行政院國家科學委員會補助專題研究計畫
□ 成 果 報 告
■期中進度報告
克雷白氏肺炎桿菌中 Fur 的功能性分析
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 97-2314-B-039-042-MY2
執行期間: 97 年 12 月 1 日至 99 年 7 月 31 日
計畫主持人:林靖婷
共同主持人:
計畫參與人員: 陳育聖
成果報告類型(依經費核定清單規定繳交):■精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
中文摘要:
關鍵字:螯鐵系統、莢膜多醣體、克雷白氏肺炎桿菌、Fur、FURTA 莢膜多醣體 (capsular polysaccharides, CPS) 和螯鐵系統都是克雷白氏肺炎桿菌 (Kp) 的重 要致病因子。大部分的 Kp 臨床菌株會產生厚實的 CPS 來幫助細菌躲避免疫細胞及血清分 子的攻擊,以及產生多組的螯鐵系統來獲得環境中的鐵,以幫助其生長和感染宿主。在革 蘭氏陰性菌中,螯鐵系統的基因表現會受到 Fur 所抑制。然而在克雷白氏肺炎桿菌中除了 證實 Fur 可以自我調控外,對於其他的基因調控方面的研究非常的少,因此為了研究 Fur 在 Kp 上的功能,我們已經在具有高毒性 K2 血清型的 Kp CG43 中構築了 fur 的缺損株 ; 其 中發現 fur 的缺損會造成 CPS 的大量生成並提高 Kp 對於小鼠的毒性,因此推測 Fur 在毒 性因子的調控上扮演重要的角色。為了研究 Fur 在 cps 和螯鐵系統的基因調控上的角色, 過去六個月我們已經完成下列的幾項實驗: 1. 我們已經構築好了 fur 的互補質體,並且以轉殖入 fur 的缺損株中觀察其顯型的變化, 其中發現 fur 的缺損株會減緩 Kp 的沈澱速度並且減低生物膜的生成,然而 fur 的互補株 則可恢復其顯型表現。 2. 我們構築了 Kp 中 K1 和 K2 的 cps 基因調控組的啟動子在 pT7-7 中,並且利用 FURTA 來 偵測大腸桿菌的 Fur 對於 K1 和 K2 的cps 基因調控組的啟動子之結合能力。初步的結果 發現這些啟動子在 FURTA 的實驗中皆呈現陰性反應,所以大腸桿菌的 Fur 不會直接結合 到這些啟動子上進行調控反應。 3. 在 cps 的調控迴路中,我們也構築 rcsA 和 rcsB 的啟動子在 pT7-7 中,並且利用 FURTA 來偵測大腸桿菌中 Fur 對於這兩各啟動子的結合能力,其結果也是呈現陰性反應,因此 我們將近一步構築並偵測在 cps 的調控迴路中其他調控子的啟動子對於大腸桿菌 Fur 的 結合親和力。 4. 利用生物資訊所分析出來在螯鐵系統中含有 Fur 結合區域的啟動子,我們進一步構築並 利用 FURTA 來偵測 Fur 對於這些預測有 Fur 結合區域的啟動子的結合能力,其中發現 只有 iroN、iroB、entC、 ybtP、 fecB 和 kfuA 顯示為典型的 FURTA 陽性反應,其他基 因雖然有 Fur 的結合區域但是卻呈現陰性反應。 在本年度,我們已經藉由完成fur 的互補質體的構築完成來確認 Fur 對於 Kp 中沈澱速率及 生物膜生合成的專一性影響,並且發現大腸桿菌的 Fur 並不會直接結合至 K1 或 K2 的cps 基因啟動子區域進行調控動作,而在 cps 的調控迴路中,rcsAB 的啟動子也不會受到 Fur 所結合調控。而在螯鐵系統中,我們發現並非所有帶有 Fur 結合區域的啟動子都能順利的 受到 Fur 所調控,因此之後我們將利用生物資訊方式比較 Fur 固定結合區域的序列並且利 用 FURTA 和電泳交遲滯實驗証實 Fur 的結合能力。下年度,我們會進一步利用二維電泳膠 和即時聚合酶連鎖反應來找尋 Fur 在莢膜多醣體生合成中所調控的下游基因,綜合以上幾 點,我們希望能夠瞭解 Fur 在克雷白氏肺炎桿菌的致病過程中所扮演的角色。Abstract:
Key words
:
Iron-acquisition system, capsule polysaccaharides, Klebsiella pneumoniae, Fur, FURTAIron-acquisition system and capsule polysaccaharides (CPS) are important virulence factors in Klebsiella pneumoniae (Kp) infection. Most clinical isolated of Klebsiella strains produce large amounts of to protect the bacteria from phagocytosis and prevent from killing by serum factors, and develop several iron-acquisition systems for sequestering ferric iron from the environment to promote the growth and infection. In Gram-negative bacteria, iron-acquisition genes are negatively regulated by ferric uptake regulator (Fur). Due to the regulatory roles of Fur in Kp remains large unknown. We have demonstrated that the fur deletion mutant rendered the bacteria more mucoid phenotype and slight reduced the LD50 using mouse lethality assay suggesting an
involvement of Fur in virulence regulation. To study the functional role of Fur in mediating cps and iron-acquisition system gene clusters, we had been finish the experiments in the past six month are:
1. The construction of the fur complement strain was performed. The fur deletion mutant slowed the Kp precipitation rate and reduced biofilm formation, while the fur complement stain could restore the phenotype.
