行政院國家科學委員會專題研究計畫 成果報告
玉葉金花花部器官決定基因之功能與蛋白質交互作用研究
(3/3)
研究成果報告(完整版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2311-B-002-001- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學生態學與演化生物學研究所 計 畫 主 持 人 : 胡哲明 共 同 主 持 人 : 賴爾 計畫參與人員: 碩士級-專任助理:曾妤馨 博士班研究生-兼任助理:徐馨怡、張世白 處 理 方 式 : 本計畫可公開查詢中 華 民 國 96 年 12 月 28 日
行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告 □ 期 中 進 度 報 告玉葉金花花部器官決定基因之功能與蛋白質交互作用研究(3/3)
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 93 - 2311 - B - 002 - 024 -
計畫編號:
NSC 94 - 2311 - B - 002 - 007 -
計畫編號:
NSC 95 - 2311 - B - 002 - 001 -
執行期間:民國93年08月01日至 96年07月31日
計畫主持人:胡哲明
共同主持人:賴爾珉
計畫參與人員: 張世白,徐馨怡,曾妤馨
成果報告類型(依經費核定清單規定繳交):■精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計
畫、列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:國立台灣大學生態學與演化生物學研究所
中
華 民 國 96 年 12 月 25 日
I. Abstract 1. 中文摘要
毛玉葉金花(Mussaenda pubescens)是台灣低海拔中常見的植物之一,常可以見
到它花萼其中的一個裂片尖端膨大成一個外形類似葉子、但顏色為白色或紅色,稱為 葉狀萼片(calycophylls, 或floral semaphylls)的構造。而玉葉金花名稱的由來,即源於 它的花序中黃色的花瓣和白色的葉狀萼片而來。這些葉狀萼片不管在細胞形狀或細胞 排列上,都與正常的花瓣類似,加上有著顯著的顏色,一般皆認為這些葉狀萼片在功 能上與花瓣無異,因此也被認為是類花瓣(petaloid)的構造。我們在這個三年期的研究 計畫中,期望藉由花部基因的選殖與功能性的研究,希望瞭解有關花瓣與類花瓣調控 的各類因子。先前的研究結果顯示兩個B群基因產物MupDEF及MupGLO有交互作用, 而MupTM6不與另外兩個B群蛋白作用。在本研究中,我們建構了玉葉金花整個花部及 膨大萼片的cDNA library,並以在膨大萼片中表現的兩個B群基因MupDEF及MupTM6 為餌進行可能交互作用蛋白的篩選,不過未能得到具正向反應的基因。另外我們也獲 得了一個B群基因的可能下游基因MPF2在毛玉葉金花中的同源基因MupMPF2。MPF2 在酸漿(Physalis)已被證實與其膨大花萼有關。我們檢測MupMPF2在毛玉葉金花的各部 器官的表現,結果顯示該基因確實會在毛玉葉金花的膨大花萼表現,故推測該基因可 能與該膨大花萼的形成有關。另外南方雜合分析的結果顯示,在毛玉葉金花中可能仍 有其他B群基因的副本,這些重複的片段是否參與毛玉葉金花的膨大花萼表現,仍須 進一步的研究探討。 關鍵詞:玉葉金花,花部器官決定基因,蛋白質交互作用,MPF2 2. English abstract
Mussaenda pubescens is a plant with enlarged and petaloid calyx, named calycophylls,
which are phenotypically similar to true petals in having bright color and papilliate epidermal cells. We proposed the petaloid structure were formed by the ectopic expression of genes that specifying petal organ identity, but our preliminary data suggested that there might be
unknown factors involved other than floral ABE genes. In our previous studies, the result reveals the interactions between the two B-class proteins, MupGLO and MupDEF, and MupTM6 did not interact with other B-class proteins, even though it is generally thought TM6 and DEF homologues are redundant copies. In this year, we constructed a cDNA library of flower buds and enlarged sepals, and screened with the MupDEF and MupTM6, two B-class genes expressed in petaloid sepal. Unfortunately, no positive transformant was selected in our screening. We also cloned a putative target of B-class genes, MPF2, which is
responsible for the enlarged calyx in Physalis, from M. pubescens. We named this gene
MupMPF2. The expression pattern based on RT-PCR showed it is expressed in many tissues,
including the enlarged sepals. This suggests that it may play a role in the petaloid structure. We also test the copy numbers of these five floral ABE genes with southern blot. The result shows there may be two copies of MupGLO and MupDEF in the genome, and the functions of the possible redundant copies remain obscured.
