Phytomedicinexxx (2012) xxx–xxx
ContentslistsavailableatSciVerseScienceDirect
Phytomedicine
jo u r n al h om e p a g e :w w w . e l s e v i e r . d e / p h y m e d
Antileukemia
component,
dehydroeburicoic
acid
from
Antrodia
camphorata
induces
DNA
damage
and
apoptosis
in
vitro
and
in
vivo
models
Ying-Chi
Du
a,1,
Fang-Rong
Chang
a,b,c,d,1,
Tung-Ying
Wu
a,
Yu-Ming
Hsu
a,
Mohamed
El-Shazly
a,e,
Chieh-Fu
Chen
f,
Ping-Jyun
Sung
g,h,
Yan-Yu
Lin
a,
Yi-Hsin
Lin
a,
Yang-Chang
Wu
a,i,j,k,∗∗,
Mei-Chin
Lu
a,g,h,∗aGraduateInstituteofNaturalProducts,CollegeofPharmacy,KaohsiungMedicalUniversity,Kaohsiung807,Taiwan bCancerCenter,KaohsiungMedicalUniversityHospital,Kaohsiung807,Taiwan
cResearchandDevelopmentCenterofChineseHerbalMedicinesandNewDrugs,CollegeofPharmacy,KaohsiungMedicalUniversity,Kaohsiung807,Taiwan dDepartmentofMarineBiotechnologyandResources,NationalSunYat-senUniversity,Kaohsiung804,Taiwan
eDepartmentofPharmacognosyandNaturalProductsChemistry,FacultyofPharmacy,Ain-ShamsUniversity,OrganizationofAfricanUnityStreet11566,Abassia,Cairo,Egypt fChih-YingPlantResearchandDevelopmentFoundation,TaipeiCity100,Taiwan
gGraduateInstituteofMarineBiotechnology,NationalDongHwaUniversity,Pingtung944,Taiwan hNationalMuseumofMarineBiology&Aquarium,Pingtung944,Taiwan
iSchoolofChineseMedicine,CollegeofChineseMedicine,ChinaMedicalUniversity,Taichung404,Taiwan jNaturalMedicinalProductsResearchCenter,ChinaMedicalUniversityHospital,Taichung404,Taiwan kCenterforMolecularMedicine,ChinaMedicalUniversityHospital,Taichung404,Taiwan
a
r
t
i
c
l
e
i
n
f
o
Keywords: Antrodiacamphorata Dehydroeburicoicacid(DeEA) DNAdamage
Apoptosis
TopoisomeraseIIinhibitor
a
b
s
t
r
a
c
t
Antrodiacamphorata(AC)isanativeTaiwanesemushroomwhichisusedinAsianfolkmedicineasa chemopreventiveagent.Thetriterpenoid-richfraction(FEA)wasobtainedfromtheethanolicextractof ACandcharacterizedbyhighperformanceliquidchromatography(HPLC).FEAcausedDNAdamagein leukemiaHL60cellswhichwascharacterizedbyphosphorylationofH2A.XandChk2.Italsoexhibited apoptoticeffectwhichwascorrelatedtotheenhancementofPARPcleavageandtotheactivationof caspase3.Fivemajortriterpenoids,antcinK(1),antcinC(2),zhankuicacidC(3),zhankuicacidA(4), anddehydroeburicoicacid(5)wereisolatedfromFEA.ThecytotoxicityofFEAmajorcomponents(1–5) wasinvestigatedshowingthatdehydroeburicoicacid(DeEA)wasthemostpotentcytotoxiccomponent. DeEAactivatedDNAdamageandapoptosisbiomarkerssimilartoFEAandalsoinhibitedtopoisomerase II.InHL60cellsxenograftanimalmodel,DeEAtreatmentresultedinamarkeddecreaseoftumorweight andsizewithoutanysignificantdecreaseinmicebodyweights.Takentogether,ourresultsprovided thefirstevidencethatpureACcomponentinhibitedtumorgrowthinvivomodelbackingthetraditional anticanceruseofACinAsiancountries.
© 2012 Elsevier GmbH. All rights reserved.
Introduction
Antrodiacamphorata(AC),alsoknownasAntrodiacinnamomea orTaiwanofungus camphoratus,is anendemic fungusinTaiwan which only parasitizes in the internal heartwood or the dark humidwoodsurfaceofCinnamomumkanehirai(Bullcamphortree)
∗ Correspondingauthorat:GraduateInstituteofMarineBiotechnology,National DongHwaUniversity,Pingtung944,Taiwan.Tel.:+88688825038;
fax:+88688825087.