2. The putative promoter regions of K1 and K2 cps gene cluster were constructed and detected the E. coli Fur binding affinity by Fur titration assay (FURTA). The preliminary results showed that all the cloned promoter of either K1 or K2 cps gene cluster showed FURTA-negative phenotype.
3. The promoter regions of rcsA and rcsB in cps regulon were constructed and detected by FURTA, the results exhibited FURTA-negative phenotype implying RcsAB is not involved in Fur regulating.
4. The iron-acquisition systems containing the putative Fur boxes were constructed and detected by FURTA. The cloned promoters containing iroN, iroB, entC, ybtP, fecB, and kfuA showed typical FURTA-positive phenotypes. However, the clones containing the promoter fepA, fepB, fyuA, and fhuA exhibited negative phenotypes.
In next year, we will employ the 2D and real-time PCR to demonstrate the regulatory roles of Fur in CPS biosynthesis. However, the typical Fur box in Kp will be further analyzed and demonstrated by bioinformatics, FURTA, and EMSA. Take together, we hope to understand the functional role of Fur in Kp pathogenesis.
Introduction:
Klebsiella pneumoniae (Kp) is a common nosocomial pathogen. As an opportunistic pathogen, it often causes pneumonia, septicemia, bacteriemia, suppurative lesion, wound infection, burn infection, and urinary or respiratory tract infections in immunocompromised patients [1-4]. However, the wide spread of extended spectrum lactamase producing K. pneumoniae (ESBLKp) has prompted the search for new drugs to intervene the bacterial infections [5, 6].
Klebsiella usually have well developed polysaccharide capsules, which make their colonies characteristic mucoid. CPS is an important virulence factor in Kp infection to protect the bacteria from phagocytosis and prevent from killing by serum factors [7, 8]. Apart from their antiphagocytic function, Klebsiella CPS have been reported to inhibit differentiation and functional capacity of macrophage in vitro and mediate the resistance to antimicrobial peptides and proteins by limiting the interaction of the agents with membrane targets [9, 10]. The capsular serotypes of Kp have been classified as more than 77 recognized capsular antigens [11, 12]. In Taiwan, a high prevalence of K1 and K2 serotypes of Kp was documented in liver abscess of diabetes mellitus patients [13].
Many bacteria have evolved iron-acquisition systems for sequestering ferric iron from the environment. The iron/heme acquisition systems in bacteria can be separated into two general mechanisms. One is a direct contact between the bacteria and the exogenous iron/heme sources. Another one is compounds (siderophores and hemophores) synthesized and released by bacteria into extracellular medium to scavenge iron or heme from various sources [14]. The transport of these compounds across the outer membrane involves outer membrane receptors. The energy for siderophore transport is delivered from the inner membrane by a TonB-ExbB-ExbD complex [15]. Analysis of the genomic sequence of Kp NTUH-K2044 have found ten putative iron-acquisition systems, seven of ten putative iron-acquisition systems contain putative TonB-dependent domains [16].