Keywords: Mussaenda pubescens, floral organ identity genes, protein-protein interactions,
MPF2
II. Introduction
In most flowering plants, petals are generally used as an attractant agent for luring pollinators. However, there are many examples that use non-petal organs to mimic showy petals, and thus called "petaloid structures". These petaloid structures are usually found in sepals and bracts, from diverse taxa in angiosperms (Albert et al., 1998). For example, some of the dogwoods (Cornus) have showy bracts that serve as attractants to pollinators just like normal petals do. Similar situations also can be found in the calyx of some Hydrangea species (Hydrangeaceae), whereas the calyx is expanded and become petal-like in the peripheral flowers. In the flowers of Mussaenda species (Rubiaceae), there are one to several sepals in a flower expands during maturation and the enlarged sepal(s) will turn white or red like a showy petal. This special structure is called a calycophyll in Rubiaceae (Claben-Blockhoff, 1996). The calycophylls have vivid white or red color, loose mesophylls, and papilliate epidermis, which are all typical features of a true petal. The formation of enlarged sepals is
not restricted to Mussaenda and has been named 'inflated-calyx syndrome (ICS)', which is also found in Anisodus, Physalis, Withania of Solanaceae (He et al. 2004) and Hydrangea of Hydrangeaceae.
In order to elucidate the possible genetic mechanism for the formation of petaloid structures, we selected Mussaenda pubescens as our working system. M. pubescens has an enlarged and petaloid calycophylls on the peripheral flowers of a inflorescence. Based on the current floral genetic model, petals are determined by combination of three classes of genes, A (e.g. AP1 and AP2), B (e.g. PI and AP3), and E (SEPs). In Arabidopsis studies,
overexpression a combination of AP1, AP3, PI, and SEP showed an interesting phenotype that converts all its leaves into petals (Honma and Goto, 2001). Theses results suggest that, petaloid structures could be regulated by ectopic expression of petal identity genes, as proposed by several authors (Bowman, 1997; Albert et al., 1998; Baum, 1998; Baum and Whitlock, 1999). The project's rationale is to apply this model to the petaloidy regulation in
M. pubescens.
Floral development has been best studied in two model systems, Arabidopsis and
Antirrhinum. Genetic studies of these two systems have shown that flower organ identities
are determined by a combination of three classes of genes (A, B, and C), and has been
described as the "classic ABC model" (Coen and Meyerowitz, 1991). Recently, several other genes are found to be involved in floral organ formation, and the model is further elaborated as "ABCDE model" that incorporates SEEDSTICK (STK) and SEPALLATA (SEP) genes in the case of Arabidopsis (Pelaz et al., 2000; Theiben, 2001). In these model, the organ identity of first whorl is controlled by A class gene, the second whorl is controlled by A+B+E class genes, the third whorl is controlled by B+C+E class genes, and the forth whorl is controlled by C+E class genes. D class genes refer to genes responsible to ovule development. In angiosperms, the A class genes are represented by homologues of the Arabidopsis thaliana
APETALA1 (AP1) and APETALA2 (AP2) genes, whereas the B class genes are represented by
homologues of the A. thaliana APETALA3 (AP3) and PISTILLATA (PI) genes; and the C class genes are represented by homologues of the A. thaliana AGAMOUS (AG). E class genes are represented by homologues of the A. thaliana SEPALLATA genes. Based on current studies, many other angiosperms may also share similar developmental program since these genes are
very conserved among flowering plants (Bowman, 1997; Kramer et al., 1998; Ma and dePamphilis, 2000), although some exceptions can be found in basal angiosperm and lower eudicots (Kramer and Irish, 1999, 2000). For example, the expression patterns of B class genes are variable in time and regions in the Ranunculiales species (Kramer et al. 1998, 2003).
