禽流感病毒 H6N1 亞型基因定序及其與病原性的關係
計畫類別: 整合型計畫 計畫編號: NSC93-2317-B-002-002- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學獸醫學系暨研究所 計畫主持人: 王金和 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 9 月 12 日
行政院國家科學委員會專題研究計畫 成果報告
禽流感病毒 H6N1 亞型基因定序及其與病原性的關係
計畫類別: 整合型計畫 計畫編號: NSC 93-2317-B-002-002 執行期間: 93 年 08 月 01 日 至 94 年 07 月 31 日 執行單位: 國立臺灣大學獸醫學系暨研究所 計畫主持人: 王金和 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 9 月 11 日
流感病毒預防及控制-禽流感病毒 H6N1 亞型基因定序及其與病原性
的關係
Ching-Ho WANG
Department of Veterinary Medicine, National Taiwan University, Taipei 10617, Taiwan
Department of Veterinary Medicine, National Taiwan University, Taipei 10617, Taiwan
No 1, Sec 4, Roosevelt Road, Taipei 10617, Taiwan Tel: 02 2369 0628
Fax: 02 2363 1542
E-mail: [email protected]
ABSTRACT
The current concern of influenza is that avian influenza viruses transmit or provide to humans. Although the genomic information of human influenza A viruses is increasing, little of this type of data is available for viruses circulating in poultry farms. This study presented the genetic characterization of 9 H6N1 viruses isolated from chicken farms from 2000 to 2003. The whole complete eight genes of 3 avian isolates and the six genes of the rest 6 avian isolates were sequenced and compared. The phylogenetic trees of those genes demonstrate that influenza H6N1 viruses circulating in chickens in Taiwan are quite unique and different from those of other countries. *Corresponding author TEL: 02-23680628, Fax: 02-23631542, E-mail:
Key words: Avian influenza, H6N1, gene analysis
INTRODUCTION
There are 15 hemagglutinin (HA) and 9 neuraminidase (NA) subtypes of influenza A viruses. Most of the subtypes have been recognized in avian species, especial wild birds [6], but only three HAs and two NAs (H1, H2, H3, N1, and N2) have been recognized in human influenza viruses. The appearance of new subtypes of influenza A viruses can lead to pandemics in humans because humans have no
antibodies against the new subtype viruses. The genes responsible for the emergence of a new virus are thought to have been introduced from avian species [7,15]. Avian influenza viruses, H6N1 and H9N2 have been enzootic in Asia during the past decade and have been isolated from different types of terrestrial poultry
worldwide [2]. The H6N1 associated with H9N2 have been involved in the genesis of the pathogenic H5N1 influenza viruses of 1997, which caused the death of human beings [4].
The first H6N1 avian influenza outbreak in Taiwan was reported in 1972 [8]. Since then, the surveillance has been continuous performed [3]. Recently, more and
more H6N1 viruses were isolated from the field around Taiwan [14]. Some of them caused mortality and egg production drop in chickens. Several H6N1 avian influenza viruses were isolated from chickens in this laboratory during 2000 and 2003. The present study was to investigate the epidemiology of those viruses and whether their genes emerge to human or other viruses.
MATERIALS AND METHODS Viruses.
The viruses used in this study were isolated from the field in Taiwan during 2000 and 2003 (Table 1). Viruses were propagated in 11-day-old SPF chicken embryos (Animal Health Research Institute, Chidin, Taiwan). The aliquots were kept in -70 C for gene analysis.
Cloning and sequencing of virus gene fragments.
The eight gene fragments of both strains were extracted with TriSolution
(Genmedika Biotechnol. Corp., Taipei) and amplified with specific primers by reverse transcription–polymerase chain reaction (RT-PCR) for different fragments (Table 2-9). Different primers were used for the same gene fragment from different virus isolates. Because some primers didn’t amplify some isolates, we designed new primers for those isolates.
The RT-PCR products were cloned into yT&A cloning kit (Yeastern Biotech. Co., Taipei) and sequenced using ABI prism cycle sequencing kit (Perkin Elmer,
Banchburg, NJ). Forward and reverse sequences were performed.
Sequence analysis.
The nucleotide sequences of the AIV we isolated and the sequences we gathered from the GenBank were aligned using the Vector NTI Suite 6 software (InforMax, Inc., Bethesda, MD). Phylogenetic trees were generated based on the alignment of the complete or partial nucleotide sequences of each viral gene by using the Neighbor Joining method of Saitou and Nei. The regions of each gene used in analysis included: PB2, nt position 1-2277; PB1, 1-2271; PA, 1-2148; HA, 1-1701; NP, 7-1479; NA, 28-1402 ; M, 7-909 and NS, 1-836. Accession numbers of virus sequences from the GenBank used in this study were as the followings.
PB2
Qa/HK/SF595/00(H6N1) AJ410499, Ph/HK/NT261/00(H6N1) AJ410500, Ck/Taiwan/7-5/99(H6N1) AF255624, Qa/HK/1721-20/99(H6N1) AJ410496, Ph/HK/SH39/99(H6N1) AJ410495, Teal/HK/W312/97(H6N1) AF250476,
Bird/Thailand/3.1/04(H5N1) AY651715, Ck/Guangdong/174/04(H5N1) AY609309, Ck/Jilin/9/04(H5N1) AY653193, Ck/Vietnam/C58/04(H5N1) AY818127,
Ck/HK/31.4/02(H5N1) AY576386, Gs/Guangdong/1/96(H5N1) AF144300, Vietnam/1203/04(H5N1) AY818126, Vietnam/3062/04(H5N1) AY651721, HK/212/03(H5N1) AY576380, HK/156/97(H5N1) AF036363,