To avoid iron overloading that would lead to the formation of hydroxyl radicals, microorganisms have tight regulation of iron uptake. In Gram-negative bacteria, iron-acquisition genes are negatively regulated by ferric uptake regulator (Fur) [17-19]. Under iron-replete conditions, the Fur protein functions as a dimmer with Fe2+ as cofactor to bind to a 19-bp consensus sequence, called the Fur box (GATAATGATwATCATTATC; w=A or T), in the promoters of genes requires for iron uptake, thereby preventing their transcription [20]. In Kp, the functions of Fur on mediating the gene expression remains large unknown. However, we further have constructed the fur deletion mutant in Kp CG43S3 to observe Fur function in Kp. Interestingly, the deletion rendered the bacteria more mucoid phenotype and reduced the LD50
implying Fur has diverse regulatory functions on mediating CPS and iron-acquisition system. In order to demonstrate the regulatory functions of Fur, the experiments we have finished in the past six month are as following:
1. Investigation of regulatory roles of Fur on mediating the cps expression in Kp;
deletion mutant and the complement strain in the viscous phenotype, biofilm formation, and CPS content were performed.
(2.) We have constructed the cps promoter region containing K1 and K2 serotype of Kp in pT7-7 vector respectively, and transferred into H1717 (fhuF::lacZ)to detect the direct binding affinity of Fur by Fur titration assay (FURTA).
(3.) The promoter region of RcsA and RcsB were also performed the construction in pT7-7 vector and transferred into H1717 to detect the direct binding affinity of Fur by FURTA. 2. Investigation of regulatory roles of Fur on mediating the gene expressions of iron-acquisition
systems in Kp;
(1.) The promoter regions of five putative iron-acquisition systems containing the putative Fur boxes in Kp NTUH-K2044 by bio-informatics analysis were constructed in pT7-7 vector and transferred into H1717. FURTA was also performed to analysis the Fur direct binding affinity.
Results and discussions:
The phenotype of fur deletion mutant in Kp CG43. In this study, the fur complement strain was
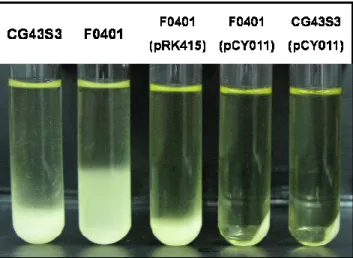
performed the construction and compared with wild type (CG43S3), fur deletion mutant (F0401), and fur complement strain (the entire fur gene in pRK415, named pCY011) in the viscous phenotype, CPS content, and biofilm formation. During low-speed centrifugation, the fur mutant strain exhibited much slower precipitation than the parental strain. While the fur mutant and wild type stain supplied with the plasmid pCY011 respectively, revealed that faster precipitation via low-speed centrifugation (Fig. 1). Subsequently, the CPS production in CG43S3, F0401, and F0401 containing pCY011 will be quantified by measuring the glucuronic acid content, the core component of the K2 CPS. The production of extracellular polysaccharides in bacteria is important for the maturation of biofilms. In Kp, the interaction of CPS and fimbriae was able to modulate the bacteria biofilm formation [21]. Therefore, the biofilm formations in CG43S3, F0401, and F0401 (pCY011) were detected in polyvinylchloride (PVC) plates by staining 1% crystal violet, which could be quantitatively measured at 595 nm for the absorbance. As shown in Fig. 2, the biofilm formation was significantly lower in F0401 than in wild type, and supplied with the plasmid pCY011 restored the ability of biofilm synthesis. These results implied that Fur is involved in the regulation of CPS biosynthesis and affects the biofilm biosynthesis in Kp CG43.