∗∗ Correspondingauthorat: School of ChineseMedicine, College ofChinese Medicine,ChinaMedicalUniversity,Taichung404,Taiwan.Tel.:+886422057153; fax:+886422060248.
E-mailaddresses:[email protected](Y.-C.Wu), [email protected](M.-C.Lu).
1Thefirsttwoauthorscontributedequallytothisstudy.
(Aoet al.2009;Luet al.2009a).MatureACfruitingbodies are
regarded asthecoloredgemsof theforests, rubyin Taiwanese forest,due totheircolorful orangetobrown-redappearance.It takesoneyearforACfruitingbodiestogrowtothesizeofoneEuro forcingthepriceofonekilogramtoreach10,000–28,000D.Due totheslowgrowthrateandthehighpriceofwildfruitingbodies, ACiswidelycultured.
AC,whichisknowninfolkmedicineas“chang-chih”hasalong historyinthepreventionandtreatmentofseverallifethreatening diseasesincludingcancer,inflammatory disorders,andhepatitis
(GeethangiliandTzeng2009;Huangetal.2010;Luetal.2009b;
Shenetal.2003;Yehetal.2009).Currently,ACisusedasfood
dietarysupplementforcancerpreventionandhepatoprotectionin severalAsianand Europeancountries.Itsproductsare commer-ciallyavailableinChina,Japan,Korea,Malaysia,andSingapore.It isalsomarketedintheUnitedKingdomunderthenameAntrodia camphorata.
0944-7113/$–seefrontmatter © 2012 Elsevier GmbH. All rights reserved. doi:10.1016/j.phymed.2012.03.014
ARTICLE IN PRESS
GModelPHYMED-51211; No.ofPages9
2 Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx
OurgrouphasbeendevotedtotheanticancerresearchofAC for many years. In ourprevious report, we demonstrated that the water extract of AC wild fruiting bodies (ACW) promoted thefunctional maturationofdendriticcellsintheexpressionof phenotypiccharacteristics,IL-12productionandchemotaxic
activ-ity (Lu et al.2009b). Moreover,according to1H NMR analysis,
it wassuggested thatthe polysaccharidesof ACWcouldact as apotentadjuvantforcancerimmunotherapythroughpromoting Th1immuneresponse.Wealsoreportedthattheethanolicextract fromthewildfruitingbodiesofAC(EEAC)couldinduceHL60cell apoptosisandpotentiatetheanticancereffectoftrichostatinA(Lu
etal.2009a).FractionationofEEACusingdifferentsolvents
pro-videdthreefractions,amongthemtheethylacetatefraction(FEA) wastheprimarycontributortoEEACcytotoxicity.The aforemen-tionedresultsencouragedustoidentifyFEAmajorcomponentsand studytheircytotoxicactivity.Webelievethatathorough under-standingof thecytotoxicmolecularmechanism inducedbyFEA majorcomponentswillshedlightonthepotentialutilizationof thesecomponentsasfutureanticanceragents.
Materialsandmethods
Assessmentofapoptosis
TheaccumulationofthesubG1populationincancercellswas
determinedbyflowcytometry.Cellswereseededonto6cmdishes and treatedwithor withouttheindicated concentrationofthe testedsamplesfor24h.CellswerethenwashedtwicewithPBSand collectedbycentrifugationat200×gfor5minat4◦C.Cellswere fixedin70%(v/v)ethanolat4◦C for30min.Afterfixation,cells weretreatedwith0.2mlofDNAextractionbuffer(0.2MNa2HPO4
and0.1Mcitricacidbuffer,pH7.8)for30min,centrifuged,and resuspended in 1ml of propidium iodide staining buffer(0.1% TritonX-100,100g/mlRNaseA,500g/mlofpropidiumiodide inPBS)at37◦Cfor30min.Cytometricanalyseswereperformed usingaflowcytometerFACS-Calibur(Becton-Dickinson,SanJose, CA,USA)andCellQuestsoftware.Approximately10,000cellswere countedforeachdetermination.
Theexternalizationofphosphatidylserine(PS)andmembrane integrity wasquantifiedusing anAnnexinV-FLOUS stainingkit (RocheDiagnosticsGmbH,Mannheim,Germany).Inshort,106cells
weregrownin35mmdiameterplatesandlabeledwithAnnexin V-FLOUS(10g/ml)andpropidiumiodide(PI)(Sigma–AldrichCorp., St.Louis,MO,USA)(20g/ml)priortoharvesting.Afterlabeling, allplateswerewashedwithbindingbufferandharvestedby scrap-ing.Cellswereresuspendedinbindingbufferataconcentrationof 2×105cells/mlbeforeanalysisbyflowcytometry.