HK/1774/99(H3N2) AJ293920, Taiwan/3/71(H3N2) AY210146, Taiwan/2/70(H3N2) AY210144, Taiwan/1/69(H3N2) AY210141, HK/1/68(H3N2) AF348170,
HK/427/98(H1N1) AF258525, HK/470/97(H1N1) AF258524, Fiji/15899/83(H1N1) AJ564805
Qa/HK/SF595/00(H6N1) AJ410506, Ph/HK/NT261/00(H6N1) AJ410507, Ck/Taiwan/7-5/99(H6N1) AF262210, Qa/HK/1721-20/99(H6N1) AJ410503, Ph/HK/SH39/99(H6N1) AJ410502, Teal/HK/W312/97(H6N1) AF250477,
Bird/Thailand/3.1/04(H5N1) AY651661, Ck/Guangdong/174/04(H5N1) AY609310, Ck/Jilin/9/04(H5N1) AY653199, Ck/Vietnam/C58/04(H5N1) AY818130,
Ck/HK/31.4/02(H5N1) AY576398, Gs/Guangdong/1/96(H5N1) AF144301, Vienam/1203/04(H5N1) AY818129, Vietnam/3062/04(H5N1) AY651667, HK/212/03(H5N1) AY576392, HK/156/97(H5N1) AF036362,
HK/1774/99(H3N2) AJ293921, Taiwan/3/71(H3N2) AY210279, Taiwan/2/70(H3N2) AY210282, Taiwan/1/69(H3N2) AY210276, HK/1/68(H3N2) AF348172,
HK/427/98(H1N1) AF258526, HK/470/97(H1N1)AF258527, Fiji/15899/83(H1N1) AJ564807
PA
Qa/HK/SF595/00(H6N1) AJ410515, Ph/HK/NT261/00(H6N1) AJ410516, Ck/Taiwan/7-5/99(H6N1) AF262211, Qa/HK/1721-20/99(H6N1) AJ410512, Ph/HK/SH39/99(H6N1) AJ410511, Teal/HK/W312/97(H6N1) AF250478,
Bird/Thailand/3.1/04(H5N1) AY651607, Ck/Guangdong/174/04(H5N1) AY609311, Ck/Jilin/9/04(H5N1) AY653198, Ck/Vietnam/C58/04(H5N1) AY818133,
Ck/HK/31.4/02(H5N1) AY576410, Gs/Guangdong/1/96(H5N1) AF144302, Vietnam/1203/04(H5N1) AY818132, Vietnam/3062/04(H5N1) AY651612, HK/212/03(H5N1) AY576404, HK/156/97(H5N1) AJ289874,
HK/1774/99(H3N2) AJ293922, Taiwan/3/71(H3N2) AY210202, Taiwan/2/70(H3N2) AY210199, Taiwan/1/69(H3N2) AY210198, HK/1/68(H3N2) AF348174,
HK/427/98(H1N1) AF258519, HK/470/97(H1N1)AF258518, Fiji/15899/83(H1N1) AJ605762
HA
Dk/Hainan/6/04(H6N2) AY773907, Dk/Korea/S17/03(H6N1) AY862613, Qa/HK/SF595/00(H6N1) AJ410523, Ph/HK/NT261/00(H6N1) AJ410524, CK/Taiwan/7-5/99(H6N1) AF310983, Qa/HK/1721-20/99(H6N1) AJ410520, Ph/HK/SH39/99(H6N1) AJ410519, Dk/HK/3461/99(H6N1) AJ410537, Ck/Taiwan/na3/98(H6N1) AF310984, Teal/HK/W312/97(H6N1) AF250479, Pintail/Alberta/179/93(H6N1) AY633332, Dk/HK/202/77(H6N1) AJ410544, NP
Qa/HK/SF595/00(H6N1) AJ410552, Chukka/HK/NT261/00(H6N1) AJ410553, CK/Taiwan/7-5/99(H6N1) AF261750, Qa/HK/1721-20/99(H6N1) AJ410549, Ph/HK/SH39/99(H6N1) AJ410548, Teal/HK/W312/97(H6N1) AF250480,
Bird/Thailand/3.1/04(H5N1) AY651495, Ck/Guangdong/174/04(H5N1) AY609313, Ck/Jilin/9/04(H5N1) AY653196, Ck/Vietnam/C58/04(H5N1) AY818139,
Ck/HK/31.4/02(H5N1) AY575911, Gs/Guangdong/1/96(H5N1) AF144303, Vietnam/1203/04(H5N1) AY818138, Vietnam/3062/04(H5N1) AY651501, HK/212/03(H5N1) AY575905, HK/156/97(H5N1) AF036359,
HK/1774/99(H3N2) AJ293924, Taiwan/3/71(H3N2) AY210230, Taiwan/2/70(H3N2) AY210227, Taiwan/1/69(H3N2) AY210225, HK/1/68(H3N2) AF348180,
HK/427/98(H1N1) AF258516, HK/470/97(H1N1) AF258517, Fiji/15899/83(H1N1) AJ628066
Qa/HK/SF595/00(H6N1) AJ410561, Ph/HK/NT261/00(H6N1) AJ410562, CK/Taiwan/7-5/99(H6N1) AF208598, Qa/HK/1721-20/99(H6N1) AJ410558, Ph/HK/SH39/99(H6N1) AJ410557, Teal/HK/W312/97(H6N1) AF250481,
Bird/Thailand/3.1/04(H5N1) AY651441, Ck/Guangdong/174/04(H5N1) AY609314, Ck/Jilin/9/04(H5N1) AY653195, Ck/Vietnam/C58/04(H5N1) AY818142,
Ck/HK/31.4/02(H5N1) AY575887, Gs/Guangdong/1/96(H5N1) AF144304, Vietnam/1203/04(H5N1) AY818141, Vietnam/3062/04(H5N1) AY651448, HK/212/03(H5N1) AY575881, HK/156/97(H5N1) AF036357,
HK/1774/99(H3N2) AJ293923, Taiwan/3/71(H3N2) AY210131, Taiwan/2/70(H3N2) AY210126, Taiwan/1/69(H3N2) AY210122, HK/1/68(H3N2) AF348184
Fiji/15899/83A(H1N1) J006954, Dk/Hainan/4/04(H6N2) NA AY706954 MP
Qa/HK/SF595/00(H6N1) AJ410572, Ph/HK/NT261/00(H6N1) AJ410573, CK/Taiwan/7-5/99(H6N1) AF262213, Qa/HK/1721-20/99(H6N1) AJ410569, Ph/HK/SH39/99(H6N1) AJ410568, Teal/HK/W312/97(H6N1) AF250482,
Bird/Thailand/3.1/04(H5N1) AY651384, Ck/Guangdong/174/04(H5N1) AY609315, Ck/Jilin/9/04(H5N1) AY653194, Ck/Vietnam/C58/04(H5N1) AY818145,
Ck/HK/31.4/02(H5N1) AY575899, Gs/Guangdong/1/96(H5N1) AF144306, Vietnam/1203/04(H5N1) AY818144, Vietnam/3062/04(H5N1) AY651390, HK/212/03(H5N1) AY575893, HK/156/97(H5N1)AF036358,
HK/1774/99(H3N2) AJ293925, Taiwan/3/71(H3N2) AY210263, Taiwan/2/70(H3N2) AY210260, Taiwan/1/69(H3N2) AY210256, HK/1/68(H3N2) AF348188,
HK/427/98(H1N1) AF258523, HK/470/97(H1N1) AF258522, Fiji/15899/83(H1N1) AJ298947
NS
Dk/Korea/S17/03(H6N1) AY862677, Qa/HK/SF595/00(H6N1) AJ410581, Ph/HK/NT261/00(H6N1) AJ410582, Ck/Taiwan/7-5/99(H6N1) AF262212, Qa/HK/1721-20/99(H6N1) AJ410578, Ph/HK/SH39/99(H6N1) AJ410577, Teal/HK/W312/97(H6N1) AF 250483, Dk/HK/3461/99(H6N1) AJ410594,
Bird/Thailand/3.1/04(H5N1) AY651550, Ck/Guangdong/174/04(H5N1) AY609316, Ck/Jilin/9/04(H5N1) AY653197, Ck/Vietnam/C58/04(H5N1) AY818148,
Ck/HK/31.4/02(H5N1) AY576374, Gs/Guangdong/1/96(H5N1) AF144307, Vietnam/1203/04(H5N1) AY818147, Vietnam/3062/04(H5N1) AY651555, HK/212/03(H5N1) AY576368,HK/156/97(H5N1) AF036360,
HK/1774/99(H3N2) AJ293941,Taiwan/3/71(H3N2) AY210307, Taiwan/2/70(H3N2) AY210306, Taiwan/1/69(H3N2) AY210302, HK/1/68(H3N2) AF348198,
HK/427/98(H1N1) AF258521, HK/470/97(H1N1) AF258520, Fiji/15899/83(H1N1) AJ298950
RESULTS Clinical cases and viruses.
