中文摘要
第一部份 功能性 T 細胞對於對抗感染是非常重要的。文獻報告指出,利用其產生細胞激 素的能力所定義之功能性 T 細胞,可代表在疾病感染模式中具抗原特異性的 T 細胞。在本實驗中,我們計算在脾臟中產生干擾素γ 之組織胞漿菌特異性細胞數 目,並發現一種巨噬細胞胞內病源菌-組織胞漿菌之感染可以活化 CD4 及 CD8 T 細胞。CD8 T 細胞反應強度雖較 CD4 T 細胞的反應強度弱,二種細胞反應之擴 張與收縮的動力學則是相同的模式。超過 90% 的產生干擾素 γ 之 CD4 T 細胞 及超過 85% 的產生干擾素 γ 之 CD8 T 細胞均表現高量之 CD44。產生干擾素 γ 與高量 CD44 表現之間的高度相關性不只在反應高峰期可觀察到,在整個感染 過程均是如此。此外,廣大系列 Vβ 族群均會對全身性以及肺部感染產生反應, 代表在抵抗組織胞漿菌之初次免疫反應中並無使用明顯 T 細胞受體傾向。幾乎 每一個Vβ 族群都對產生干擾素 γ 有貢獻,數個特定 Vβ 族群組成較大比例之產 生干擾素γ 細胞。我們的研究對於後續對組織胞漿菌感染所引發之免疫反應的探 討提供基礎之了解。此部分已被接受發表於 International Immunology, November 2004
第二部份
CD8 T 細胞對於宿主抵抗組織胞漿菌感染的貢獻,在 CD4 T 細胞完整之小鼠是
較其次的,因若去除CD8 T 細胞只會延遲但並不影響病菌的清除。然而,CD8 T
討。本實驗室研究中發現,MHC class II 基因缺陷小鼠在感染組織胞漿菌後能 將病菌數量維持如同第一週之數目長達十六週,顯示CD8 T 細胞能夠限制病菌 之複製。研究結果顯示,由組織胞漿菌感染小鼠體內取出之CD8 T 細胞胞內會 表現干擾素γ 及 granzyme B。此外,CD8 T 細胞也具有細胞毒殺的功能,能夠 毒殺含組織胞漿菌的IC-21 巨噬細胞。因此,CD8 T 細胞對於宿主抵抗胞內非病 毒之病源菌感染是相當重要的,尤其當在宿主缺乏功能性 CD4 T 細胞的情況 下。 此部分已寫成論文投稿至Journal of Immunology 目前在修改中 關鍵字: CD4 T 細胞、CD8 T 細胞、組織胞漿菌、功能性 T 細胞
Abstract
Part I.
Functional T cells are critical to host defense against infection. It has been reported that functional T cells as determined by their cytokine production represent antigen-specific T cells in infectious disease models. In this study, we enumerated
Histoplasma-specific interferon γ-producing cells in bulk splenocyte culture and
showed that infection with Histoplasma capsulatum, an intracellular pathogen of the macrophage, activated both CD4 and CD8 T cells. The magnitude of CD8 T cell response was lower than CD4 T cell, but the expansion and contraction of both cell types followed the same kinetics. Over 90% of interferon γ-producing CD4 T cells and >85% of CD8 T cells expressed CD44hi phenotype. The strong correlation between interferon γ production and CD44hi expression was observed not only at the peak of response but also throughout the course of infection. Moreover, a broad spectrum of Vβ populations responded to systemic as well as pulmonary infections, suggesting no obvious T cell receptor bias in primary immune response to histoplasmosis. While each Vβ population contributed to interferon γ production, several specific Vβ populations made up higher percentages of interferon γ-producing cells. Our study laid the groundwork for further investigations in immune response to histoplasmosis.
Part II.