NeutralcometassayforthedetectionofDNAdouble-strand breaks(DSBs)
TheassaywascarriedoutusingaCometAssayTMKit(Trevigen
Inc.,Gaithersburg,MD,USA)followingthemanufacturer’sprotocol fortheneutralcometassay.Briefly,cancercells(2×105cells/ml)
weretreatedwiththetestedsamplesfor18h.Cellswere com-binedwith1%lowmeltingpointagaroseataratioof1:10(v/v). From themixture,75l werepipetted onto CometSlideTM and
allowedtostandat4◦Cin thedark.The slideswereimmersed inice-coldlysissolutionfor30–60min.Theslideswereplacedin ahorizontalelectrophoresisapparatusandelectrophoresedin1× TBE(90mMTris–HCl,90mMboricacid,and2mMEDTA,pH8.0) at20V for10min.Thesampleswerethen fixedin70% ethanol anddriedbeforebeingstainedwith1:10,000SYBRGreenIto visu-alizecellularDNA.Thefluorescenceimageswereanalyzedusing theTriTekCometImageprogramtocircumscribethe‘head’and the‘tail’regions ofeach cometand theintegratedfluorescence
valuesofeachdefinedareawererecorded.Thecometlengthwas measuredfromthetrailingedgeofthenucleustotheleadingedge ofthetail.ThislengthwasindicativetotheextentofDNAdamage. Calculationswereaveragedperreplicate.
Westernblottinganalysis
Celllysateswerepreparedasdescribedpreviously(Yangetal. 2006), by treating thecells for 30minin RIPA lysis buffer[1× PBS, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 1mM sodium orthovanadate, 100g/ml phenylmethylsulfonylfluorideand30g/mlaprotinin,all chem-icalswereobtainedfromSigma–AldrichCorp.].Thelysateswere centrifugedat20,000× gfor30min,and theprotein concentra-tioninthesupernatantwasdeterminedusingaBCAproteinassay kit (Pierce, Rockford, IL, USA).Equal amounts of proteinswere respectivelyseparatedby7.5%,10%or12%ofSDS-polyacrylamide gelelectrophoresisandthenelectro-transferredtoaPVDF mem-brane.Themembranewasblockedwithasolutioncontaining5% fatfree drymilk TBSTbuffer(20mM Tris–HCl,pH7.4,150mM NaCland0.1%Tween20)for1handwashedwithTBSTbuffer. Pro-teinexpressionwasmonitoredbyimmunoblottingusingspecific antibodies:Bcl-2andBcl-x(Santa-CruzBiotechnology,SantaCruz, CA,USA);caspases3,7,8and9;Bax,Bid,Bad,p-H2AX,andPARP (CellSignalingTechnologies,Beverly,MA,USA).Caspases3,8,and 9inhibitors(Z-DEVD-FMK,Z-IETD-FMK,andZ-LEHD-KMK)were purchasedfromR&DSystemsInc.(Minneapolis,MN,USA). Anti-mouseandrabbit1gGperoxidase-conjugatedsecondaryantibodies werepurchasedfromPierce.Theseproteinsweredetectedbyan enhancedchemiluminescencekit(Pierce,Rockford,IL,USA). AssayoftopoisomeraseIIinhibitorsandpoisons
Theassaywasperformedasdescribedpreviouslywithminor modifications(Girietal.2010).Standardrelaxationreaction mix-tures(20l)containing50mMTris–HCl(pH8.0),10mMMgCl2,
200mM potassium glutamate, 10mM dithiothreitol, 50g/ml bovineserumalbumin,1mMATP,0.3gofpHOT1plasmidDNA, two unitsof human topoisomerase II(Topogen,Columbus, OH, USA)andtheindicatedconcentrationsofetoposideandDeEAwere incubatedat37◦Cfor30min.Reactionswereterminatedbyadding 2lof10%SDStofacilitatetrappingtheenzymeinacleavage com-plex,followedbytheadditionof2.5lofproteinaseK(50g/ml) todigesttheboundprotein(incubated37◦Cfor15min)andfinally byadding 0.1volofthesampleloadingdye.TheDNAproducts wereanalyzedbyelectrophoresisthroughvertical2%agarosegels at2V/cmin0.5×TAEbuffer.Gelswerestainedwithethidium bro-mideandphotographedusinganEagleEyeIIsystem(Stratagene, LaJolla,CA,USA).
