~ Pergamon
0305-0491(94)00073-5
Comp. Biochem. Physiol. Vol. 109B, No. 1, pp. 81 88, 1994 Copyright © 1994 Elsevier Science Ltd Printed in Great Britain. All rights reserved 0305-0491/94 $7.00 + 0.00
Molecular cloning of the common carp
(Cyprinus carpio) rhodopsin cDNA
Huai-Jen Tsai, Su-Ru Shih, Ching-Ming Kuo and Lu-Ku Li*
Institute of Fisheries Sciences, National Taiwan University, Taipei, Taiwan 106; and
*Department of Ophthalmology: Research, Health Sciences, Columbia University, New York, U.S.A.
A recombinant phage clone containing a 1584 nucleotides rhodopsin cDNA was screened from a carp retinal cDNA library. The inserted DNA consisting of a single open reading frame of 1062 nucleotides at positions 72 to 1133 encodes a 354 amino acid polypeptide. The deduced amino acid sequence of carp rhodopsin showed 95.7, 85.5 and 74.4% identity with that of goldfish, sand goby and lamprey, respectively. The sites of palmitoylation, glycosylation, disulfide bond formation and Schiff base formation in the putative rhodopsin are all conserved.
Key words: Carp; PCR screening; Photoreceptor; Retinal cDNA library; Rhodopsin; Signal transduction; Transmembrane domain; Visual pigment.
Comp. Biochem. Physiol. 109B, 81-88, 1994.
Introduction
The underwater environment limits both the intensity and the spectral bandwidth of ambient light for vision and aquatic sur- vival (Lythgoe, 1980). Yet retina anatomy and photoreceptor proteins (opsins) show structural similarities between land and aquatic vertebrates (Yokote, 1982; Nathans et al., 1986). Of the photoreceptor proteins, rhodopsin is the most completely investi- gated GTP binding protein in terms of structure and function relationship; it be- longs to the family of seven-membrane- span receptors such as the fl-adrenergic receptor (Ferretti et al., 1986; Stryer, 1991; Khorana, 1992). Based on single cell elec- trophysiological measurements, a rod and Correspondence to: H u a ; - J e n Tsai, Institute o f Fish- eries Sciences, N~ ,onal T a i w a n University, Taipei, T a i w a n 106.
Received I1 J a n u a r y 1994; accepted 9 M a y 1994. 81
three cone cell photoreceptors have been suggested (Tomita et al., 1967). Recently several opsin cDNAs of the goldfish, a teleost that occupies a more surface habitat have been reported (Johnson et al., 1993). In order to further understand the com- parative aquatic visual physiology we have initiated a study on the opsins of the com- mon carp, Cyprinus carpio, an important aquacultured species and a closely related teleost with goldfish but occupies a more bottom habitat.
In this paper, we report the isolation and characterization of a putative carp rhodopsin cDNA clone from the retina of the cultured carp. A comparison of the nucleotide sequence with those reported for goldfish, sand goby and lamprey indicates that factors other than phylogeny may be also important in determining sequence relatedness.
82 Huai-Jen Tsai et al.
Materials and Methods
Isolation of RNA and eDNA library con- struction
Retinae were freshly dissected from 25 carps (300-500 g each, a gift from Dr I- Chio Liao, and Fu-Guang Liu, Taiwan Fisheries Research Institute, Keelung). After grinding in a mortar in liquid nitro- gen, the homogenate was suspended in prechilled denaturing solution containing 4 M guanidinium thiocyanate, 0.51% N-laurylsarcosine, 0.125 mM fl-mercapto- ethanol, 5/tg/ml polyvinyl sulfate, and 26.25 m M sodium citrate; this suspension was then drawn 5-6 times with a syringe fitted with an 18-gauge needle. Total retinal R N A was prepared by using the acid/phenol method described by Chom- czynski and Sacchi (1987), with an ad- ditional gel filtration through a PD-10 column (Pharmacia) to remove melanin. The poly(A) + RNA was isolated and the retinal c D N A library was then constructed in a lambda gtl0 (Promega) using Es- cherichia coli VC257 as the host cells as previously described by Tsai et al. (1993). Screening by polymerase chain reaction (PCR)
The initial screening of the c D N A library (430,000 phages divided into ten equal fractions) was achieved by PCR using a synthetic 27-mer, 5'-GGCTGGAGCAG- G T A T A T C C C G G A G G G T - 3 ' as the for- ward primer. It corresponds to the nucleotide positions 731 to 757 of the Drosophila rhodopsin e D N A and is hom- ologous with bovine sequence (Zuker et al., 1985). This sequence mimics the conserved rhodopsin intradiscal segment involved in disulfide and tertiary structure formation (Khorana, 1992). The reverse primer, 5'- G A G G T G G C T T A T G A G T A T T T C T T C - CAGGGTA-3', was derived from lambda gtl0 (Promega).