The contribution of CD8 T cells in host defense against histoplasmosis is minor in the CD4-T-cell-intact mouse, as it has been shown that depleting CD8 T cells delays but does not affect fungal clearance. However, it remains to be determined whether the CD8 T cells are protective in a host lacking functional CD4 T cells. In this study, MHC class II-deficient mice infected with Histoplasma kept the fungus in check for up to 16 weeks, indicating CD8 T cells are able to limit fungal replication. Ex vivo studies showed that CD8 T cells from Histoplasma-infected mice expressed both intracytoplasmic interferon-gamma and granzyme B. Furthermore, CD8 T cells exhibited cytotoxic activity against IC-21 macrophage targets containing Histoplasma. Therefore, CD8 T cells are critical for host defense against the intracellular nonviral pathogen, especially in a host lacking functional CD4 T cells. (In revision)
Part I.
The paper has been published on International Immunology, 16: 1663-1673, 2004, the PDF file of the publication is attached.
Part II.
Background and Aims
Infection with HIV generates an immune response that usually contains but does not eliminate the virus. Both HIV-specific CD4 Th cells and CD8 CTL cells are activated. However, with progressive HIV infection, the CD4 T-cell counts gradually decline and are finally lost, while total CD8 T-cell counts increase and remain constant for a prolonged period of time (1-3). Therefore, there is much to be learned about the function of CD8 T cells in the absence of CD4 T cells, especially in defense against opportunistic infections.
Histoplasma capsulatum is an opportunistic fungal pathogen that kills AIDS
patients (4). Depletion (5) and adoptive transfer (6) studies have established the vital role of CD4 T cells in clearing Histoplasma in the mouse model. The functional role of CD4 T cells is to produce IFNγ (7-9). Depleting CD4 T cells dramatically decreases IFNγ production and accelerates mortality in mice (8, 9). On the contrary, CD8 T cell depletion or β2m deficiency only marginally affects fungal clearance (9,
10). Since the role of CD8 T cells in a CD4-T-cell-intact host against histoplasmosis is comparatively minor, not much is known about CD8 T cell function or how they are activated in Histoplasma infection. Considering the importance of CD8 T cells in defense against infections in patients with low CD4 T cell counts, a thorough study of CD8 T cells against this fungal pathogen is warranted.
In this study, we first established the importance of CD8 T cells in protecting mice lacking functional CD4 T cells against histoplasmosis. We also showed CD8 T cells function by both perforin-independent as well as –dependent mechanisms, because CD8 T cells isolated from Histoplasma-infected mice produce IFNγ as well as kill Histoplasma-containing IC-21 macrophage targets in vitro through
perforin-mediated mechanism. Therefore, CD8 T cells are functional and critical in host defense against histoplasmosis in the absence of CD4 T cells.
Materials and Methods
Mice
Wild type (WT), MHC class II-deficient (IIKO) (11), C57BL/6-Prf1tm1Sdz
perforin-deficient mice (PKO, stock number 002407, (12)) and B6.129P2-B2mtm1Unc
β2m-knockout (β2m-/-, stock number 002087, (13)) mouse breeders were obtained
from The Jackson Laboratory (Bar Harbor, ME), and bred at the Laboratory Animal Center, National Taiwan University College of Medicine. Mice were housed in sterilized cages with sterilized beddings and filter cage tops and fed with sterilized food and water. Mice at 8 to 10 wk of age were used in all the experiments. All mice with different genetic deficiencies are on the C57BL/6 background.
Fungus and Infection
Histoplasma capsulatum strain 505 yeast cells were cultured at 37℃ on brain-heart
infusion (BHI) agar supplemented with cysteine (1mg/ml) and glucose (20mg/ml). Fresh yeast cell suspensions were prepared in RPMI 1640 medium (GIBCO-BRL, Grand Island, NY) for injection. Mice were injected intravenously with 2.5×104 yeast cells.