HumanleukemiaHL-60cellsxenograftanimalmodel
Establishmentof nudemice withxenograftswasperformed asdescribedpreviously(Dowdyetal.2006).Six-week-oldfemale immunodeficientathymicmicewerepurchasedfromtheNational LaboratoryAnimaland ResearchCenter(Taipei,Taiwan).Allthe animalswere maintained under standard laboratoryconditions (temperature24–26◦C,12–12hdark–lightcircle)andfedwith lab-oratorydietandwater.Theprotocolsofanimalexperimentswere conformedtotheGuidefortheCareandUseofLaboratoryAnimals, publishedbytheUSNationalInstituteofHealth(NIHpublication no.85-23,revisedin1996).
HL60cells(1×107)resuspendedin0.2mlPBSwereinjecteds.c.
intotherightflankofeachmouse,andtumorgrowthwas moni-toredeveryday.Fourteendaysaftertumorcellinjection,micewith confirmedtumorgrowthwererandomlydividedintotwogroups.
Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx 3
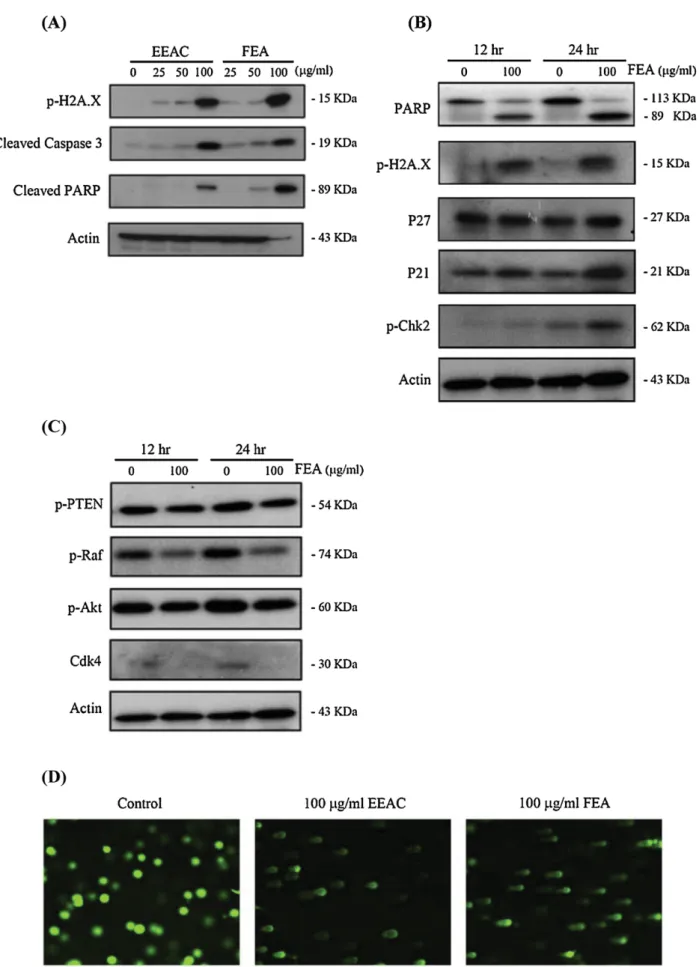
Fig.1.Westernplottingandalkalinecometassays.(A)EffectofEEACandFEAonapoptoticandDNAdamaged-relatedproteinsinHL60cells(24h).(B)EffectofFEAon tumorsuppressorgenes,apoptoticandDNAdamaged-relatedproteinsinHL60cells(12hand24h).(C)EffectofFEAontheexpressionofsurvival-relatedproteinsandCdk4 (12hand24h).(D)AssessmentofDNAdamagebycometassay.HL60cellsweretreatedwithEEACorFEAfor18handthensubjectedtothealkalinecometassaytodetect thebrokenDNA(comettails).
ARTICLE IN PRESS
GModelPHYMED-51211; No.ofPages9
4 Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx
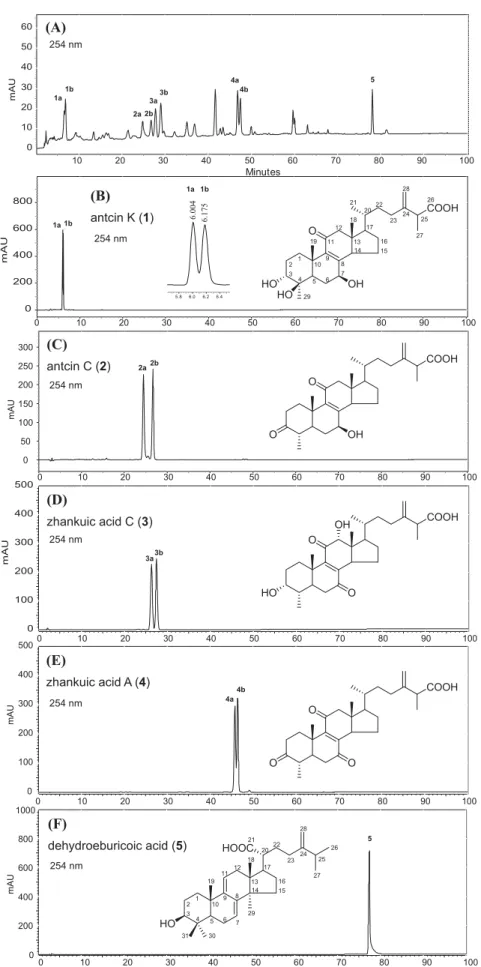
Fig.2. HPLCchromatogramoftheethylacetatefraction(FEA,anactivetriterpenoid-richfraction)fromtheethanolicextractofwildAC.Standardisolates(compounds1–5) areshownforcomparison.
Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx 5
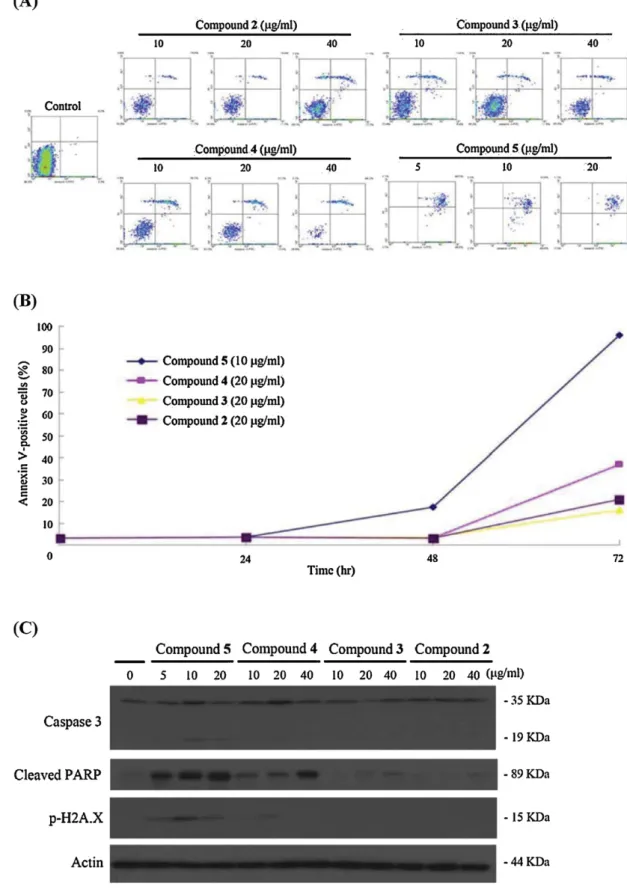
Fig.3.TheeffectofFEAcompounds2–5onapoptosisinductionandDNAdamageinHL60cells.(A)FACSdotplotsrepresentingHL60cellsstainedwithAnnexinVandPI aftertreatmentwith2–5(72h).(B)Effectof2–5onthepercentageofAnnexinV-positivecellsatdifferenttreatmenttimes(24,48,and72h).(C)Westernblottingdetecting theexpressionofapoptoticandDNAdamaged-relatedproteinscausedby2–5.
DeEA(10g/g)wasorallyadministeredtothetreatmentgroup and thecontrolgroup receivedsolventonly.DeEAwas admin-istratedeveryotherdayforfiveweeks.Animalsweresacrificed bycarbondioxide.Thetumorswerecarefullydissectedfromeach
mouse,andtumorweightsweremeasured.Halfofthetumortissue wasfixedwithformalinandusedforimmunohistochemical stud-ies,whereastheotherhalfwassnapfrozeninliquidnitrogenand storedat−80◦C.Thesetissueswereusedtoextractproteinsfor
ARTICLE IN PRESS
GModelPHYMED-51211; No.ofPages9
6 Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx
Fig.4.ThecytotoxiceffectofDeEAonHL60cells.(A)DeEAandFEAcytotoxicactivityatvariousconcentrations(72h)inMTTassay.(B)EffectofDeEAandFEAon anti-apoptoticandpro-apoptoticproteinsandcaspases3,7,and9(72h).(C)GraphicalrepresentationofthepercentagesofcellsatSubG1,G1,SandG2/MphasesinDeEA-treated
cells.(D)RepresentativeimagesofcometassayshowingabnormalcomettailscausedbychromosomalDNAdoublestrandbreaksinDeEA(2.5,5and10g/ml)treatedHL 60cells.(E)DeEAeffectontailmovementinHL60cells.HistogramwasgeneratedbyTriTekCometImageprogram.Valuesareexpressedasmean±SEM.*Significantly differentfromcontrolgroupsatp<0.05.(F)EffectofDeEAonthecleavageofPARPandonthephosphorylationofH2A.XandChk2.
westernblottingassay.Tumorsizewasmeasuredthreetimesa weekusingcalipersandtumorvolumeswerecalculatedaccording tothestandardformula:width2×length/2.