Nine AIVs were isolated form 9 flocks, including broiler breeder farms, layer breeder farms, and a broiler farm (Table 1). The onsets lasted for 5 to 7 days and then recovered. The weekly mortality ranged from 0.3% to 6.0%. The egg production decreased 0% to 33%.
Hemorrhage was found on the laryngeal, pharyngeal and tracheal mucosa. Air sac showed cloudy and thickening. Papillary hemorrhage was noted in the
proventriculus. Kidney was swollen and had urate deposition. Escherichia coli was always isolated from the infected chickens.
Genetic sequences.
The PB2 full-length gene segments of 2838N/00, 2838V/00, and 2937/01 were determined. RNA segment 1 contained 2341 nucleotides and coded for a 759-a.a. PB2 protein from MERIK to RMAIN. The nucleotide percent identity among Taiwan H6N1 strains were from 95.4% to 99.6%.
The PB1 full-length gene segments of 2838N/00, 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03 and 3153/03 were determined. This RNA segment 2 contained 2341 nucleotides and coded for a 757-a.a. PB1 protein from MDVNP to LRRQK. The nucleotide percent identity among Taiwan H6N1 strains were from 93.5 to 99.8%.
The PA full-length gene segment of 2838N/00, 2838V/00, and 2937/01 were determined. This RNA segment 3 contained 2233 nucleotides and coded for a 716-a.a. PA protein from MEDFV to THALK. The nucleotide percent identity among Taiwan H6N1 strains were from 98 to 99.6%.
The HA full-length gene segments of 2829/00, 2838N/00, 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03 and 3153/03 were determined. This RNA segment 4 contained 1776 nucleotides. The main HA protein was 551 in length with 329 HA1 and 222 HA2. The mature HA1 was from DKICI to QIETR. The nucleotide percent identity among Taiwan H6N1 strains were from 93.7 to 99.6%. The
connecting sequences consisted of a single R, IETR↓GIFG in the present isolates (arrow means cleavage site) except 2838N/00, which was IETR↓GILR. The sequence after the cleavage site in most influenza viruses is GLF. Surprisingly, the related sequence in the present H6N1 HA sequence was GIFG, the same as A/chicken/7-5/99 (accession number: AF310983 in the GenBank). In the critical HA receptor-binding site, the 228-related position (226 in H6) of the HA from the present isolates was S, like the human-adapted H3 or H2 strains (225-203 of H3 numbering, G-Q-R-S-RI). The NP full-length gene segments of 2829/00, 2838N/00, 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03 and 3153/03 were determined. This RNA segment 5 contained 1487 nucleotides and was coded for protein of 495 a.a., begin from SQGTK to AEEYD. The nucleotide percent identity among Taiwan H6N1 strains were from 95.8 to 99.3 %. The anchor residue of the cytotoxic T lymphocyte epitope SRYWAIRTR (383 to 391) was found, which could be recognized by specific cytotoxic T lymphocytes [13].
The NA full-length gene segments of 2829/00, 2838N/00, 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03 and 3153/03 were determined. This RNA segment 6 contained 1375 nucleotides and was coded for protein of 456 a.a., begin from MDVNP to LGRQK. The nucleotide percent identity among Taiwan H6N1 strains were from 93.7 to 99.7 %. The glycosylation of N at 144 a.a. conserved from the functional requirement was also found[11]. Some strains showed 12 a.a. deletion in the NA stalk and some not.
The M full-length gene segments of 2829/00, 2838N/00, 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03 and 3153/03 were determined. This RNA segment 7 contained 903 nucleotides. The encoded polypeptide divided into M1 (250 a.a.), begin from MDVNP to LGRQK and M2 (97 a.a.). The nucleotide percent identity among Taiwan H6N1 strains were from 95.6 to 99.8 %. RNA segment 7
coded for 252 a.a. of M1 and 97 a.a. of M2 proteins, respectively. The a.a. positions 27 to 30 of M2 protein associated with resistance was VIAS. The a.a. change from I to T or S at position 27 and from A to T or S at position 30 of M2 are associated with amantadine resistance [1]. The position 30 were S, indicating those strains might resistant to amantadine.
The NS full-length gene segments of 2829/00, 2838N/00, 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03 and 3153/03 were determined. This RNA segment 8 contained 890 nucleotides and was coded for protein of 230 a.a. (NS), begin from MDSNT to IESEV. The nucleotide percent identity among Taiwan H6N1 strains were from 94 to 99.9 %. RNA segment 8 coded for 230 a.a. of NS1 and 121 a.a. of NS2 proteins, respectively. The position 92 of NS1 protein was D, which was
sensitive to cytokine and less virulent than highly pathogenic H5N1 [12].
Phylogenetic data.