Reagents
RPMI 1640 medium (GIBCO-BRL) was supplemented with 10% heat-inactivated FCS (Biological Industries, Israel), 1mM sodium pyruvate, 2mM L-glutamine, 0.1mM nonessential amino acid, 100U/ml penicillin, 100μg/ml streptomycin, 5×10-5
M 2-ME, and 25mM HEPES buffer. All supplements were obtained from GIBCO-BRL. Dulbecco’s phosphate buffered saline (dPBS, Biological Industries) supplemented with 1% FCS and 0.1% NaN3 (Sigma-Aldrich, St. Louis, MO) was
used as staining buffer. Dulbecco’s PBS supplemented with 1% FCS, 0.1% NaN3 and
0.1% saponin (Sigma-Aldrich) was used as Perm/Wash buffer.
Quantitation of fungal load in the spleen
Spleens were harvested from mice at different time points after infection. The spleen was homogenized in a tissue grinder with 1ml of RPMI 1640 medium. One to 10 serial dilutions were made and 0.1ml was plated onto glucose-peptone agar.
Histoplasma mycelial colonies were enumerated after incubation at 30℃ for 10 to 14
days.
Splenic T cell enrichment by nylon-wool passage and CD4 and CD8 T cell purification by MACS beads
Spleen cells were harvested from mice at day14 of infection. Single-cell suspensions were applied to pre-rinsed nylon-wool columns. After incubation in an upright position at 37oC incubator for 90 min the nonadherent cells were eluted. Nearly 80% of the cells collected were CD3+ T cells.
To purify CD4 and CD8 T cells, nylon-wool-passaged cells were washed and resuspended in MACS buffer (PBS with 0.5% FCS) containing anti-CD4 or anti-CD8 magnetic microbeads (Miltenyi Biotec, Germany). The suspension was mixed and incubated at 4oC for 30 min. The positive selection column (LS+/VS+ column for up to 108 total cells) was prepared by washing with buffer. The cell suspension was
loaded onto the column and the negative cells were allowed to pass through the column in a magnetic field. To elute positive cells, the column was removed from the separator. A plunger which was supplied by the manufacturer was used to flush out the positive fraction. To release beads from positively selected cells, the cells were further incubated at 37℃ for 90 min followed by centrifugation. In the resuspended cell pellet, about 95% of cells was either CD4+ or CD8+ T cells after MACS purification.
In vivo cell depletion
To deplete CD4+ or CD8+ T cells, mice were treated with concentrated anti-CD4 (clone GK1.5) or anti-CD8 (clone 2.43) hybridoma supernatant at the time of infection and twice weekly until concluding the experiment. The in vivo depletion efficiency was confirmed by staining splenic and lymph node cells with anti-CD4 (clone RM4-4) or anti-CD8 (clone 53-6.7) mAb. The results showed that GK1.5 treatment depleted >95% CD4 T cells and 2.43 treatment depleted >99% CD8 T cells.
Intracytoplasmic cytokine, granzyme B staining and flow cytometry
One million nylon-wool-enriched T cells, or MACS-purified CD8+ T cells were cultured in flat-bottom 96-well plates in medium containing 2.5×104 live Histoplasma yeast cells for 24 h. At 6 h before harvest, monensin (2μM, Sigma-Aldrich) was added. Cells were harvested and stained with allophycocyanin-conjugated anti-CD4 (clone GK1.5) or anti-CD8 (clone 53-6.7) at 4℃ in the dark for 30 min. Cells were washed and then fixed in Cytofix (4% paraformaldehyde in dPBS) for 20 min on ice and permeabilized by two washes with Perm/Wash solution. To stain intracellular
granzyme B, the cells were treated with purifed 2.4G2 antibody directed against mouse FcγII/III receptors (Fc Block) and stained with PE-labeled mouse anti-human granzyme B mAb (Caltag, Burlingame, CA) of which the specificity has been confirmed (14). PE-labeled mouse IgG1 was used as isotype control. To stain for intracellular cytokine, FITC- or PE-conjugated anti-IFNγ mAb (clone XMG1.2) in the Perm/Wash buffer was added and cells were left on ice in the dark for 30 min. Cells were washed with Perm/Wash buffer and fixed in staining buffer/2% paraformaldehyde. FACSCalibur flow cytometer (BD Biosciences, Mountain View, CA) was used for cell acquisition. The data were analyzed by CellQuest (BD Biosciences). All antibodies except granzyme B were purchased from eBioscience (San Diego, CA). Dead cells were excluded from analysis by both forward- and side- scattering.