A “floral quartet” model has been proposed to explain the protein-protein interactions in regards to floral organ development (Theissen and Saedler, 2001). The model states that there are four components of protein complex involved in determining floral organ identity based on known protein-protein interaction patterns (Theissen, 2001). Based on this model, petal identity is determined by a tetramer protein complex consists of AP3, PI, AP1, and SEP. It is therefore any mutations in these genes will cause abnormality in petals, and ectopic expression will lead to formation of petaloid structures. Although the validity of this model is still remain to be test, other studies on protein-protein interaction of floral identity genes have suggested a complex pattern existed among seed plants. For example, in Arabidopsis and
Antirrhinum, heterodimerization of two B class genes is absolutely required for DNA binding
in vitro (Reichmann et al., 1996; Egea-Cortines et al., 1999). However, it is evident that B class proteins form homodimers rather than heterodimers in gymnosperms like Gnetum, and monocots such as Lilium and Oncidium (Winter et al., 2002; Hsu & Yang, 2002). We have very limited information on the correlation of protein complex formation and the petal
identity in other flowering plants. Most of the data are scarce or with only in vitro assay, and this field definitely needed to be explored.
In our previous studies, we have successfully identified A, B, and E class genes from
M. pubescens, namely MupSQUA (A class homologue); MupDEF, MupGLO, MupTM6 (all B
class homologues); and MuSEP (E class homologue). The expression patterns based on RT-PCR results suggest that B and E class genes are expressed in the normal sepals as well as calycophylls. The only exception is MupGLO, its transcript is not detected in calycophylls and very weak in normal sepals.
The almost equal levels of B class gene expression in calycophylls and normal sepals of M. pubescens suggest that there must be some other factors involved in the formation of calycophylls. In order to identify additional factors involved in calycophyll formation, a
screen for proteins that interact with MupDEF, MupTM6, and/or MupSEP was carried out. This is based on the rationale that multimerization is likely present in floral identity gene products (Theiben, 2001; Lohmann & Weigel, 2002). Yeast two-hybrid screening system was employed using the previously identified ABE genes as baits, to screen the potential
counterparts from a constructed floral cDNA library, such system has successfully identified putative interacting proteins in rice (Lim et al., 2000; Masiero et al., 2002).
III. Materials and methods
1. Sample collections and RNA isolation
Plant materials of Mussaenda pubescens have been collected from the populations at Shu-Mei-Pin (Taipei Co.) and Kenting (Pingtung Co.). Total RNAs were isolated from different organs of Mussaenda pubescens by Pine Tree method (Chang et al. 1993). mRNA was isolated by PolyAtract mRNA isolation systems kit (Promega). cDNA were synthesis from mRNA by HybriZAP® -2.1 XR cDNA synthesis Kit (Stratagene).
2. Full-length open reading frame of cDNAs
Since previously identified ABE genes were obtained through 3' RACE (Rapid Amplification of cDNA Ends, Invitrogen, Life technologies, Carlsbad, CA, USA), specific primers were designed at K domain region of previously identified ABE genes for 5' RACE (Rapid Amplification of cDNA Ends, Invitrogen, Life technologies, Carlsbad, CA, USA) reaction in order to obtain 5' region of the genes. The 5' RACE products were then cloned into pGEM®-T vector (Promega, Madison, WI, USA), and determined the sequences. New specific primers at the 5' end of the genes were then designed to amplify full-length cDNAs in conjugated with 3' specific primers for PCR. The products were then cloned again into pGEM®-T vector (Promega, Madison, WI, USA), and determined the sequences.
3. Yeast two-hybrid screening
Construction and screening of cDNA library
The cDNAs were constructed into the lambda vector using the HybriZAP 2.1® two-hybrid system, into pAD-GAL4 and pBD-GAL4 following the manufactory's suggestions
(Stratagene, La Jolla, CA, USA). New primers were designed to include a EcoRI or SalI overhead at their 5' (Table 1) for amplifying open reading frame of cDNAs. The flower buds and enlarged sepal cDNAs were constructed in the HybriZAP-2.1 vector with the GAL4 activation domain to generate the primary lambda phage library. After amplification, phage library was converted to the phagemid (pAD-GAL4-2.1) library by mass excision. Phagemid DNA were isolated by mini-prep plasmid DNA extraction kit (Viogene). The library DNA or pAD-MupGLO as the positive control was co-transformed into yeast strain YRG2 with pBD-MupDEF or pBD-MupTM6 as the baits. Positive transformants were screened on synthetic dropout (SD) plates under His-, Leu- and Trp- selective condition. Transformation efficiency was evaluated with the growth of colonies on SD plates lacking Leu and Trp. Then filter lift assay to detect β-galactosidase activity was used to confirm the protein-protein interactions.