The PCR procedure was carried out in a 100#1 solution containing 9 x 106 phage D N A as template, 20 pmol reverse primer, 6pmol forward primer, 2 0 0 # M of each dNTP, 10 # g of bovine serum albumin, and 5 U Taq polymerase with 10#1 of 10x buffer (Promega). A D N A Thermal Cycler (Perkin-Elmer Cetus, Norwalk, CT) was used for PCR amplification. The tempera-
ture profiles were: 94°C for 1 min, 68°C for 2 min, and 72°C for 3 min for the first two cycles; 94°C for 1 min, 65°C for 2 min, and 72°C for 3 rain for a second three cycles; and 94°C for 1 rain, 60°C for 2 min, and 72°C for 3 rain for the final 30 cycles. The resulting PCR products were analyzed by 0.8% agarose gel electrophoresis.
Plaque hybridization screening
Those phage fractions generating the most intense PCR products of the predicted sizes were screened with a goldfish rhodopsin c D N A probe. This probe, mol- ecular size 1.3 kb, was isolated from an amplified plasmid pRJ1 (a gift from Dr. Kathy Grant of the Department of Chem- istry, Columbia University, NY) followed by Eco RI digestion. Approximately 75,000 recombinant phages were plated on three NZY (0.5% NaC1, 0.2% MgSO4"7H20, 0.5% yeast extract and 1% N Z amine) plates; they were then transferred to nitro- cellulose membranes by blotting. Hybridiz- ation with the 1.3 kb 32P-labeled goldfish probe was carried out in a buffer (10% dextran sulfate, 1% SDS, 1 M NaC1, 5 0 m M Tris-HC1; pH 7.5) at 42°C overnight. The membranes were washed twice (20 min each) with a solution contain- ing 0.5% SDS and 6 x (0.15M NaCI, 1 mM EDTA, and 30 mM Tris-HC1; pH 8) at 27°C. Positive clones were identified via exposure to X-ray film (Kodak) at - 7 0 ° C for two days, then amplified in a second screening.
The hybridized nitrocellulose membranes from the second screening were washed three times (20 rain each) in 0.5% SDS and 6 x SSC (1 x SSC: 0.15M NaC1 and 0.015 M sodium citrate) at 27°C, followed by another two washings in 0.1% SDS and 2 x SSC at 42°C, plus a final washing in 0.05% SDS and 1 x SSC at 65°C for 10 min.
Southern blotting
Recombinant D N A inserts prepared from the positive clones after Eco RI diges- tion were analyzed by Southern blotting (Southern, 1975). The probe used was the 1.3kb goldfish rhodopsin c D N A radio- labeled by the random priming (Feinberg and Vogelstein 1983).
Carp rhodopsin eDNA 83
Subcloning
The cDNA insert isolated from the 0.8% agarose gel was ligated onto an Eco RI digested phagemid pGEM-7ZF( + ), (Promega). The transformation of E. coli DH5ct was carried out following the pro- cedures described by Hanahan (1985). The transformants harboring the cDNA inserts were screened via plasmid DNA mini-prep- aration analysis (Sambrook et al., 1987). DNA sequencing
Our strategy for determining the nucleo- tide sequences of the isolated carp retinal cDNA is shown in Figure 1. We used the dideoxynucleotide chain-termination method (Sanger et al., 1977) with a Sequenase kit (US Biochemical Corp., Cleveland). In addition to the T7 and SP6 primers, four sequence extension oligomers were synthesized: primer CRH1, +188 5 ' - C C T G G C C G C G T A C A T G T T C - 3 ' + 206 sense strand; primer CRH2, + 1345 5'-GGGACATGGTCTGTC-Y + 1331 an- tisense strand; primer CRH3, +536 5'- GGGGGTTGTCTTCAC-Y +551 sense strand; and primer CRH4, +1034 5'- CAGGGTGGTGATCATGC-3' +988 an- tisense strand (numbers refer to the nucleotide position on the carp eDNA sequence).