Cytotoxicity assay
Mouse macrophage cell line IC-21 (ATCC TIB-186, C57BL/6 origin, SV40 transformed) was used as target cells. IC-21 cells at 1×104 per well were cultured in triplicate or quadruplicate in 96-well round bottom plate. After 2 h adhesion, heat-killed (1×105) Histoplasma yeasts were added to the wells for another 2 h. Free extracellular yeasts were removed by several washes with warm HBSS. The cultures were incubated for 16-18 h. Splenic T cells were obtained from normal or infected mice pre-treated with anti-CD4 mAb (clone GK1.5) two days prior to experiment. After nylon-wool enrichment, CD8 T cells were purified by MACS as described above. Different numbers of CD8 T cells were added to the wells containing antigen-pulsed IC-21 cells to obtain different E/T ratios. After 6 h of incubation, 50μl of culture supernatant was removed from the wells and transferred to an ELISA plate.
An equal volume of substrate provided in CytoTox96® Non-Radioactive Cytotoxicity Assay kit (Promega, Madison, WI) was added. The coupled-enzymatic assay detects lactate dehydrogenase (LDH) in the culture supernatnts. The plates were left at room temperature in the dark for 30 min to allow conversion of a tetrazolium salt (INT) into a red formazan product. The reaction was stopped by Stop Solution and color absorbance was read at 492 nm. The amount of color formed is proportional to the number of lysed cells. Thus, % cytotoxicity was calculated from the absorbance values as follows:
Experimental – Effector Spontaneous – Target Spontaneous % Cytotoxicity =
Target Maximum – Target Spontaneous
× 100%
Statistics
Results
CD8 T cells are critical in host defense against histoplasmosis in the absence of CD4 T cells
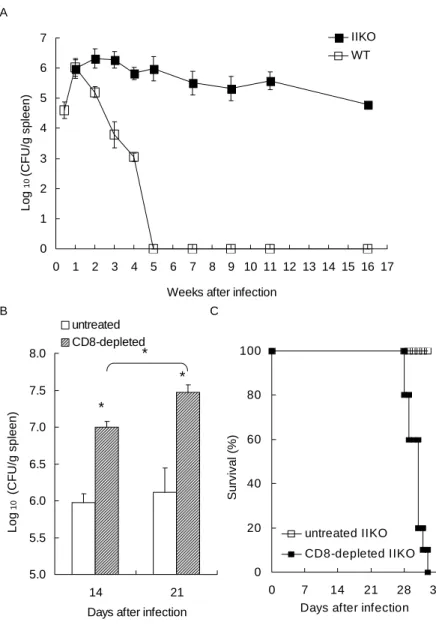
To address whether CD8 T cells are protective against histoplasmosis, MHC class II-deficient mouse (IIKO) were infected with a sublethal dose of Histoplasma yeast cells. The IIKO mice lacking mature CD4 T cells were able to contain the fungal burden at the same level as that at wk 1 through wk 16 (Fig. 1A). To further confirm that CD8 T cells and not other cells in the IIKO mice were responsible for maintaining the chronic state, IIKO mice were depleted of CD8 T cells and infected with the same dose of Histoplasma yeast cells as was given to IIKO mice that were not depleted of CD8 T cells. Depleting CD8 T cells not only significantly increased the fungal burden (p < 0.05, Fig. 1B) but also resulted in 100% mortality (Fig. 1C). These results clearly show that CD8 T cells alone, although not able to clear the infection, are functional in limiting fungal replication and are protective in the absence of CD4 T cells.