The regulatory role of Fur in mediating cps gene expression in FURTA. To investigate the
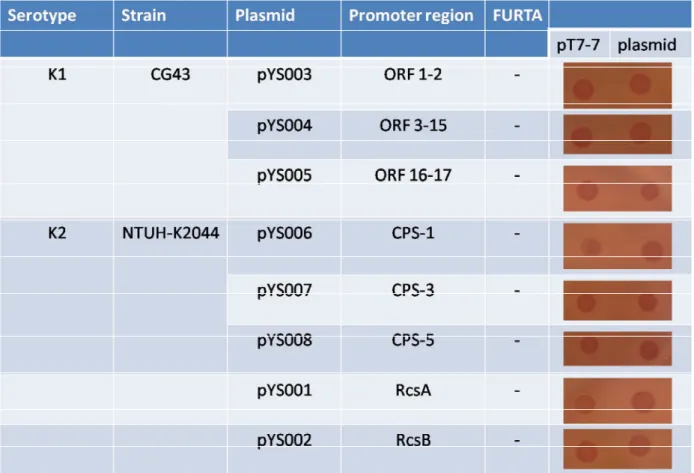
regulatory functions of Fur on mediating cps and iron-acquisition system gene expression, FURTA is a good reporter assay for examine the in vivo Fur binding function in the promoter regions of target genes. We asked for FURTA indicator strain, Escherichia coli H1717, from Dr Hantke to perform this study. H1717 contains a chromosome fhuF::lacZ fusion, and the low affinity Fur box in fhuF has demonstrated. The introduction of high-copy-number plasmids that contain Fur-binding sequences in the multiple cloning site causes removal of Fur from the fhuF Fur box [22]. The resultant expression of LacZ is visualized as lactose-positive colonies on MacConkey agar plates. In this study, the promoter regions of Kp K1 and K2 cps gene cluster were constructed in pT7-7 vector respectively and transferred into H1717 to assess the interaction of cps promoter regions with Fur. As a result, all the cloned promoter of either K1 or K2 cps gene cluster showed FURTA-negative phenotype, which suggests that non-direct binding of Fur on mediating the cps gene expression (Table 1). Furthermore, the clones carrying the promoters of rcsA and rcsB also exhibited FURTA-negative phenotype implying RcsAB is not involved in Fur regulating (Table 1). The concentration of ferric ammonium sulfate and the incubation time are critical for Fur regulating the target genes. However, the different concentration of ferric ammonium sulfate and different incubation time will be detected in this assay. The -galactosidase activity will be detected to quantify the binding activity.
The regulatory role of Fur in mediating iron-acquisition system gene expression in FURTA. To
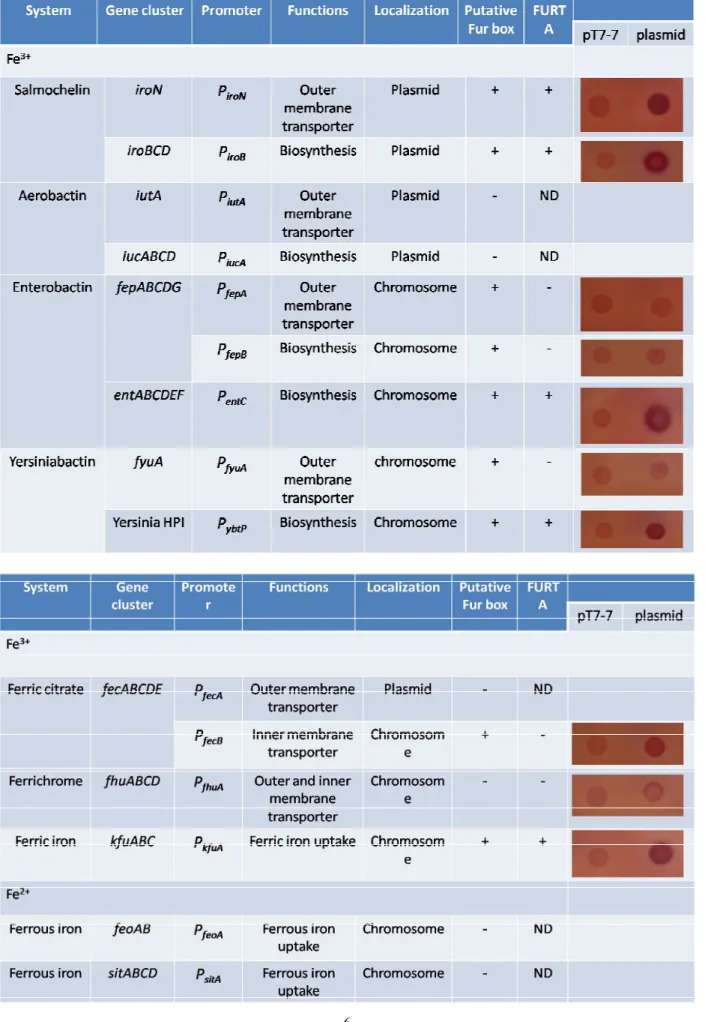
investigate the regulatory functions of Fur on mediating iron-acquisition system gene expression, five of ten putative iron-acquisition systems containing the putative Fur boxes were frist detected the direct binding affinity of Fur by FURTA. As showed in Table 2, the cloned promoters