Sequence analysis
Both the nucleotide and predicted amino acid sequences of the isolated carp rhodopsin eDNA were compared to those of other fishes deposited with the GenBank and National Biomedical Research Foun-
dation data bases. To align these sequences, we introduced gaps to maximize identity via procedures described by the Genetic Com- puter Group (Higgins and Sharp, 1988). Hydropathicity profile of carp rhodopsin amino acid sequence was obtained accord- ing to analyses described by Kyte and Doolittle (1982). In addition, we used a Chargpro program from the PC Gene soft- ware package for estimating the isoelectric point (pI) of carp rhodopsin.
R e s u l t s and D i s c u s s i o n
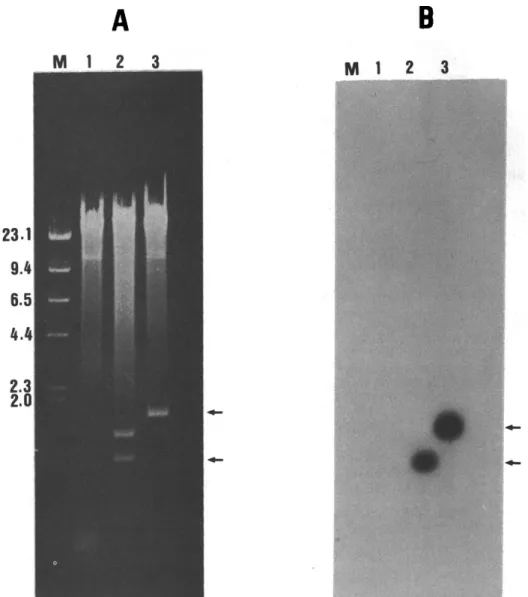
Identification of the putative rhodopsin clone The initial PCR screening of the carp retinal eDNA library revealed bands of 0.4-0.6kb in five of the 10 fractions on the agarose gel (Fig. 2). Since the forward primer corresponds approximately to the middle segment (see Materials and Methods) of the rhodopsin sequence, one fraction (lane 2, Fig. 2) containing the longest PCR product was chosen for plaque hybridization screening. Twenty-four posi- tive recombinant phage clones were isolated following the first screening; seven clones were purified via a second plaque screening, and the inserted DNA from three of these clones was digested with EcoRI (Fig. 3A) and analyzed by Southern blot- ting (Fig. 3B). Two inserts of approximately 0.9 and 1.6 kb showed strong hybridization with the goldfish probe (lanes 2 and 3, Fig. 3B). The 1.6 kb insert (lane 3, Fig.3) was constructed in plasmid pGEM-7Zf( + ) and transformed into host E. coli, then sequenced. SP6 / ! I Sp Sa Ac Ba Ac
A
I
F"///////////////////////~///~////////////////////////////////////A
"72 1134 A ! 198 c 57; o J 5~ 536 E F lO3Fig. 1. A partial restriction map and sequencing strategy of carp rhodopsin eDNA. The untranslated region (empty box) and coding region (hatched box) of the eDNA are shown. DNA fragments A, B, C, D, E and F were sequenced by using primers SP6, T7, CRHI, CRH2, CRH3, and CRH4, respectively. See Materials and Methods for details. Ac, AccI; Ba, BamHI;
Sp, SphI; Sa, SmaI restriction site.
I 12~9 B 158Z I 1345 T7 I
84
M 1 1 2
Huai-Jen Tsai et al.
3 4 5 6 7 8 9 10 M2
Fig. 2. Analysis of PCR products. PCR products from the carp retinae c D N A library divided into 10 43,000 phages fractions were shown in lanes 1 to 10. The molecular markers, M1 and M2, were HindIII-digested lambda genome and HaeIII-digested q~X-174-RF DNA,
respectively.