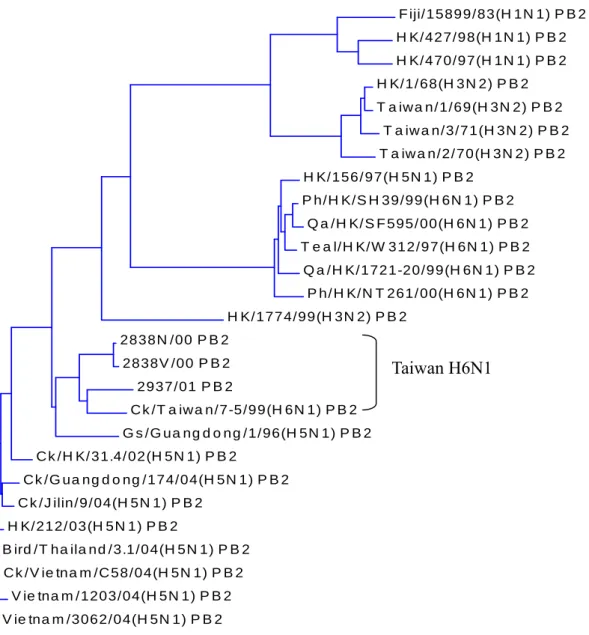
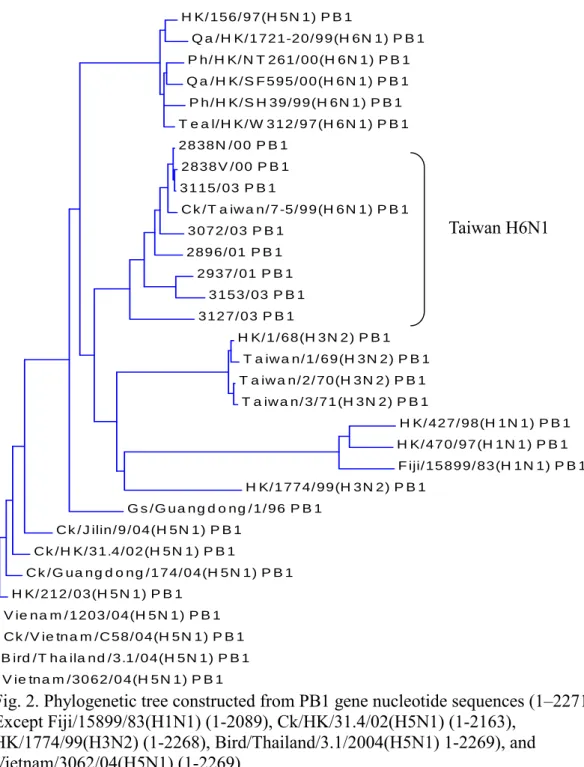
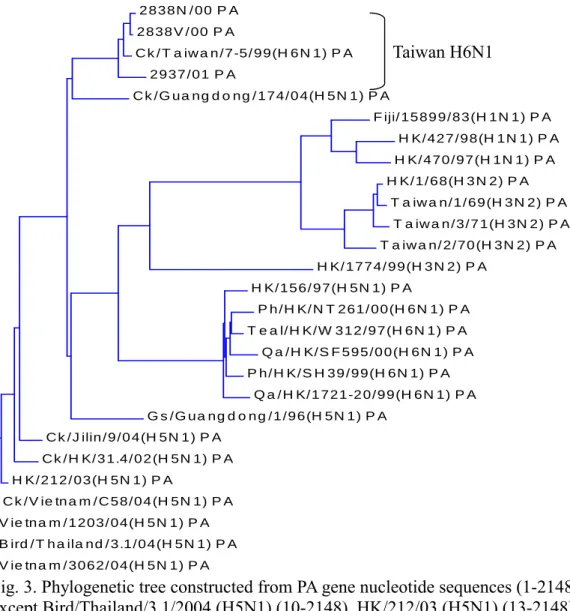
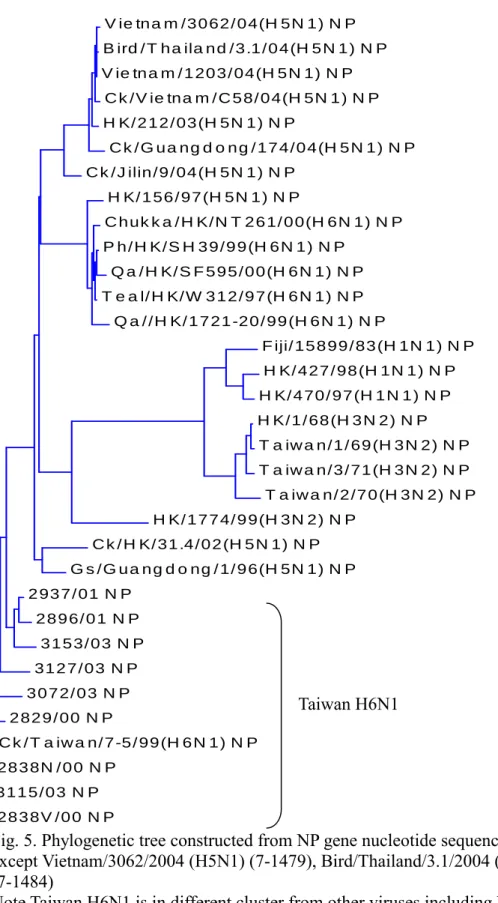
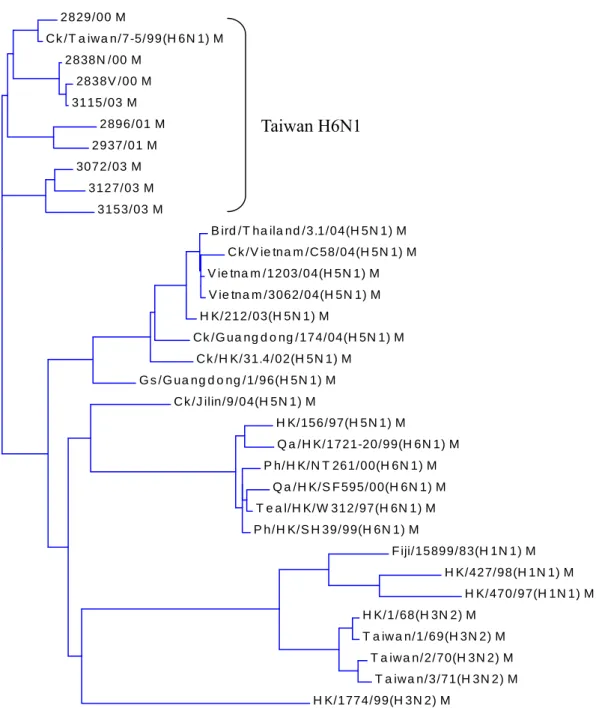
All of the H6N1 influenza viruses isolated from Taiwan were clustered together and different from viruses of other countries (Fig. 1,2,3,4,5,6,7,8). They could be subgrouped into two different clades from all the eight gene fragments (Fig. 1,2,3,4,5,6,7,8), with 3115/03 and 2838V/00 in a clade and the others in another clade.
DISCUSSION
Consistent with accepted dogma, genes from influenza viruses isolated from birds are in evolutionary stasis and the majority of the genes clustered together are separated from viruses from other areas. Interestingly, Taiwan AIVs are quite unique because they are clustered together and separate from AIVs of other areas. All of the isolates are clustered in spite of their origin. From the phylogenetic trees of PB1, HA, NP, NA, M, and NS, 2838N/00 and 2838V/00 are clustered into the same clade as 3115/03 in spite of their origin was quite different. The percentage identity between 2838N/00 and 3115/03 were PB1: 99.7%, HA:99.1%, NP:99.3%, NA:97.2%, M:99.7%, and NS:99.9%. Isolate 2838/00 wasfrom a broiler breeder in Yilan and 3115/03 was from a broiler farm in Taoyen. The broilers were not from that breeder farm. The similar sequences indicated that AIVs in Taiwan are circulated over the all island but not restricted to a specific area.
Another unique finding in this study is the presence of AIVs spread by wind from clinical epidemiologic investigation. Those viruses infected in a flock and then transmitted to adjacent flocks in spite of restricted biosafety management even the movement of workers and equipments between different premises were restricted.
The host range of influenza A viruses is associated with differences in the specificity of the HA to attach sialic acid-containing receptors on susceptible cells [10]. HAs of avian influenza viruses preferentially recognize sialic acid linked to galactose by α2,3 linkage, whereas HAs of human influenza viruses preferentially recognize sialic acidα2,6Gal linkages [9]. Residues 226 and 228 in the HAs of the H2 and H3 subtypes of human influenza viruses may be important in determining receptor specificity [5]. In all H6N1, the HA posses a Q residue at the position 226 and an S residue at position 228. This combination of amino acids binds preferentially to sialic acidα2,3 if there is an additional substitution of Q226L in; it may enable this virus to recognize the mammalian receptor and significantly increase the potential of the virus to infect mammals [5]. The potential of human transmission of the present H6N1 viruses should be further studies.