Table 1. Primer design for yeast-2-hybrid constructs
Primer name Sequences(5'→ 3')
MuAP1_Eco ggAATTCATggggAgAggAAAg MuAP1_Sal ACgCgTCgACgggCTTTATCAggC MuTM6_Eco ggAATTCATgggTCgTgggAAg MuTM6_Sal ACgCgTCgACCATgCAATAggATg MuGLO_Eco ggAATTCATggggAgAggTAAg MuGLO_Sal ACgCgTCgACgTTCTTgggTAgAgC MuDEF_Eco ggAATTCATggCTCgTgggAAg MuDEF_Sal ACgCgTCgACgTAAATAAATTTGCTAC MuSEP_Eco ggAATTCATgggAAgAggTAgg MuSEP_Sal ACgCgTCgACgTATCATggTAACC
4. Identification of MPF homologues in M. pubescens
(1) RNA isolation and amplification of MPF homolog
Total RNA was extracted by using the Pine tree method (Chang et al., 1993) with some modifications. For first-strand cDNA synthesis, nearly 1 µg RNA was mixed with reverse transcriptase (Invirtogen, Life Technologiesm Carsbadm CA, USA) in 20 µl reaction volume with primer PolyT-R1 (5’-CCGGATCTCTAGACGGCCGCTTT- TTTTTTTTTTTTTT-3’), followed by polymerase chain reaction (PCR) amplification with degenerate primer- DSVP
(5’-ATGGCRAGRSARAARATTCAGATMAAG- AA-3’) (modified from primer DSVP.1 in Hecht et al., 2005). PCR reactions were performed with T-Gradient thermocycler
(Biometra, Gottingen, UK) using the following program: 95ºC for 15min; 95ºC for 45 sec, 60ºC for 45 sec, 68ºC for 45 sec (35 cycles); 68ºC for 7 min and stopped at 4ºC. The 1000-bp PCR product obtained was cloned into the pGEM-T easy vector (Promega, Madison, WI, USA).
Alignment of the MPF2 amino acid sequences was conducted using the program ClustalX 1.83 (Thompson, 1997), and then visualized for minor corrections in MacClade 4.08 (Maddison and Maddison, 2000). Neighbor-joining trees were generated by PAUP* 4.0b10 (Swofford, 2002) and SQUA from Antirrhinum majus was used as the outgroup. (2) Expression of MuMPF2
In order to examine the expression pattern of MPF2 homologues in M. pubescens, total RNAs were extracted from leaves, enlarged sepals, small sepals, petals plus stamens, carpels, and fruits respectively, by the methods described above. The specific primers used to amplify MPF2 homologues of M. pubescens were MuMPF2-F
(5’-CAGGTGACTTTCTCCAAGAGGAGACG) and MuMPF2-R (5’-
ATTGAAGCAGAGCTGGCTGCAACC). The PCR reaction was 95ºC for 15min; 95ºC for 45 sec, 65ºC for 45 sec, 68ºC for 45 sec (35 cycles); 68ºC for 7 min and then stopped at 4ºC. In addition, we also amplify the UBQ gene of M. pubescens to serve as a positive control (Primers: MuUBQ10F-5’GGAGGT GGAGAGTTCGGATA3’ and
MuUBQ10R-5’CGACAAATAAACACCACCACC3’).
5. Southern hybridization
Plant materials of M. pubescens have been collected from the populations at Shu-Mei-Pin (Taipei Co.) and Kenting (Shu-Mei-Pingtung Co.). Total DNA was isolated by 2X CTAB method (Doyle & Doyle, 1987), and digested with BamHI, HindIII and EcoRI. After electrophoresed on 0.9% agarose gels, DNA was transferred to nylon membranes using the alkaline transfer method (Sambrook et al, 1989). C-terminal fragments of all five floral ABE genes about 0.4kb were labeled with dig-dUTP by PCR. Hybridization with high stringency wash and signal detection were followed the protocols suggested by Dig application manual (Roche).