Nucleotide sequence of carp rhodopsin cDNA and comparison with that of other known genes
The nucleotide sequence of carp rhodopsin c D N A is shown in Fig. 4. The putative carp rhodopsin c D N A sequence contains a single open reading frame of 1062 nucleotides located between nucleo- tide positions 72 and 1133, which encodes 354 amino acid residues. The translation initiation codon (ATG) was observed at nucleotide positions 72 to 74; it was pre- ceded by 71 untranslated leader sequences. The translation termination codon TAA, nucleotide positions 1134 to 1136, was fol- lowed by an untranslated region of 448 nucleotides at the 3' end. An AATAAA hexanucleotides, the polyadenylation sig- nal, starting at nucleotide position 1511, was 15 nucleotides upstream from the start- ing site of the poly (A) tail of 52 As.
The carp c D N A was identified as that of a putative rhodopsin sequence on basis of its 93.4% nucleotide homology with the coding region of that of the goldfish rhodopsin (Johnson et al., 1993). In con- trast, it showed a low respective homology of 56.0, 61.8, 63.4 and 65% with that of goldfish red, blue, green-1 and green-2 opsin. Compared with other known fish
rhodopsins it exhibits 80.1 and 73.4% iden- tity with that of sand goby (Archer et al., 1992) and lamprey (Hisatomi et al., 1991) respectively, suggesting a more distant phy- logeny with the latter. It is interesting to note that the carp rhodopsin also showed 72.6, 72,8 and 74.3% identity with that of human (Nathans and Hogness, 1984), mouse (A1-Ubaidi et al., 1990) and chicken (Takao et al., 1988) respectively. These results suggest that other factor(s) in ad- dition to phylogeny may also determine the extent of sequence similarity.
Comparison of the deduced amino acid se- quences of carp rhodopsin with those of other known fish species
Figure 5 shows a comparison of our deduced amino acid sequence in carp rhodopsin with those reported for other fish species. Identity was found to be 95.7% with that of goldfish (Johnson et al., 1993), 85.4% with the sand goby (Archer et al., 1992) and 74.4% with the lamprey (Hisatomi et al., 1991).
Membrane protein topology. The deduced amino acid sequence of the carp rhodopsin differs from that observed for goldfish in only 16 out of 354 residues; 10 of the 16 are homologous replacements for
Carp rhodopsin cDNA 85
A
B
M 1 2 3 M 1 2 3
Fig. 3. Southern blot analysis of the DNA insert isolated from the lambda gtl0 clones. (A) Ethidium bromide staining of genomic DNA from three positive clones extracted and digested with Eco RI; (B) autoradiogram of the nitrocellulose membrane hybridized with a 32p-labeled goldfish rhodopsin cDNA probe. Lane M shows the molecular markers of HindlII-digested lambda genome in kb, and lanes 1-3 show the recombinant phages. Arrows indicate the positively hybridized 0.9. and 1.6 kb bands. In this study, the 1.6 kb clone was used for
sequencing. residues having similar properties. The ob- served carp sequence encodes a largely hydrophobic polypeptide with a predomi- nantly hydrophobic N-terminal, while the C-terminal loop contains a 12 Ser/Thr seg- ment which is essential for the phosphoryl- ation control of visual signal transduction. The similarities observed in the hydropathy plots (data not shown) of carp and bovine rhodopsin suggest similarities in the transmembrane segmentation, i.e., repeat- ing segment of 19-27 residues (Helix I-VII in Fig. 5), as well as in high proline and aromatic amino acid contents.
Conserved amino acid residues. Lys-296
and Glu-113 are conserved for the retinal binding and counterion interaction required for Schiff base formation (Nathans, 1987). Disulfide between Cys-110 and Cys-187 is required for the formation of the retinal pocket from the seven mem- brane helical segments. The N-terminal Asn-2 and Asn-15, corresponding to the glycosylation sites of the Asn-X-Thr sequences, and the C-terminal Cys 322-Cys 323, the palmitoylation site essential to membrane anchoring, are completely conserved.
8
=
<8 S~o~ c~c.o SS
"i
~E r..,jr..~ r~ S~ S
~2
,-.a 8- ~J r~ [--. r~ ~z ~J83
i
i
i
oo
B S S S S~S~S o~~ d ~ N~ ~~~.