Statistics
Theresultswereexpressedasmean±standarddeviation(SD). Comparisonineachexperimentwasperformedusinganunpaired Student’st-test,anda pvalueofless than0.05wasconsidered statisticallysignificant.
Resultsanddiscussion
TheapoptoticandtheDNAdamagingactivitiesofEEACandFEA Ourpreviousstudyrevealedthattheethanolicextractofwild AC(EEAC)anditsactivefraction(FEA,theethylacetatefraction fromEEAC)possessedcytotoxicityagainst leukemiaHL60 cells withIC50valuesmeasuredafter24hof104.82and80.53g/ml,
respectively(Luetal.2009a).Inthecurrentstudy,EEACandFEA werepreparedaspreviously reported(seesupplementarydata,
Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx 7 Table1
ComparisonsofpeakareaandheightfortherespectivecomponentsinFEAwithHPLCat254nm.
Compound Peaknumber Retentiontime(min) Area(%) Height(%)
AntcinK 1a 6.004 4.96 7.08 1b 6.175 6.23 9.40 AntcinC 2a 24.805 7.34 4.20 2b 26.733 5.36 4.36 ZhankuicacidC 3a 27.778 6.91 6.59 3b 28.981 8.07 7.59 ZhankuicacidA 4a 46.825 7.17 9.24 4b 47.453 6.37 7.68 Dehydroeburicoicacid 5 78.058 4.58 8.79 Others 43.01 35.07 Total 100.00 100.00
and100g/mlofEEACorFEAandsubsequentlycelllysateswere analyzedbywesternblotting.FEAorEEACinduced phosphoryla-tionofH2A.XatSer-139(␥H2A.X)(Fig.1A)whichisconsideredas abiomarkerforDNAdouble-strandbreaks(KuoandYang2008; Mahetal.2010).Additionally,theuseofFEAorEEACenhancedthe cleavageofPARPandtheactivationofcaspase3,whichareboth regardedasmarkersofapoptosis(Fig.1A).Theseresultsimplied thatFEAistheactiveapoptoticfractionofEEACandthusselected forfurtherinvestigation.
TheeffectofusingFEA(100g/ml)onproteinexpressionwas also determined. FEA ledto a marked increase in the amount oftumorsuppressorgenes,p21andp27,enhancementofH2A.X and Chk2 phosphorylation, aswellas induction ofPARP cleav-age(Fig.1B).Ontheotherhand,phosphorylationlevelsineachof thesurvivalproteins(Akt,Raf,andPTEN)aswellasthe expres-sionofCdk4 decreasedfollowing treatmentwithFEAina time dependentmanner(Fig.1C).TheDNAdamagingeffectofFEAwas furtherconfirmedusingalkalinecometassayresultingincomets withabnormal tailssizes (Fig. 1D). Theaforementioned results confirmedtheDNAdamagingandtheapoptoticactivitiesofFEA implyingitsimportanceasapotentialcytotoxicagent(Kimetal.
2011).
TheapoptoticandtheDNAdamagingactivitiesofFEAmajor components
FEAmajorcomponentswereseparatedbynormalphase chro-matography and purified by reverse phase high performance liquidchromatography(HPLC)yieldingfivemajorconstituents(see
supplementarydata, sectionS4). Theisolated compoundswere
fourergostanes,antcinK(1),antcinC(2),zhankuicacidC(3),and zhankuicacidA(4)aswellasonelanostane,dehydroeburicoicacid
(5)(Chenetal.1995;Shenetal.2003).Theisolationofthesemajor
triterpenesasstandardsledtotheestablishmentofHPLC finger-print ofFEA(Fig.2)(seesupplementarydatafor detailedHPLC
condition,sectionS5).Accordingtothepublishedreport(Chenetal.
1995),eachACergostanetriterpene(compounds1–4)existsasa pairofstereoisomersduetothepresenceofchiralcenteratC-25. Twopeaksrepresentinganisomericpairofeachergostanecanbe observedintheHPLCfingerprint.Forexample,peakpairs1aand 1b,2aand2b,3aand3b,aswellas4aand4bcorrespondingto antcinK(1),antcinC(2),zhankuicacidC(3),andzhankuicacid A(4),respectivelywereobserved(Fig.2B–E).Ontheotherhand, dehydroeburicoicacidwithnochiralcenteratC-25existedonly asasingleisomer,peak5(Fig.2F).PeakareasandheightsofFEA majorcomponents(1–5)at254nmwerecalculatedandcompared asshowninTable1.Theareasofthelabeledpeaks(1–5)accounted for56.99%ofthetotalpeakareasobservedintheHPLCfingerprint.