CD8 T cell-mediated protection is perforin-dependent as well as perforin–independent
We reasoned that CD8 T cells could be contributing to holding Histoplasma in check by (i) production of IFN γ which would activate macrophages to limit its intracellular growth and/or (ii) via a perforin/granzyme B-mediated cytotoxic effect on the infected macrophages. To determine the mechanism(s) whereby CD8 T cells defend the host against histoplasmosis, we compared perforin-deficient (PKO) mice
to those with MHC class I deficiency (β2m-/-) in clearing the fungus. The results
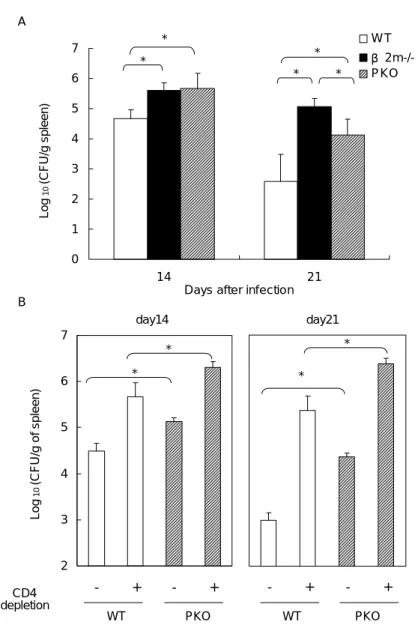
show that both β2m-/- and PKO mice had greater fungal burden than WT controls (p
< 0.05) (Fig. 2A), confirming that CD8 T cells are involved in limiting fungal replication. These results also showed that perforin was important in controlling the fungus. However, fungal counts in β2m-/- mice were higher than in PKO mice at
day21 (p < 0.05) of infection, suggesting a CD8 T cell-mediated perforin-independent mechanism was also operative in controlling the fungus. Together these results strongly demonstrate that CD8 T cell-mediated perforin-dependent as well as -independent mechanisms are operative in controlling the fungus.
To better define the protective role of perforin in the absence of CD4 T cells, the PKO mice were treated with anti-CD4 Ab and their ability to clear the fungus was evaluated. The results show that CD4 T cell depletion increased fungal burden in both PKO mice and WT mice as it has been reported (p < 0.05, Fig. 2B) (5, 9). More importantly, PKO mice depleted of CD4 T cells had greater fungal counts than WT mice with the same treatment, indicating perforin-dependent cytotoxic activity was critical in restricting the growth of Histoplasma in mice without CD4 T cells (p < 0.05, Fig. 2B).
CD8 T cells from Histoplasma-infected mice have cytotoxic function
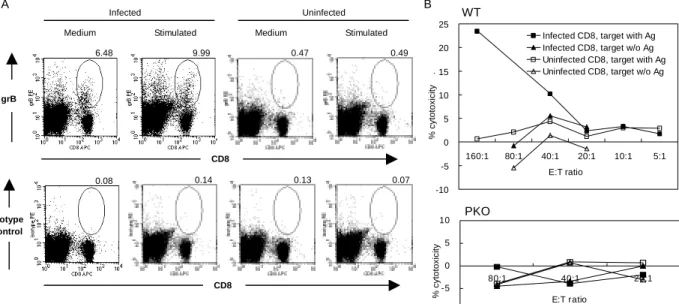
The cytotoxic granules of CD8 T cells contain both pore-forming perforin and granzymes, which belong to the family of serine proteases (15, 16). Upon delivery to target cells, granzyme B, the most abundant granzyme in the granules, induces caspase-dependent as well as -independent (15, 16) cytoxicity. The appearance of intracytoplasmic granzyme B in Ag-specific CD8 T cells correlates well with their cytotoxic function (14). Since the specificity of the anti-granzyme B mAb has been
confirmed (14) and the isotype control was negative (Fig. 3A), we used granzyme B expression as a functional marker for CD8 T cell cytotoxicity. The results show that CD8 T cells from Histoplasma-infected mice expressed granzyme B with or without further antigenic stimulation in vitro whereas CD8 T cells from normal uninfected mice did not. It is noticeable that a non-CD8 T population also expressed granzyme B. These cells were identified to be NK1.1-, B220-, CD11c-, and CD4- (data not shown). Importantly, CD8 T cells from infected WT mice at an effector:target cell ratio of 160:1, killed 25% of macrophage IC-21 targets containing Histoplasma yeasts, while CD8 T cells from infected PKO mice did not exhibit cytotoxity (Fig. 3B). Taken together, these data demonstrated the role of cytotoxicity by CD8 T cells as a protective mechanism in Histoplasma-infected mice.