~°~=o 8~~
~
~..~
~'~
_e~<
gSe<~
-~< Z---Carp rhodopsin cDNA 87 CP MNGTEGPMFYVPMSNATGVVKSPYDYPQYYLVAPWAYGCLAAYMFFLIITGFPINFLTLY 6 0 GF . . . D . . . [ . R . . . A . . . V . . . 6 0 S G . . . F . . I ~ . V . T . . I . R . . . E . . . N . A . . A A . . . L . . . 6 0 LA . . . DN . . . . F . . K . . L A R . . . E . . . A E . . K . S A . . . L V . . . V . . . F 6 0 II C P V T I E H K K L R T P L N Y I L L N L A I S D L F M V F G G F T T T M Y T S L H G Y F V F G R I G C N L E G F F A T L G 1 2 0 G F . . . V . . . P . . . 1 2 0 S G . . L . . . A . . L . . . V A . . . M ... L . . L . . . V . . . 1 2 0 L A . . V Q . . . M A N .... L F . . . V ... M N . . . PTM.SI . . . 1 2 0 III IV C P G E M G L W S L V V L A F E R W M V V C K P V S N F R F G E N H A I M G V V F T W F M A C T C A V P P L V G W S R Y I P 1 8 0 GF . . . 180 S G . . I A . . . V . . . V ... IS .... T . . . A . S . I . . A . . . 1 8 0 L A . . V A . . . I . . Y I . I . . . M G ... N T ... A . . . I . . L A . . A . . . 1 8 0 V C P EGMQCSCGVDYYTRAPGYNNESFVIYMFLVHFIIPLIVIFFCYGRLVCTVKDAAAQQQES 240 GF . . . PQA . . . ! . . . E . . . . HE.. 240 S G . . . E . F . . . I . . . L A . . . L . A . . E . . . A .... 2 4 0 LA . . . P . . . . LN.NF . . . . Y . V . . . V . . . L V . F V I . . . L . . . . E . . . A . . . . 240 VI V~ CP ETTQRAEREVTRMVVIMVIGFLICWIPYASVAWYIFTHQGSEFGPVFMTVPAFFAKSAAV 300 G F . . . L . . . T . . . 3 0 0 S G . . . ! . . . T S . L . . . T .... L . . . I . . . S . L 3 0 0 LA AS..K..K . . . L . . . V..V . . . F . . . D . . A T . . . L . . . S.L 300 C P Y N P C I Y I C M N K Q F P , H C M I T T L C C G K N P F E E E E G A S T T A S K T E A S S V S S S S V S P A 3 5 4 G F . . . 354 SG ...M . . . - - . . . . 352 LA . . . V . . . L . . . N . . . LGDD.SGAS.- . . . . V . . . . T.P . . . . 353
Fig. 5. Comparisons of the predicted amino acid sequences of carp (CP) rhodopsin with that of goldfish (GF) (Johnson et al., 1993), sand goby (SG) (Archer et al., 1992) and lamprey (LA) (Hisatomi et al., 1991). Numbers begin at the first amino acid residue; identical amino acid residues are indicated by dots. Dashes represent gaps created to maximize the degree of identity among all compared sequences. The seven transmembrane domains are labeled I to
VII above the sequence.
N e t charges a n d p I. T h e c h a r g e c h a r a c t e r - istics o f t h e p u t a t i v e r h o d o p s i n w e r e e s t i m a t e d b a s e d o n t h e f o l l o w i n g p K v a l u e s : a r g i n i n e , 12.48; lysine, 10.79; h i s t i d i n e , 6; a s p a r a t a t e , 3.65; g l u t a m a t e , 4.25; c y s - CBP(B) 109/I---G teine, 8.35; a n d t y r o s i n e , 10.13. A v a l u e o f 6.3 w a s o b t a i n e d f o r p I o f t h e p r e d i c t e d c a r p r h o d o p s i n . T h i s is in t h e r a n g e o f 5 . 9 - 6 . 3 o b s e r v e d f o r b o v i n e a n d h u m a n r h o d o p s i n ( O k a n o et ai., 1992), b u t o n e
88 Huai-Jen Tsai et al.
which differs markedly from the alkaline pI values of approximately 9.5 observed for color and/or photopic cone pigments (Okano et al., 1992).
Acknowledgements--We thank Mr Jean-Leon Chong for help in some of the sequencing work. This work was supported by a grant from the National Science Council, Republic of China (NSC-82-0211-B 002-003).
References
AI-Ubaidi M. R., Pittler S. J., Champagne M. S., Triantafyllos J. T., McGinnis J. F. and Baehr W. (1990) Mouse opsin: gene structure and molecular basis of multiple transcripts. J. biol. Chem. 265, 20563-20569.