IV. Results and Discussion 1. Full length cDNA construction
We have obtained full-length cDNA sequences of all five genes (1 A, 3 B, and 1 C class genes), and have them cloned into pGEM-T vectors. No new homologues were found during 5' RACE or full-length cDNA PCR amplification after examining the sequence alignments.
2. Yeast-2-hybrid for previously identified MADS-box genes
We have obtained full-length cDNA clones of all five genes in either pAD-GAL4 or pBD-GAL4 vectors and confirmed the sequences were not modified during our preparation by re-sequencing. All plasmids were transformed separately into yeasts and examined by filter lift assay. There is no -galactosidase activities detected in these transformants (data not shown), which indicates no auto-activation in all of our constructs. Different pairs of plasmids were then co-transformed into yeasts and grown on selection media using HIS3 as reporter genes to investigate the interactions. A typical result of yeast-2-hybrid is shown in Fig. 1. The result of the yeast-two-hybrid experiment is summarized in Table 2.
The result reveals interactions between the two B-class proteins, MupGLO and MupDEF, and between the A- and E-class proteins, MupAP1 and MupSEP based on the confirmation of domains (AD vs. BD) swapping. For pairwise comparison, no homodimer was detected in our studies except for MupSEP. In addition, our data show all proteins including LaminC, a human protein for negative control, could interact with MupSEP while it fused to the GAL4 DNA binding domain. This suggests the MupSEP might interact directly with GAL4 activation domain, therefore, the positive result for the BD-MupSEP constructs might be an artifact. More detailed experiment is needed for solving this problem. Another interesting result is that we did not find the interaction between MupTM6 and other B-class proteins, although TM6 homologue is generally thought to be a partial redundant copy of DEF homologue. A TM6 homolog, PHTM6, from Petunia hybrida, is shown to involve in stamen, but not petal formation since it can only complement the phdef mutation in stamen development but not in petal development (Vandenbussche et al. 2004). This
indicates MupTM6 might be involved in a new function distinct from the ordinary B function. PHTM6 has been shown to interact with PHGLO2, but only weakly interact with another
GLO homolog, PHGLO1 (Vandenbussche et al. 2004). It is possible that MupTM6 needs additional factors to stabilize its dimerization, a yeast-three-hybridization may help to elucidate this possibility.
Fig. 1. A representation of the yeast-2-hybrid result, showing the positive interaction of AP1 (fused to BD) and SEP (fused to AD).
Table 2. Protein-protein interactions between ABE genes by Y2H.
Bait
MupAP1 MupDEF
Prey
MupGLO MupTM6 MupSEP LaminC
MupAP1 ○ --
MupDEF ○ --
MupGLO ○ --
MupTM6 --
MupSEP ○ ○ ○ ○ ○ ○
3. cDNA library construction and screening
choose two kinds of plant tissues, flower buds and mature enlarged calyx, to construct the cDNA library, because the floral organ identity genes has known not only expressed at early stage but continually expressed to maintain the organ identity. The entire open-reading frame of two B-class genes was cloned into the yeast vector pBD-GAL4 to produce a protein fusion with the GAL4 DNA-binding domain as the baits for yeast two-hybrid screening. In last year projects, we have demonstrated this fusion was workable and didn’t affect the interaction between MupDEF and MupGLO. Based on our previous data, the pairs, MupDEF and MupGLO, could be a good positive control to confirm the process of the library screening.