ACKNOWLEDGMENTS
These studies were supported by a grant from National Science Council (NSC93-2317-B-002-002)
REFERENCES
1. Bean WJ, Threlkeld SC, Webster RG. Biological potential of amnatadine-resistant influenza A virus in an avian model. J Infect Dis 159: 1050-1056. 1989.
2. Cameron KR, Gregory V, Banks J, Brown IH, Alexander DJ, Hay AJ. Lin YP. H9N2 subtype influenza A viruses in poultry in Pakistan are closely related to the H9N2 viruses responsible for human infection in Hong Kong. Virology 278:36-41. 2000.
3. Chan CH, Shieh HK, Lu YS, Lee YL, Lin DF, Liao YK. Study on avian influenza in Taiwan ROC. Taiwan J Vet Med Anim Husb 53: 75-88, 1989.
4. Chin PS, Hoffmann E, Webby R, Webster RG, Guan Y, Peiris M, Shortridge KF. Molecular evolution of H6 influenza viruses from poultry in southeastern
China:prevalence of H6N1 influenza viruses possessing seven A/hong Kong/156/97 (H5N1)-like genes in poultry. J Virol 76:507-516. 2002.
5. Connor RJ, Kawaoka Y, Webster RG, Paulson JC. Receptor specificity in human, avian , and equine H2 and H3 influenza virus isolates. Virology 205: 17-23, 1994 6. Hinshaw VS, Webster RG, Rodriguez RJ. Influenza A viruses: combinations of hemagglutinin and neuraminidase subtypes isolated from animals and other sources. Arch Virol 67:191-201, 1981.
7. Kawaoka Y, Krauss S, Webster RG. Avian-to-human transmission of the PB1 gene of influenza A viruses in the 1957 and 1968 pandemics. J Virol 63:4603-4608. 1989. 8. Lu YS, Sugimura T, Shieh HK, Lee YL, Jong MH. Isolation and identification of an influenza A virus in ducks in Taiwan. J Chin Soc Vet Med 11: 23-34, 1985.
9. Matrosovich MN, Gambaryan AS, Teneberg S, Piskarev VE, Yamnikova SS, Lvov DK, Robertson JS, Karlsson KA. Avian influenza A viruses differ from human viruses by recognition of sialyloligosaccharides and gangliosides and by a higher
conservation of the HA receptor-binding site. Virology 233: 224-234, 1997 10. Rogers GN, Paulson JC, Daniels RS, Skehel JJ, Wilson IA, Wiley DC. Single amino acid substitutions in influenza haemagglutinin change receptor binding specificity. Nature 304: 76-78, 1983
11. Saito T. Kawano K. Loss of glycosylation at Asn 144 alters the substrate preference of the N8 influenza A virus neuraminidase. J Vet Med Sci 59: 923-926. 1997.
12. SeoSH, Hoffmann E, Webster RG. Lethal H5N1 viruses escape host anti-viral cytokine responses. Nat Med 8: 950-954. 2002.
13. Voeten JTM, Bestebroer TM, Nieuwkoop NJ, Fouchier RAM, Osterhaus ADME, Rimmelzwaan GF. Antigenic Drift in the Influenza A Virus (H3N2) Nucleoprotein and Escape from Recognition by Cytotoxic T Lymphocytes. J Virol 74: 6800-6807. 2000.
14. Wang, CW, Wang CH. Experimental selection of virus derivatives with variations in virulence from a single low-pathogenicity H6N1 avian influenza virus field isolate. Avian Dis 47: 1416-1422, 2003.
15.Webster RG, Laver WG. The origin of pandemic influenza. Bull WHO 47:449-452, 1971.
Table 1. Histories of the flocks having H6N1 viruses by isolation.
Isolate Location Host Week
age
Eggb production
drop
Weekly
mortality deletionNA stalk C
2829/00 Yilan Broiler breeder 46 0% 0.3% No
2838/00a Yilan Broiler breeder 45 33% 3.8% Yes
2896/01 Miaoli Broiler breeder 45 10% 2.5% No
2937/01 Tainan Layer 4 NR 0.5% Yes
3072/03 Taichung Layer breeder 43 10% 2.8% Yes
3115/03 Taoyen Broiler 5 NR 6% Yes
3127/03 Taichung Broiler breeder 14 NR 0.5% Yes
3153/03 Yilan Broiler 5 NR 3% Yes
a. From which, 2838N/00 (nonvirulent) and 2838V/00 (virulent) were obtained. b. Usually, the egg production drop lasted for 5 days and recovered. ND: no record because those chickens are immature.
c. NA stalk deletion site: 120-159 (except 137-139)
Table 2. Primers used for PB2 gene cloning and sequencing
Gene Strains Primera Sequence Position
PB2 2838N/00 F: Ba-PB2-1 Ref. Hoffmann, 2000 -41 - -14
R: Ba-PB2-2341r Ref. Hoffmann, 2000 2329-2296
Internal primer: F: D PB2 F/F1 CAGACAGAGTGATGGTGT 257 - 274 R: D PB2 F/R2 GCACTGAGATCTGCATGG 467 - 450 F: D PB2 F/F2 GGTAAGGATGGTGGACAT 873 - 890 R: D PB2 F/R1 TCCTCAGGATAGCTGTTG 1126-1109 F: PB2-1286f TCGTAAACAGAGCAAACCAAAG 1259-1280 R: PB2-1458r TCCCATCTTACTAACTCTCACC 1452-1431
a. F: forward primer, R: reverse primer
Table 3. Primers used for PB1 gene cloning and sequencing
Gene Strains Primer Sequence Position
PB1 2838N/00 F: Ba-PB1-1 Ref. Hoffmann, 2000 -38 - -11
R:Ba-PB1-2341r Ref. Hoffmann, 2000 2332-2300
Internal primer: F: PB1-639f AGGGAAGAAGAAGCAGAGG 615-634 R: K PB1 F/R1 TTCTCACAGATGCTCCTC 794-777 F: PB1-990f AACATACATCACAAGAAGCCAG 966-987 R:PB1-1259r ATCATCATTCCAGGACTCAA 1226-1207 F: PB1r-630f ACAATATGATAAACAACGACCTTGG 1595-1619 R: PB1r-777r CCCACAGCCTCTTCAGCTCG 1741-1722
a. F: forward primer, R: reverse primer
b. The same primers were used for strain 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03,and 3153/03.