Archer S. S., Lythgoe J. N. and Hall L. (1992) Rod opsin cDNA sequences from the sand goby
(Pomatoschistus minutus) compared with those of other vertebrates. Proc. Royal Soc. Lond. Ser. B, 248, 19-25.
Chomczynski P. and Sacchi N. (1987) Single-step method of R N A isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal.
Biochem. 162, 156-159.
Feinberg A. P. and Vogeistein B. (1983) A technique for radiolabeling D N A restriction endonuclease fragments to high specific activity. Anal. Biochem. 132, 6-10.
Ferretti L., Karnik S. S. and Khorana H. G. (1986) Total synthesis of a gene for bovine rhodopsin.
Proc. natn. Acad. Sci. U.S.A. 83, 599~03. Hanahan D. (1985) Techniques for transformation of
E. coli. In DNA Cloning: A Practical Approach (Edited by Glover D. M.), Vol. 1, pp. 109-135. IRL Press, Oxford.
Higgins D. G. and Sharp P. M. (1988) CLUSTAL: a package for performing multiple sequence align- ment on a microcomputer. Gene 73, 237-244. Hisatomi O., Iwasa T., Tokunaga F. and Yasui A.
(1991) Isolation and characterization of lamprey rhodopsin cDNA. Biochem. biophys. Res. Com-
mun. 174, 1125-1132.
Johnson R. L., Grant K. B., Zankel T. C., Boehm M. F., Merbs S. L., Nathans J. and Nakanishi K. (1993) Cloning and expression of goldfish opsin sequences. Biochemistry 32, 208-214.
Khorana H. G. (1992) Rhodopsin, photoreceptor of the rod cell. J. biol. Chem. 267, 1-4.
Kyte J. and Doolittle R. F. (1982) A simple model for displaying the hydropathic character of a protein.
J. molec. Biol. 157, 105-132.
Lythgoe J. N. (1980) Vision in fishes. In Environmen-
tal Physiology o f Fishes (Edited by Ali M. A.), pp. 431-445. Plenum Press, New York.
Nathans J. (1987) Molecular biology of visual pig- ments. Ann. Rev. Neurosci. 10, 193-194.
Nathans J. and Hogness D. S. (1984) Isolation and nucleotide sequence of the gene encoding human rhodopsin. Proc. natn. Acad. Sci. U.S.A. 81, 4851--4855.
Nathans J., Thomas D. and Hogness D. S. (1986) Molecular genetics of human color vision: the genes encoding blue, green and red pigments.
Science 232, 193-202.
Okano T., Kojima D., Fukata Y., Schichida Y. and Yoshizawa T. (1992) Primary structure of chicken cone visual pigments. Proc. natn. Acad. Sci. U.S.A. 89, 5932-5936.
Sambrook J., Fritsch E. F. and Maniatis T. (1987) Small-scale preparations of plasmid DNA. In
Molecular Cloning: A Laboratory Manual, second edition, pp. 1.25-1.32. Cold Spring Harbor Lab- oratory Press, Cold Spring Harbor.
Sanger F., Nicklen S. and Coulson A. R. (1977) DNA sequencing with chain terminating inhibitors. Proc.
natn. Acad. Sci. U.S.A. 74, 5463-5467.
Southern E. (1975) Detection of specific sequences among D N A fragments separated by gel electro- phoresis. J. molec. Biol. 98, 503-517.
Stryer L. (1991) Rhodopsin-fl adrenergic receptor.
J. biol. Chem. 266, 10711-10714.
Takao M., Yasui A. and Tokunaga F. (1988) Iso- lation and sequence determination of the chicken rhodopsin gene. Vision Res. 28, 471-480. Tomita T., Kaneko A., Murakami M. and Pautler
E. LI (1967) Special response curves of single cones in the carp. Vision Res. 7, 519-528.
Tsai H. J., Lin K. L. and Chen T. T. (1993) Molecular cloning and expression of yellowfin porgy
(Acanthopagrus latus Houttuyn) growth hormone cDNA. Comp. Biochem. Physiol. 104B, 803-810. Yokote M. (1982) Sensory organs: Eye. In An Altas
ofFish Histology (Edited by Hibiya T.), pp. 42-47. Gustav Fischer, Stuttgart.
Zuker C., Cowman A. F. and Rubin G. M. (1985) Isolation and structure of a rhodopsin gene from