CD8 T cells from Histoplasma-infected mice produce IFNγ
Since IFNγ is critically important in host defense against histoplasmosis (7-9, 17, 18), we examined whether CD8 T cells in Histoplasma-infected mice were activated to produce IFNγ. Intracytoplasmic cytokine and surface staining revealed that CD8 T cells, similar to CD4 T cells, produced IFNγ (Fig. 4), although the CD8 T cell response was less than that of CD4 T cell. These data together with those in Fig. 2A demonstrate that activated CD8 T cells produce IFNγ which could account, at least in part, for the perforin-independent anti-Histoplasma activity.
Discussion
In CD4-T-cell-intact mice, the contribution of CD8 T cells to defense against histoplasmosis is minor (9, 10). Thus, the functional role of CD8 T cells in
Histoplasma infections has not been thoroughly studied. In the present study, we
demonstrate that CD8 T cells alone in the absence of functional CD4 T cells, as in the situation of opportunistic infection in patients with low CD4 T cell counts, provide protection against Histoplasma infection. We further showed the functions of CD8 T cells in normal mice include IFN γ production and killing antigen-containing macrophage targets.
Perforin-mediated host defense against histoplasmosis has been reported (9). Perforin-/- mice exhibit increased fungal burden and accelerated mortality after infection (9). However, in vitro CD8 T cell cytotoxicity activity has never been demonstrated. Nor has it been clarified whether perforin-dependent anti-Histoplasma activity was attributed to CD8 T cells or NK cells. In this study, we showed specifically that CD8 T cells from Histoplasma-infected mice express granzyme B and exhibit cytotoxic activity against Histoplasma-containing IC-21 macrophage targets. These results provide strong evidence that CD8 T cells in mice infected with
Histoplasma have cytotoxic function.
Histoplasma yeast cells are facultative intracellular organisms that reside within
the macrophages (8, 17, 19). It has been reported that macrophages activated by IFN γ are armed to inhibit the intracellular growth of the fungus (20-22). CD8 T cell killing target cells by cytotoxic activity through granule exocytosis kills the intracellular pathogen (16, 23). Thus, by producing IFNγ and exhibiting cytotoxic activity against macrophage targets, the CD8 T cells in Histoplasma-infected mice contribute to restricting intracellular growth and killing of the fungus within the
Reference
1. Altfeld, M., and E. S. Rosenberg. 2000. The role of CD4(+) T helper cells in the cytotoxic T lymphocyte response to HIV-1. Curr Opin Immunol 12:375. 2. Grossman, Z., R. B. Herberman, N. Vatnik, and N. Intrator. 1998.
Conservation of total T-cell counts during HIV infection: alternative
hypotheses and implications. J Acquir Immune Defic Syndr Hum Retrovirol
17:450.
3. Rabson, A. 1995. Enumeration of T-cells subsets in patients with HIV infection. AIDS Clin Care 7:1.
4. Wheat, L. J., P. A. Connolly-Stringfield, R. L. Baker, M. F. Curfman, M. E. Eads, K. S. Israel, S. A. Norris, D. H. Webb, and M. L. Zeckel. 1990. Disseminated histoplasmosis in the acquired immune deficiency syndrome: clinical findings, diagnosis and treatment, and review of the literature.