A lambda phage library of flower buds and enlarge calyx cDNA was constructed in the HybriZAP GAL4-activation-domain vector, and subsequently converted to a phagemid library in the yeast vector pAD-GAL4. For the library screening, the bait, pBD-MupDEF or pBD-MupTM6, and the phagemid library fusion plasmids were co-transformed into the yeast strain YRG2 containing two reporter genes, HIS3 and lacZ. To estimate how many
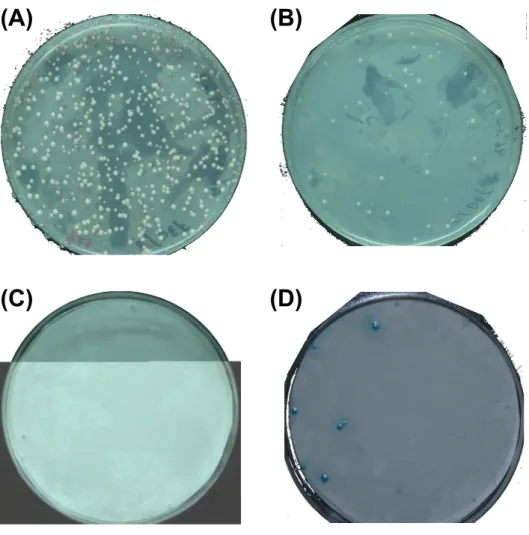
transformants were screened, part or transformants were plated on SD plate without Trp and Leu (Fig. 3A). About 1,000 clones have grown on SD plates without His, Trp, and Leu from ~ 40,000 transformants (Fig. 3B). To distinguish between HIS3 leaky expression and specifically interactions, the expression of the second reporter gene (lacZ) was detected by the filter lift assay. Unfortunately, no positive transformant was detected after the double confirming (Fig. 3C) and positive control shown in Fig. 3D.
4. Characterization of MPF homolog
(1) Cloning and identification of MPF homologues in M. pubescens
We successfully cloned MPF2 homologues in M. pubescens (MuMPF2); besides, we also isolated homologues from the other two subclades of STMADS11 gene family,
STMADS11 and SVP; here we named them MuMPF1 and MuSVP, respectively. Forty clones
were screened for STMADS11-like genes in M. pubescens and seventeen of those clones had correct insert size and were sequenced. Based on the BLAST analysis, 15 clones are the members of STMADS11 superclade. Among them, three copies out of seven clones are
MupMPF2, seven copies from seven clones are MuSVP and only one clone is MupMPF1.
The phylogenetic analysis further demonstrates that MupMPF2, MupMPF1 and MuSVP fall into their corresponding subclades within the STMADS11 superclade (Figure 4).
Fig. 3. A typical results of the yeast-2-hybrid library screening, showing the transformants grown on the SD plate without Trp and Leu (A), or without Trp, Leu, and His (B). The selective clones were subsequently detected the activity of β-galactosidase by filter list assay (C), and transformants of pAD-MupGLO/ pBD-MupDEF were positive control (D).
(A)
(B)
(D)
(C)
Fig. 4. Result of phylogenetic analysis of MPF homologues using NJ criteria, including three found in this study (marked as red). Numbers above the branches are bootstrap values from 1000 replicates.
(2) Expression pattern of MupMPF2 in M. pubescens
The expression pattern of MuMPF2 was examined by RT-PCR on leaves and different parts of the flowers. The result showed that MuMPF2 is expressed in all tissues except for the fruits (Fig. 5). It has highest expression level in the leaves, mediate for the sepals, and lowest in the carpel and fruits. This pattern is similar the expression pattern of
MPF2 in Physalis, which is predominately expressed in leaves. Although MuMPF2 is also
expressed in the enlarged sepals, whether or not it contributes to the inflated calyx is to be examined.
Fig. 5. Expression pattern of MuMPF2 by RT-PCR. L: Leaves; ES: Enlarged sepal; SS: Small sepal; PS: Petal and stamen; C: Carpel; F: Fruit
5. Southern hybridization analysis for floral ABE genes
In the flowering plants, there are several duplication events of floral organ identity genes to recruit new developmental plan (Irish, 2003). In M. pubescens the numbers of these genes should be confirmed, only the single copy of cDNA was found in our previous studies though. To distinguish these genes in the same MADS-box family, the divergent C-terminal fragments of these genes were used to be the probe in southern analysis (Fig. 6). The blots shows only one major band that been found among different restriction enzyme digestion for
MupAP1, MupDEF, and MupSEP. However, the multiple bands for MupGLO and MupTM6
means there might be more copies in M. pubescens genomes. The roles of these possible redundant genes should be clarified in future studies.
In our lab previous survey, some populations of M. pubescens in north Taiwan flowers without the enlarged petaloid calyx. Although the cDNA sequences of these five petal organ identity genes are completely identical between north and south population, the patterns of southern blot shows the divergence between the two populations (Fig. 6). The mechanism of the loss of the petaloid structure in north population is still unknown. More detail comparisons might help us to understand the formation of the petaloid structure in
Mussaenda species.