containing iroN, iroB, entC, ybtP, fecB, and kfuA showed typical FURTA-positive phenotypes. However, the clones containing the promoter fepA, fepB, fyuA, and fhuA exhibited negative phenotypes. The results implied that Fur regulate the target gene is not dependent on the Fur box. We will attempt to align the promoter regions of iroN, iroB, entC, ybtP, fecB, and kfuA to find the conserve domain or residue for affecting the regulation of Fur. The -galactosidase activity will be detected to quantify the Fur binding activity.
Fig. 1 Comparison of precipitation speed of Kp wild type (CG43S3), fur mutant (F0401), and the strains carried the complement plasmid (pCY011).
Fig. 2 Biofilm formation of Kp wild type (CG43S3), fur mutant (F0401), and the strains carried the complement plasmid (pCY011). CG43S3-pRK415, CG43-pFur, △fur-pRK415,
and △fur-pFur (fur complement) strains were grown in LB broth at 37 ℃ for 48 h, and then measured OD550 with crystal violet strained.
Table 1 The binding ability of Fur on the promoter regions of K1 and K2 cps gene cluster
and rcsAB in FURTA. The promoters were constructed to pT7-7 in E. coli H1717, and 5 μl of
overnight grown bacteria were spotted on MacConkey agar plate containing 30 μM [Fe+3] for 16 h. + means positive result. – means negative results, and ND means not detectible.
Table 2 The classification of siderophores in K. pneumoniae and the Fur binding ability via FURTA. The promoters were constructed to pT7-7 in E. coli H1717, and 5 μl of overnight grown
bacteria were spotted on MacConkey agar plate containing 30 μM [Fe+3] for 16 h. + means positive result. – means negative results, and ND means not detectible.
參考文獻:
[1] F.F. Chou, H.K. Kou, J Am Coll Surg 182 (1996) 33-36. [2] S.H. Han, West J Med 162 (1995) 220-224.
[3] Y.J. Lau, B.S. Hu, W.L. Wu, Y.H. Lin, H.Y. Chang, Z.Y. Shi, J Clin Microbiol 38 (2000) 412-414.
[4] H.L. Peng, P.Y. Wang, J.L. Wu, C.T. Chiu, H.Y. Chang, Zhonghua Min Guo Wei Sheng Wu Ji Mian Yi Xue Za Zhi 24 (1991) 264-271.
[5] J. Wiener, J.P. Quinn, P.A. Bradford, R.V. Goering, C. Nathan, K. Bush, R.A. Weinstein, Jama 281 (1999) 517-523.
[6] E.N. Lagamayo, Am J Infect Control 36 (2008) S101-108.
[7] H. Sahly, R. Podschun, T.A. Oelschlaeger, M. Greiwe, H. Parolis, D. Hasty, J. Kekow, U. Ullmann, I. Ofek, S. Sela, Infect Immun 68 (2000) 6744-6749.