Table 4. Primers used for PA gene cloning and sequencing
Gene Strains Primer Sequence Position
PA 2838N/00 F: Bm-PA-1 Ref. Hoffmann, 2000 -38 - -10
R: Bm-PA-2233r Ref. Hoffmann, 2000 2224-2192
Internal primer:
F: PA-525f CAGGGCAAGAATCAAAACCAG 501-521
R: K PA F/R1 GCATCATACAGTGGTATAC 920-902
F: PA-1009f GGCATAAACCCCAATTACCTC 985-1005
R: PA-1155r CCATATTCTCACCAAGTGCCC 1123-1103
a. F: forward primer, R: reverse primer
Table 5. Primers used for HA gene cloning and sequencing
Gene Strains Primer Sequence Position
HA 2838N/00 F: P1(+)b 5’-GGAAAATGATTGCAATCATT -5 - 15
R: P4(-)b 5’-TTATATACATATCCTGCATTGCAT 1704-1681
Internal primer:
F: P2(+) CCTTGGTATGCATACAAATT 802 - 821
R: P3(-) GGTAAATTTGACTTGAAGAC 866 - 847
2829/00 F: Bm-HA-1 Ref. Hoffmann, 2000 -31 - -4
R: Bm-NS-890r Ref. Hoffmann, 2000 1745-1711 Internal primer: F: HA-400f AAATGGGATTTGCTACCCAGG 306 - 326 R: HA-600r GATTGGCTGGTTTCCAGTGTTG 570 - 549 F: HA-1206f CAAAATGAACACACAATTCGAAGC 1206-1229 R: HA-1398r CCCTTAGCTGCGATTTGACC 1417-1398
3115/03 F:Bm-HA-1 Ref. Hoffmann, 2000 -31 - -4
R: P4(-) Dr. Chang 1704-1681
Internal primer:
F: P2(+) CCTTGGTATGCATACAAATT 802 - 821
R: P3(-) GGTAAATTTGACTTGAAGAC 866 - 847
a. F: forward primer, R: reverse primer
b. From Dr. Chang PC of National ChungHsing Unviersity
c. The same primers used for 2838N/00 were used for strain 2838V/00.
d. The same primers used for 2829/00 were used for 2896/01, 2937/01, 3072/03, 3115/03, 3127/03, and 3153/03.
Table 6. Primers used for NP gene cloning and sequencing
Gene Strains Primer Sequence Position
NP 2838N/00 F: Bm-NP-1 Ref. Hoffmann, 2000 -59 - -31 R: Bm-NP-1565r Ref. Hoffmann, 2000 1593-1559 Internal primer: F: K- NP/F1 5’-TGATGCCACATACCAGAG 432-449 R: Bm-NP-756r 5’-TGTTTGGAATTTCCCTTTGAGG 755-734 R: K-NP/R1 5’-CTCTTGGKACCACTCTTG 1125-1108 F: NP2 5’-YAGATATTGGGCTATAAGAAC 1208-1228 2829/00 F: NPf 5’-TCTCAAGGCACCAAACG 7 - 23 R: NPr 5’-TTGTCATACTCCTCTGC 1493-1477 Internal primer: F: K- NP/F1 5’-TGATGCCACATACCAGAG 432-449 R: Bm-NP-756r 5’-TGTTTGGAATTTCCCTTTGAGG 755-734 R: K-NP/R1 5’-CTCTTGGKACCACTCTTG 1125-1108 F: NP2 5’-YAGATATTGGGCTATAAGAAC 1208-1228
a. F: forward primer, R: reverse primer
b. The same primers used for 2838N/00 were used for strain 2838V/00 and 2896/01. c. The same primers used for 2829/00 were used for 2937/01, 3072/03, 3115/03,
3127/03, 3152, and 3153/03. d. K=G/T, Y=C/T
Table 7. Primers used for NA gene cloning and sequencing
Gene Strains Primer Sequence Position
NA 2838N/00 F: Ba-NA-1 Ref. Hoffmann, 2000 -34 - -6
R: Ba-NA-1413R Ref. Hoffmann, 2000 1411-1376
Internal primer: F: Naf 5’-ACGGACCAAGTAATGGACAG 689 - 708 R: Nar 5’-TTCAATACAGCAACAGCCCC 578 - 559 2829/00 F: NC1 5’-ATTGGRTCAATCTGTATGG 28 - 46 R: NC2 5’-CAAYGGTGAATGGCAACT 1360-1343 Internal primer: F: Naf 5’-ACGGACCAAGTAATGGACAG 689 - 708 R: Nar 5’-TTCAATACAGCAACAGCCCC 578 - 559
a. F: forward primer, R: reverse primer
b. The same primers that used for 2838N/00 were used for strain 2838V/00 and 3153/03.
c. The same primers that used for 2829/00 were used for 2896/01, 2937/01, 3072/03, 3115/03, 3127/03, and 3152.
d. Y=C/T, R=A/G
Table 8. Primers used for M gene cloning and sequencing
Gene Strains Primer Sequence Position
M 2838N/00 F: M52F 5’-CTTCTAACCGAGGTCGAAACG 7 - 27
R: Bm-M-1027r Ref. Hoffmann, 2000 1017-982
R: M423R 5’-CCTATGCTGTGAATCAGC 480-463 F: M440F 5’-TGACTATCACCAATCCAC 497-514 2829/00 F: M52F 5’-CTTCTAACCGAGGTCGAAACG 7 - 27 R: M912R 5’-TCCCTCATAGACTCAGGCACTCC 909-887 Internal primer: F: M440F 5’-TGACTATCACCAATCCAC 497-514 R: M423R 5’-CCTATGCTGTGAATCAGC 480-463
a. F: forward primer, R: reverse primer
b. The same primers used for 2838N/00 were used for strain 2838V/00.
c. The same primers used for 2829/00 were used for 2896/01, 2937/01, 3072/03, 3115/03, 3127/03, 3152, and 3153/03.
Table 9. Primers used for NS gene cloning and sequencing
Gene Strains Primer Sequence Position
NS 2829/00 Bm-NS-1 Ref. Hoffmann, 2000 -40 - -12
Bm-NS-890r Ref. Hoffmann, 2000 879 - 845
K-NS/F1 5’-GGGACTGGTTAATGCTCA 299 - 316
a. F: forward primer, R: reverse primer
b. The same primers used for 2829/00 were used for strain 2838N/00, 2838V/00, 2896/01, 2937/01, 3072/03, 3115/03, 3127/03, 3152, and 3153/03.