Regardingtheantiproliferativeactivityof1–5againstleukemia HL 60 cells, 1showednoactivity evenat a highconcentration (100g/ml). Compounds2–5exhibitedantiproliferative activity againstHL60cellsandwerefurtherinvestigatedusingAnnexin VandPIdoublestainingtechnique.HL60cellsweretreatedby dif-ferentconcentrationsofantcinC(2),zhankuicacidC(3),zhankuic acidA(4),anddehydroeburicoicacid(5)for24,48,and72h.Cells werestainedwithAnnexinV/PIandexaminedwithflow cytom-etry.FACSdotplotsrepresentingAnnexinVandPIstainingafter thetreatmentwith2–5areshowninFig.3A.Thequantification ofAnnexinV/PIexperimentrevealedthat4and5increasedthe apoptoticpopulationofHL60cellsinadosedependentmanner. Ontheotherhand,only5(10g/ml)inducedapoptosisinatime dependentmanner(Fig.3B).
Compounds 4 and 5 induced cleavage of PARP in a dose dependentmanner(Fig.3C).Thetreatmentof HL60 cells with
4 (20g/ml) and 5 (10g/ml) induced the activation of cas-pase 3 and the phosphorylation of H2A.X. The analytical and thebiologicalassaysclearlyimpliedthat 5(DeEA)isFEAmajor component withthemostpotentapoptotic andDNA damaging activities.
TheapoptoticandtheDNAdamagingactivitiesofDeEA
TofurtherconfirmthefactthatDeEAistheleadactive com-poundofFEA(triterpenoid-richfraction)thecytotoxicityofDeEA andFEAagainstHL60cellswascompared(Fig.4A).DeEA exhib-itedmorepotentcytotoxicitycomparedtoFEAwithIC503.39and
22.4g/ml,respectively,after72hasindicatedbyMTTassay.The effectofDeEAandFEAontheexpressionofanti-apoptotic (Bcl-2 and Bcl-x) and pro-apoptotic (Bax, Bid,and Bad) proteinsas wellastheactivationofcaspases3,7,and9wasalsoinvestigated (Fig.4B).DeEAwasmorepotentcomparedtoFEAinenhancing theexpressionofpro-apoptoticproteinsBidandBad.AlsoDeEA inducedtheactivationofcaspases3,7,and9andsimilarresults wereobserveduponthetreatmentofFEA.Toanalyzewhetherthe apoptoticinductionofDeEAandFEAinvolvesintrinsicorextrinsic apoptoticpathways,specificinhibitorsforcaspases3,8,and9were tested.Pretreatmentofcellswithcaspaseinhibitorsdidnotinhibit theapoptoticeffectofDeEAandFEA,suggestingthattheapoptotic effectofDeEAandFEAisindependentoncaspaseactivation(data notshown).
TreatmentofHL60cellswithincreasingdosesofDeEA(0,2.5,5 and10g/ml)resultedinG2/Mdistributionratesof21.1%,26.6%, 28.7%and39.5%,respectivelyafter24h(Fig.4C).Similarresults wereobservedafter48and72h.ThusDeEAtreatmentappearedto induceG2/Mphasearrestinadosedependentmanner.
ToclarifytheroleofDeEAasDNAdamagingagent,wetreatedHL 60cellswithvariousconcentrationsofDeEAfor18handanalyzed
ARTICLE IN PRESS
GModelPHYMED-51211; No.ofPages9
8 Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx
theleveloftailmovementwiththealkalinecometassay.DeEAat 10g/mlcausedsevereDNAdamageinHL60cellsasindicatedby abnormaltailsizeincometassay(Fig.4DandE).
ToexaminetheextentofDNAdamagewhichcanleadtothe acti-vationofcellcyclecheckpointsinHL60cells,theeffectofDeEAon PARP,p-H2A.Xandp-Chk2wasstudied(Fig.4F).DeEAat10g/ml significantlyincreasedthephosphorylationofChk2andH2A.X,but didnotchangethephosphorylationofChk1(datanotshown). Addi-tionally,DeEAinducedPARPcleavage.Thesefindingsconfirmed theapoptotic and theDNA damagingactivities of DeEAon HL 60cells.