[8] J.C. Lin, F.Y. Chang, C.P. Fung, J.Z. Xu, H.P. Cheng, J.J. Wang, L.Y. Huang, L.K. Siu, Microbes Infect 6 (2004) 1191-1198.
[9] M.A. Campos, M.A. Vargas, V. Regueiro, C.M. Llompart, S. Alberti, J.A. Bengoechea, Infect Immun 72 (2004) 7107-7114.
[10] M.A. Campos, P. Morey, J.A. Bengoechea, Antimicrob Agents Chemother 50 (2006) 2361-2367.
[11] C.P. Fung, B.S. Hu, F.Y. Chang, S.C. Lee, B.I. Kuo, M. Ho, L.K. Siu, C.Y. Liu, J Infect Dis 181 (2000) 2075-2079.
[12] Y.J. Pan, H.C. Fang, H.C. Yang, T.L. Lin, P.F. Hsieh, F.C. Tsai, Y. Keynan, J.T. Wang, J Clin Microbiol 46 (2008) 2231-2240.
[13] C.P. Fung, F.Y. Chang, S.C. Lee, B.S. Hu, B.I. Kuo, C.Y. Liu, M. Ho, L.K. Siu, Gut 50 (2002) 420-424.
[14] C. Wandersman, P. Delepelaire, Annu Rev Microbiol 58 (2004) 611-647. [15] V. Braun, M. Braun, Curr Opin Microbiol 5 (2002) 194-201.
[16] P.F. Hsieh, T.L. Lin, C.Z. Lee, S.F. Tsai, J.T. Wang, J Infect Dis 197 (2008) 1717-1727. [17] J.F. Ernst, R.L. Bennett, L.I. Rothfield, J Bacteriol 135 (1978) 928-934.
[18] K. Hantke, Mol Gen Genet 182 (1981) 288-292. [19] L.A. Achenbach, W. Yang, Gene 185 (1997) 201-207. [20] D.W. Griggs, J. Konisky, J Bacteriol 171 (1989) 1048-1054.
[21] M.A. Schembri, J. Blom, K.A. Krogfelt, P. Klemm, Infect Immun 73 (2005) 4626-4633. [22] I. Stojiljkovic, A.J. Baumler, K. Hantke, J Mol Biol 236 (1994) 531-545.
計畫成果自評:
本計畫在六各月的執行過程中,順利的完成各項的基因構築的工作,其中 fur 的互補株的構 築完成也能進一步確認 fur 突變後對於 Kp 顯型方面的專一性影響,而在探討 Fur 對於 cps 基因啟動子調控上的影響,我們也順利跟 Dr. Hantke 拿到 H1717 來偵測大腸桿菌中 Fur 對 於所構築出來的啟動子序列的結合能力,因此我們分別將 K1 和 K2 的莢膜多醣體基因組的 啟動子和 rcsAB 的啟動子進行 FURTA 的分析,雖然目前發現皆為陰性反應,即沒有受到 Fur 直接結合所調控,但是我們會進一步針對不同鐵離子濃度以及構築其他與 cps 調控有關 的轉錄調控子的啟動子序列來進行更深入的探討。而對於螯鐵系統中,我們也發現在我們 先前分析十個具有 Fur 結合區域的啟動子中,只有六個在 FURTA 的實驗中呈現陽性反應, 因此我們將會進一步比較這六個陽性反應和其他四個陰性反應的啟動子在 Fur 結合區域的 相異性,整體而言,本計畫的預期目標我們也已經積極的完成,因此我們在未來的一年藉 由蛋白質體學和生物資訊能夠完成所規劃之目標,瞭解 Fur 在克雷白氏肺炎桿菌中所扮演 的調控角色。而在本人也將參與於今年七月二十六到三十一日於美國 Gordon research conference 所舉辦 的會議 microbial adhesion & signal transduction”。將目前所得到的實驗結果與之分享與討 論,期許能更深入的加以探討並且由各學者的經驗分享能夠得到更多的經驗和學習機會。