F iji/15899/83(H 1N 1) P B 2 H K/427/98(H 1N 1) P B 2 H K/470/97(H 1N 1) P B 2 H K/1/ 68(H 3N 2) P B 2 T a iw a n/1/69(H 3N 2) P B 2 T a iw a n/3/71(H 3N 2) P B 2 T a iw a n/2/ 70(H 3N 2 ) P B 2 H K/156/ 97(H 5N 1) P B 2 P h/H K/S H 39/99(H 6N 1) P B 2 Q a /H K/ S F 595/ 00(H 6N 1) P B 2 T e a l/H K/W 312/97(H 6N 1) P B 2 Q a /H K/1721-20/99(H 6N 1) P B 2 P h/ H K/N T 261/ 00(H 6N 1) P B 2 H K/1774/99(H 3N 2) P B 2 2838N /00 P B 2 2838V /00 P B 2 2937/01 P B 2 C k /T a iw a n/7-5/99(H 6N 1) P B 2 G s /G ua ng d o n g /1/96(H 5N 1) P B 2 C k /H K/ 31.4/ 02(H 5N 1) P B 2 C k /G ua ng d o ng /174/04(H 5N 1) P B 2 C k /J ilin/9/04(H 5N 1) P B 2 H K/212/ 03(H 5N 1) P B 2 B ird /T ha ila nd /3.1/04(H 5N 1) P B 2 C k /V ie tna m /C 58/04(H 5N 1) P B 2 V ie tna m /1203/04(H 5N 1) P B 2 V ie tna m /3062/04(H 5N 1) P B 2
Fig. 1. Phylogenetic tree constructed from PB2 gene nucleotide sequences (1–2277). Note Taiwan H6N1 is in different cluster from other viruses including human H3N2, H1N1, and 2004 Asian H5N1.
H K/ 156/ 97(H 5N 1) P B 1 Q a / H K/ 1 721-20/ 9 9(H 6N 1) P B 1 P h/ H K/ N T 261/ 00(H 6N 1) P B 1 Q a / H K/ S F 5 95/ 0 0(H 6 N 1) P B 1 P h / H K/ S H 39 / 99(H 6N 1) P B 1 T e a l/ H K/ W 3 12/ 9 7(H 6 N 1) P B 1 28 38N / 0 0 P B 1 2838 V / 00 P B 1 3 115/ 03 P B 1 C k / T a iw a n/ 7 -5/ 9 9(H 6 N 1) P B 1 3 072/ 03 P B 1 289 6/ 01 P B 1 2937/ 01 P B 1 3 153/ 03 P B 1 312 7/ 03 P B 1 H K/ 1 / 68(H 3N 2 ) P B 1 T a iw a n/ 1/ 69(H 3N 2) P B 1 T a iw a n/ 2/ 70(H 3N 2) P B 1 T a iw a n/ 3/ 71 (H 3N 2) P B 1 H K/ 427/ 98(H 1N 1) P B 1 H K/ 47 0/ 97 (H 1N 1 ) P B 1 F iji/ 15 899/ 83(H 1N 1) P B 1 H K/ 1 774/ 99(H 3N 2) P B 1 G s / G u a n g d o ng / 1/ 96 P B 1 C k / J ilin/ 9 / 04(H 5N 1) P B 1 C k / H K/ 3 1.4/ 02 (H 5N 1) P B 1 C k / G ua n g d o n g / 1 74/ 0 4(H 5 N 1) P B 1 H K/ 212/ 03(H 5 N 1) P B 1 V ie na m / 1203 / 04(H 5N 1) P B 1 C k / V ie tn a m / C 58/ 04(H 5 N 1) P B 1 B ird / T ha ila nd / 3.1 / 04(H 5N 1) P B 1 V ie tna m / 30 62/ 0 4(H 5 N 1) P B 1
Fig. 2. Phylogenetic tree constructed from PB1 gene nucleotide sequences (1–2271). Except Fiji/15899/83(H1N1) (1-2089), Ck/HK/31.4/02(H5N1) (1-2163),
HK/1774/99(H3N2) (1-2268), Bird/Thailand/3.1/2004(H5N1) 1-2269), and Vietnam/3062/04(H5N1) (1-2269).
Note Taiwan H6N1 is in different cluster from other viruses including human H3N2, H1N1, and 2004 Asian H5N1.
2 838N / 00 P A 2838 V / 00 P A C k / T a iw a n / 7-5/ 99(H 6 N 1) P A 2937 /01 P A C k /G ua ng d o ng / 174/0 4(H 5N 1) P A F iji/ 1589 9/83(H 1N 1) P A H K/ 4 2 7 / 9 8 (H 1N 1) P A H K/ 4 7 0 / 9 7 (H 1N 1) P A H K/ 1/ 68(H 3 N 2) P A T a iw a n/ 1/ 69(H 3 N 2) P A T a iw a n / 3 / 7 1 (H 3N 2 ) P A T a iw a n/ 2/ 70 (H 3N 2) P A H K/ 1774/ 99(H 3 N 2) P A H K/156/ 97(H 5N 1) P A P h / H K/ N T 2 6 1 / 0 0 (H 6N 1) P A T e a l/ H K/ W 3 1 2 / 9 7 (H 6N 1) P A Q a / H K/ S F 5 9 5 / 0 0(H 6 N 1 ) P A P h/ H K/ S H 39 / 99(H 6N 1) P A Q a / H K/ 1 721-2 0/99(H 6N 1) P A G s / G u a n g d o n g / 1 / 9 6 (H 5N 1) P A C k / J ilin / 9 / 0 4 (H 5N 1 ) P A C k / H K/ 3 1.4/ 0 2 (H 5N 1) P A H K/212/ 03(H 5N 1) P A C k / V ie tn a m / C 5 8 / 0 4 (H 5 N 1 ) P A V ie tna m / 1203/ 04(H 5N 1) P A B ird / T h a ila nd / 3.1/04(H 5N 1) P A V ie tna m / 3062/ 04(H 5N 1) P A
Fig. 3. Phylogenetic tree constructed from PA gene nucleotide sequences (1-2148) except Bird/Thailand/3.1/2004 (H5N1) (10-2148), HK/212/03 (H5N1) (13-2148), Vietnam/3062/04 (H5N1) (16-2148).