Medicine (Baltimore) 69:361.
5. Gomez, A. M., W. E. Bullock, C. L. Taylor, and G. S. Deepe, Jr. 1988. Role of L3T4+ T cells in host defense against Histoplasma capsulatum. Infect Immun
56:1685.
6. Allendoerfer, R., D. M. Magee, G. S. Deepe, Jr., and J. R. Graybill. 1993. Transfer of protective immunity in murine histoplasmosis by a CD4+ T-cell clone. Infect Immun 61:714.
7. Wu-Hsieh, B. 1989. Relative susceptibilities of inbred mouse strains C57BL/6 and A/J to infection with Histoplasma capsulatum. Infect Immun 57:3788. 8. Wu-Hsieh, B. A., and D. H. Howard. 1993. Histoplasmosis. In Fungal
infections and immune responses. J.Murphy, H. Friedman, and M. Bendinelli,
eds. Plenum Press, New York, p. 213.
requirements for nascent and memory immunity in pulmonary histoplasmosis.
J Immunol 162:7389.
10. Deepe, G. S., Jr. 1994. Role of CD8+ T cells in host resistance to systemic infection with Histoplasma capsulatum in mice. J Immunol 152:3491. 11. Grusby, M. J., R. S. Johnson, V. E. Papaioannou, and L. H. Glimcher. 1991.
Depletion of CD4+ T cells in major histocompatibility complex class II-deficient mice. Science 253:1417.
12. Kagi, D., B. Ledermann, K. Burki, P. Seiler, B. Odermatt, K. J. Olsen, E. R. Podack, R. M. Zinkernagel, and H. Hengartner. 1994. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice.
Nature 369:31.
13. Koller, B. H., P. Marrack, J. W. Kappler, and O. Smithies. 1990. Normal development of mice deficient in beta2M, MHC class I proteins, and CD8+ T cells. Science 248:1227.
14. Wherry, E. J., V. Teichgraber, T. C. Becker, D. Masopust, S. M. Kaech, R. Antia, U. H. von Andrian, and R. Ahmed. 2003. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat Immunol 4:225. 15. Lieberman, J. 2003. The ABCs of granule-mediated cytotoxicity: new
weapons in the arsenal. Nat Rev Immunol 3:361.
16. Barry, M., and R. C. Bleackley. 2002. Cytotoxic T lymphocytes: all roads lead to death. Nat Rev Immunol 2:401.
17. Wu-Hsieh, B. A., and D. H. Howard. 1987. Inhibition of the intracellular growth of Histoplasma capsulatum by recombinant murine gamma interferon.
Infect Immun 55:1014.
18. Allendoerfer, R., and G. S. Deepe, Jr. 1997. Intrapulmonary response to Histoplasma capsulatum in gamma interferon knockout mice. Infect Immun
65:2564.
19. Newman, S. L. 1999. Macrophages in host defense against Histoplasma capsulatum. Trends Microbiol 7:67.
20. Wu-Hsieh, B. A., W. Chen, and H. J. Lee. 1998. Nitric oxide synthase expression in macrophages of Histoplasma capsulatum-infected mice is associated with splenocyte apoptosis and unresponsiveness. Infect Immun
66:5520.
21. Lane, T. E., B. A. Wu-Hsieh, and D. H. Howard. 1993. Gamma interferon cooperates with lipopolysaccharide to activate mouse splenic macrophages to an antihistoplasma state. Infect Immun 61:1468.