MPF2
UBQ
Fig. 6. Southern blot analysis for floral A (AP1, referring to MupAP1), B (DEF, GLO, TM6, referring to MupDEF, MupGLO, MupTM6), and E (SEP, referring to MupSEP) genes. Marker sizes are marked at the right of the gel. (N: north population, S: south population, B:
BamHI, E: EcoRI, and H: HindIII)
V. References
Albert, V. A., M. H. G. Gustafsson, and L. D. Laurenzio. 1998. Ontogenetic systematics, molecular developmental genetics, and the angiosperm petal. Pages 349-374 in Molecular systematics of plants II: DNA sequencing (D. E. Soltis, P. S. Soltis, and J. J. Doyle, eds.). Kluwer Academic Publishers, Boston.
Becker, A. and Theissen, G. 2003. The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Molecular Phylogenetics and Evolution 29(3): 464-489.
Baum, D. A. 1998. The evolution of plant development. Current Opinion in Plant Biology 1: 79-86.
Baum, D. A., and B. A. Whitlock. 1999. Plant development: Genetic clues to petal evolution.
Current Biology 9: R525-R527.
Bowman, J. L. 1997. Evolutionary conservation of angiosperm flower development at the molecular and genetic levels. Journal of Biosciences 22: 515-527.
Chang, S., J. Puryear, and J. Chirney. 1993. A simple and efficient method for isolating RNA from pine trees. Plant Molecular Biology Reporter 11: 113-116.
Doyle, J.J. and J.L. Doyle. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19: 11-15.
He, C.Y., Munster, T., and Saedler, H. 2004. On the origin of floral morphological novelties. Febs
Letters 567(1): 147-151.
He, C. Y., and H. Saedler. 2005. Heterotopic expression of MPF2 if the key to the evolution of the Chinese lantern of Physalis, a morphological novelty in Solanaceae. PNAS 102: 5779-5784.
Hecht, V., F. Forcher, C. Ferrandiz, R. Macknight, C. Navarro, J. Mprin, M. E. Vardy, N. Ellis, J. P. Beltran, C. Rameau, and J. L. Weller. 2005. Conservation of Arabidopsis flowering genes in models legumes. Plant Physiology. 137: 1420-1434.
Honma, T. and K. Goto. 2001. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 409: 525-529.
Irish, V.F. 2003 The evolution of floral homeotic gene function. BioEssays 25: 637-646. Maddison, D. R., and W. P. Maddison. 2000. MacClade 4: Analysis of phylogeny and
character evolution. Sinauer Associates, Sunderland, MA, USA.
Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular Cloning: A Laboratory Maanual. 2nd Edition. Cold Springer Harbor Lab Press, NY, USA.
Swofford, D. L. 2002. Phylogenetic analysis using parsimony (*and other methods). Version 4. Sinauer Associates Inc., Sunderland, MA, USA.
Theissen, G. 2001. Development of floral organ identity: stories from the MADS house.
Current Opinion in Plant Biology 4: 75-85.
Theissen, G. and H. Saedler. 2001. Floral quartets. Nature 409: 469-471.
Thomopson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G. Higgins. 1997. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research 25: 4876-4882.
Vandenbussche, M., Zethof, J., Royaert, S., Weterings, K., and Gerats, T. 2004. The duplicated B-class heterodimer model: Whorl-specific effects and complex genetic interactions in Petunia
VI. Self-evaluation
The interactions between the proteins on previously identified floral homeotic genes were evaluated. We also constructed these genes for the yeast 2 hybrid screening. However, the screening process has had a lot of troubles. The major problem is mostly from the plausibility of constructing an authentic cDNA library and many unexpected obstacles during cloning process. Although we have tried very hard to make the floral cDNA library from fresh materials, the results still indicate the quality of cDNA might not be as good as expected.
We also encountered problems in seed germination and seedling transplantation. The quality of seeds we harvested is not consistent. This might be due to environmental factors possessed to the fruits. This is the main challenge on conducting experiments for wild plants in the field. It is not like to controlled lab environment, so that the experiment may have unexpected troubles.