2829/00 H A 2896/01 H A 2937/01 H A 3072/03 H A 3127/03 H A 2838N /00 H A 2838V /00 H A 3115/03 H A 3153/03 H A C k /T a iw a n/7-5/99(H 6N 1) H A Ck /T a iwa n/na 3/98(H 6N 1) H A P h/H K/N T 261/00(H 6N 1) H A Q a /H K/S F 595/00(H 6N 1) H A P h/H K/S H 39/99(H 6N 1) H A Q a /H K/1721-20/99(H 6N 1) H A T e a l/H K/W 312/97(H 6N 1) H A D k /H a ina n/6/04(H 6N 2) H A D k /H K/202/77(H 6N 1) H A
P inta il/A lb e rta /179/93(H 6N 1) H A D k /Ko re a /S 17/03(H 6N 1) H A D k /H K/3461/99(H 6N 1) H A
Fig. 4. Phylogenetic tree constructed from HA gene (H6) nucleotide sequences (1-1698) except Ck/Taiwan/7-5/99 (H6N1) (1-1110), Ck/Taiwan/na3/98 (H6N1) (1-1110)
V ie tn a m / 3 0 62 / 0 4(H 5N 1 ) N P B ird / T h a ila n d / 3 .1 / 0 4 (H 5 N 1 ) N P V ie tn a m / 1 20 3 / 04 (H 5 N 1) N P C k / V ie tna m / C 5 8/ 0 4(H 5N 1 ) N P H K/ 2 12 / 03 (H 5N 1 ) N P C k / G u a n g d o n g / 17 4 / 0 4 (H 5 N 1 ) N P C k / J ilin / 9 / 04 (H 5N 1 ) N P H K/ 1 56 / 97 (H 5N 1 ) N P C h uk k a / H K/ N T 2 6 1/ 0 0(H 6N 1 ) N P P h / H K/ S H 3 9 / 9 9 (H 6 N 1 ) N P Q a / H K/ S F 5 9 5 / 0 0 (H 6 N 1 ) N P T e a l/ H K/ W 3 1 2 / 9 7 (H 6 N 1 ) N P Q a / / H K/ 1 7 21 -20 / 99 (H 6 N 1) N P F iji/ 15 8 99 / 83 (H 1N 1 ) N P H K/ 4 27 / 98 (H 1N 1 ) N P H K/ 4 70 / 97 (H 1 N 1 ) N P H K/ 1 / 68 (H 3 N 2 ) N P T a iw a n / 1 / 69 (H 3 N 2 ) N P T a iw a n / 3 / 71 (H 3 N 2 ) N P T a iw a n/ 2 / 7 0 (H 3 N 2) N P H K/ 1 7 7 4/ 9 9(H 3N 2 ) N P C k / H K/ 31 .4 / 0 2 (H 5 N 1) N P G s / G u a n g d o ng / 1/ 9 6(H 5N 1 ) N P 2 93 7 / 01 N P 2 8 9 6/ 0 1 N P 3 1 5 3/ 0 3 N P 31 2 7 / 0 3 N P 3 0 7 2/ 0 3 N P 2 82 9 / 00 N P C k / T a iw a n/ 7 -5 / 99 (H 6 N 1 ) N P 28 3 8N / 0 0 N P 3 11 5 / 0 3 N P 28 3 8V / 0 0 N P
Fig. 5. Phylogenetic tree constructed from NP gene nucleotide sequences (7-1493) except Vietnam/3062/2004 (H5N1) (7-1479), Bird/Thailand/3.1/2004 (H5N1) (7-1484)
Note Taiwan H6N1 is in different cluster from other viruses including human H3N2, H1N1, and 2004 Asian H5N1.
D k /H K/3461/99(H 6N 1) N A F iji/15899/83(H 1N 1) N A P h/H K/N T 261/00(H 6N 1) N A Q a /H K/S F 595/00(H 6N 1) N A P h/H K/S H 39/99(H 6N 1) N A T e a l/H K/W 312/97(H 6N 1) N A Q a /H K/1721-20/99(H 6N 1) N A H K/156/97(H 5N 1) N A G s /G ua ng d o ng /1/96(H 5N 1) N A B ird /T ha ila nd /3.1/04(H 5N 1) N A Ck /V ie tna m /C58/04(H 5N 1) N A V ie tna m /1203/04(H 5N 1) N A V ie tna m /3062/04(H 5N 1) N A Ck /G ua ng d o ng /174/04(H 5N 1) N A C k /J ilin/ 9/04(H 5N 1) N A H K/212/03(H 5N 1) N A 3153/03 N A 3072/03 N A 3127/03 N A 2829/00 N A 2896/01 N A 2838N /00 N A 2838V /00 N A 3115/03 N A Ck /T a iwa n/7-5/99(H 6N 1) N A 2937/01 N A
Fig. 6. Phylogenetic tree constructed from NA gene (N1) nucleotide sequences (28-1333).
2829/00 M Ck /T a iwa n/7-5/99(H 6N 1) M 2838N /00 M 2838V /00 M 3115/03 M 2896/01 M 2937/01 M 3072/03 M 3127/03 M 3153/03 M B ird /T ha ila nd /3.1/04(H 5N 1) M C k /V ie tna m /C58/04(H 5N 1) M V ie tna m /1203/04(H 5N 1) M V ie tna m /3062/04(H 5N 1) M H K/212/03(H 5N 1) M Ck /G ua ng d o ng /174/04(H 5N 1) M C k /H K/31.4/02(H 5N 1) M G s /G ua ng d o ng /1/96(H 5N 1) M C k /J ilin/9/04(H 5N 1) M H K/156/97(H 5N 1) M Q a /H K/1721-20/99(H 6N 1) M P h/H K/N T 261/00(H 6N 1) M Q a /H K/S F 595/00(H 6N 1) M T e a l/H K/W 312/97(H 6N 1) M P h/H K/S H 39/99(H 6N 1) M F iji/15899/83(H 1N 1) M H K/427/98(H 1N 1) M H K/470/97(H 1N 1) M H K/1/68(H 3N 2) M T a iw a n/1/69(H 3N 2) M T a iwa n/2/70(H 3N 2) M T a iwa n/3/71(H 3N 2) M H K/1774/99(H 3N 2) M
Fig. 7. Phylogenetic tree constructed from M gene nucleotide sequences (7-909). Note Taiwan H6N1 is in different cluster from other viruses including human H3N2, H1N1, and 2004 Asian H5N1.
H K/ 47 0/ 97(H 1N 1) N S H K/ 42 7/ 98(H 1N 1) N S F iji/ 1 589 9/ 83(H 1N 1) N S T a iw a n/ 1/ 6 9(H 3N 2 ) N S H K/ 1/ 6 8(H 3N 2) N S T a iw a n/ 2/ 7 0(H 3N 2 ) N S T a iw a n/ 3/ 7 1(H 3N 2 ) N S G S / G u a n g d o ng / 1 / 96 (H 5 N 1) N S H K/ 1 774 / 9 9(H 3N 2 ) N S H K/ 15 6/ 9 7(H 5N 1) N S Q a / H K/ 1 721 -2 0/ 9 9(H 6N 1 ) N S P h / H K/ N T 261 / 0 0(H 6N 1 ) N S Q a / H K/ S F 5 95 / 00 (H 6 N 1) N S P h/ H K/ S H 3 9/ 99(H 6N 1) N S T e a l/ H K/ W 312 / 9 7(H 6N 1 ) N S D k / H K/ 3 461 / 99 (H 6N 1 ) N S D k / Ko re a / S 17 / 03 (H 6 N 1 ) N S B ird / T h a ila nd / 3 .1/ 0 4(H 5N 1) N S V ie tna m / 30 62/ 04(H 5 N 1) N S V ie tna m / 120 3/ 0 4(H 5N 1) N S C k / V ie tna m / C 58/ 04 (H 5 N 1) N S H K/ 2 12/ 03(H 5N 1) N S C k / G u a n g d o ng / 17 4/ 04(H 5N 1) N S C k / J ilin/ 9 / 04 (H 5 N 1 ) N S C k / H K/ 3 1.4/ 02(H 5N 1) N S 28 29 / 00 N S C k / T a iw a n / 7-5/ 9 9(H 6N 1) N S 2 89 6/ 0 1 N S 293 7/ 01 N S 3 15 3/ 0 3 N S 28 38 N / 0 0 N S 2 83 8V / 0 0 N S 31 15 / 03 N S 30 72 / 03 N S 3 127 / 03 N S
Fig. 8. Phylogenetic tree constructed from NS gene nucleotide sequences (1-829 to 864)