22. Nakamura, L. T., B. A. Wu-Hsieh, and D. H. Howard. 1994. Recombinant murine gamma interferon stimulates macrophages of the RAW cell line to inhibit intracellular growth of Histoplasma capsulatum. Infect Immun 62:680. 23. Russell, J. H., and T. J. Ley. 2002. Lymphocyte-mediated cytotoxicity. Annu
FIGURE 1. CD8 T cells are critical in controlling Histoplasma infection in the
absence of CD4 T cells. (A) Fungal counts in the spleen. (B) Fungal counts in IIKO mice depleted of CD8 T cells. IIKO mice were treated with anti-CD8 mAb twice weekly from day0 (* p < 0.05, n = 3 in each group). (C) Survival of IIKO mice with or without CD8 T cell depletion. IIKO mice were giving the same dose of
Histoplasma yeast cells as in (A). n = 10 in each group.
5.0 5.5 6.0 6.5 7.0 7.5 8.0 14 21
Days after infection
Log 10 (CF U /g spl een) untreated CD8-depleted * * * B 0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Weeks after infection
Log 10 (C FU /g s p leen) IIKO WT A 0 20 40 60 80 100 0 7 14 21 28 35 Days after infection
S urv iv al (% ) untreated IIKO CD8-depleted IIKO C
FIGURE 2. CD8 T cells limit Histoplasma replication by perforin-dependent as
well as –independent mechanisms. (A) Perforin or β2m deficiency impairs fungal
clearance. (* p < 0.05, n = 3-5). (B) Perforin-dependent mechanism contributes to fungal clearance in the absence of CD4 T cells. WT and PKO mice (3/group) were treated with anti-CD4 mAb twice weekly from day0 (* p < 0.05). Experiment was repeated two times.
2 3 4 5 6 7 - + - + Log 10 ( C F U /g of spl een) - + - + B day14 day21 * * * * 0 1 2 3 4 5 6 7 14 21
Days after infection
Log 10 (CF U /g spleen) W T β 2m-/-PKO A * * * * * WT PKO WT PKO CD4 depletion
FIGURE 3. CD8 T cells in Histoplasma-infected mice are cytotoxic. (A) Granzyme
B expression. Spleen cells from Histoplasma-infected WT mice were stained with mouse anti-human granzyme B (grB) mAb. PE-conjugated mouse IgG1 was used as the isotype control. The numbers indicate the percentage of CD8 T cells expressing granzyme B (CD8+grB+/CD8+). The data shown are from one representative mouse. Three mice were used in each experiment and the experiment was repeated three times. (B) CD8 T cells are cytotoxic against Histoplasma-containing IC-21 macrophages. CD8+ T cells were isolated from uninfected or infected WT (upper panel) or PKO (lower panel) mice. IC-21 macrophages with or without internalized heat-killed Histoplasma yeasts were used as targets. Cytotoxicity was determined by LDH release assay. The data shown are representative of five separate experiments.
A
Medium Stimulated Medium Stimulated Infected Uninfected grB CD8 6.48 9.99 0.47 0.49 Isotype control CD8 0.08 0.14 0.13 0.07 B -10 -5 0 5 10 15 20 25 160:1 80:1 40:1 20:1 10:1 5:1 E:T ratio % cytotoxicity .
Infected CD8, target with Ag Infected CD8, target w/o Ag Uninfected CD8, target with Ag Uninfected CD8, target w/o Ag WT -10 -5 0 5 10 80:1 40:1 20:1 E:T ratio % cytotoxi c it y PKO
FIGURE 4. Histoplasma infection induces CD8 T cell IFNγ production. Spleen
cells were harvested from infected or uninfected WT mice and cultured in medium alone or medium containing live Histoplasma yeast cells for 24 h. The splenocytes were stained for intracytoplasmic IFNγ. The numbers at the upper-right corner are the percentages of CD4 or CD8 T cells that are producing IFNγ (CD4+IFNγ+/CD4+ or CD8+IFN γ+/CD8+). The data shown are from one of eight independent experiments.
Medium Stimulated Medium Stimulated Infected Uninfected CD8 0.00 0.00 3.68 0.14 CD4 0.03 0.00 5.39 0.09 IFNγ