Taiwan 16: 153-163.
Chester R. 2000. Chapter 3. The transport of material to the oceans: the river pathway. In R Chester, ed. Marine geochemistry. Oxford, UK: Blackwell Sciences, pp. 11-51. Edmond JM, A Spivack, BC Grant, MH Hu, Z Chen, S Chen,
X Zeng. 1985. Chemical dynamics of the Changjiang estuary. Cont. Shelf Res. 4: 17-36.
Elliott M, F Dewailly. 1995. The structure and components of European estuarine fish assemblages. Netherl. J. Aquat. Ecol. 29: 397-417.
Elsdon TS, BM Gillanders. 2004. Fish otolith chemistry influenced by exposure to multiple environmental variables. J. Exp. Mar. Biol. Ecol. 313: 269-284.
Elsdon TS, BM Gillanders. 2005. Alternative life-history patterns of estuarine fish: barium in otoliths elucidates freshwater residency. Can. J. Fish. Aquat. Sci. 62:
1143-1152.
Fowler AJ, BM Gillanders, KC Hall. 2005. Relationship between elemental concentration and age from otoliths of adult snapper (Pagrus auratus, Sparidae): implications for movement and stock structure. Mar. Freshw. Res. 56:
661-676.
Hsu CC, CW Chang, Y Iizuka, WN Tzeng. 2009. A growth check deposited at estuarine arrival in otoliths of juvenile flathead mullet (Mugil cephalus L.). Zool. Stud. 48:
315-324.
Hsu CC, YS Han, WN Tzeng. 2007. Evidence of flathead mullet Mugil cephalus L. spawning in waters northeast of Taiwan. Zool. Stud. 46: 717-725.
Hsu CC, WN Tzeng. 2009. Validation of annular deposition in scales and otoliths of flathead mullet Mugil cephalus. Zool. Stud. 48: 640-648.
Huang CS, WC Su. 1989. Studies on the fluctuations of fishing conditions for grey mullet (Mugil cephalus Linnaeus) from the western coast of Taiwan. J. Fish. Soc. Taiwan 16:
47-83.
Huang CS, CF Weng, SC Lee. 2001. Distinguishing two types of gray mullet, Mugil cephalus L. (Mugiliformes: Mugilidae), by using glucose-6-phosphate isomerase (GPI) allozymes with special reference to enzyme activities. J. Comp. Physiol. B 171: 387-394.
Lee SC. 1992. Fish fauna and abundance of some dominant species in the estuary of Tanshui, northwestern Taiwan. J. Fish. Soc. Taiwan 19: 263-271.
Liao IC. 1975. The experiments on the induced breeding of the grey mullet in Taiwan from 1963-1973. Aquaculture 6:
31-58.
Liu CH. 1986. Survey of the spawning grounds of grey mullet. In WC Su, ed. Study on the resource of grey mullet in
Taiwan, 1983-1985. Kaohsiung, Taiwan: Kaohsiung Branch of the Taiwan Fisheries Research Institute, pp. 63-72.
Loher T. 2008. Homing and summer feeding site fidelity of Pacific halibut (Hippolossus stenolepis) in the Gulf of Alaska, established using satellite-transmitting archival tags. Fish. Res. 92: 63-69.
Morales-Nin B, SC Swan, JDM Gordon, M Palmer, AJ Geffen, T Shimmield, T Sawyer. 2005. Age-related trends in otolith chemistry of Merluccius merluccius from the north-eastern Atlantic Ocean and the western Mediterranean Sea. Mar. Freshwater Res. 56: 599-607.
Pannella G. 1971. Fish otoliths: daily growth layers and periodical patterns. Science 173: 1124-1127.
Ruttenberg BI, SL Hamilton, MJH Hickford, GL Paradis, MS Sheehy, JD Standish, O Ben-Tzvi, RR Warrner. 2005.
Elevated levels of trace elements in cores of otoliths and their potential for use as natural tags. Mar. Ecol.-Prog. Ser. 297: 273-281.
Thomson JM. 1966. The grey mullets. In H Barnes, ed. Oceanography and marine biology - an annual review. London: Allen and Unwin, pp. 301-355.
Tung IH. 1967. On the egg development and larval stage of the grey mullet, Mugil cephalus Linnaeus. Rep. Inst. Fish. Biol. Minist. Econ. Aff. Natl. Taiwan Univ. 3: 187-210.
Tung IH. 1981. On the fishery biology of gray mullet, Mugil cephalus L., in Taiwan. Rep. Inst. Fish. Biol. Minist. Econ. Aff. Natl. Taiwan Univ. 3: 38-102.
Tzeng WN, JC Shiao, Y Iizuka. 2002. Use of otolith Sr: Ca ratios to study the riverine migratory behaviors of Japanese eel Anguilla japonica. Mar. Ecol.-Prog. Ser.