TheinhibitoryactivityofDeEAontopoisomeraseII
TopoisomerasesareessentialenzymesforDNAmetabolismand theirinhibitorsareemergingaspotentialcytotoxicagents. Expo-sureof cellsto DNAtopoisomerase I(topoI) or topoisomerase II(topo II) inhibitorsleads toDNA damage that ofteninvolves
Fig.5.EffectofDeEAontopoisomeraseIImediatedsupercoiledpHOT1plasmid DNArelaxation.Lane1–3:positivecontrol,etoposide(100,200and400M);Lane 4:negativecontrolplasmidDNA;Lane5–9:DeEA(0,2.5,5,10,and20g/ml);Lane 10:plasmidDNA+topoisomeraseII;Lane11:linearDNA.
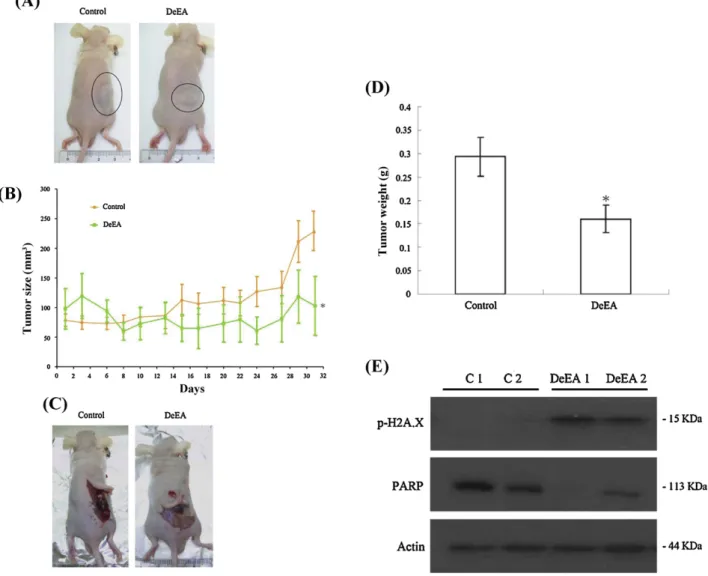
Fig.6.EffectofDeEAonHL60cellstumorgrowthinxenograftanimalmodel.FemalenudemicebearingleukemiaHL60tumorsweretreatedwiththesolvent(negative control)orDeEA(10g/g)for5weeks.(A)Arepresentativepictureoftumorgrowthinxenograftnudemiceadministratedsolventonly(left)andDeEA(right).(B)Tumor volumesweremeasuredeveryotherday,andresultsareexpressedasmean±SEM.*Significantlydifferentfromcontrolgroupsatp<0.05.(C)Arepresentativepictureof tumortissueasitappearsintherightflankofthemouse.(D)HistogramofthetumorweightfromthecontrolgroupandDeEAtreatedgroup.Valuesareexpressedas mean±SEM.*Significantlydifferentfromcontrolgroupsatp<0.05.(E)EffectofDeEAonPARPandonH2A.Xintumorsamples.Twomicewerepickedrandomlyfromeach group.C1andC2representtwomicefromthecontrolgroup.DeEA1andDeEA2representtwomicefromtheDeEAtreatedgroupchosenrandomly.
Y.-C.Duetal./Phytomedicinexxx (2012) xxx–xxx 9 theformationofDNAdouble-strandbreaks(Darzynkiewiczetal.
2009; Deng et al. 2009).Mizushina et al. (2004) revealed that
dehydroebriconicacid,fromthesclerotiumofPoriscocosinhibited DNAtopoIIwithnoactivityonDNAtopoI.Thestructureof dehy-droebriconicacidissimilartoDeEAexceptthehydroxylgroupat C-3inDeEAisreplacedbyacarbonylgroupindehydroebriconic acid.
TheinhibitoryactivityofdehydroebriconicacidonDNAtopo IIencouragedustoexaminewhetherDeEAtreatmentcaninduce DNAdamageinleukemiacellsthroughinhibitionoftopoIIornot. Forthis purpose,a cell-freeDNAcleavageassaywasperformed using enzyme-mediated negatively supercoiled pHOT1plasmid DNA.ItwasfoundthatDeEAinducedDNAcleavageinthepresence oftopoII(Lanes6–9)(Fig.5),asindicatedbytheappearanceof lin-earDNA(markedbyL).Theresultsweresimilartothoseobtained uponutilizingetoposide,astandardtopoIIinhibitor(Lanes1–3) (Dasetal.2007).
TheinhibitoryeffectofDeEAontumorgrowthinxenograftanimal model
HL60cellswereinoculatedsubcutaneouslyattherightflankof femaleimmunodeficientathymicmice.After5weeksoftreatment, thetumorgrowthofHL60cellswassignificantlysuppressedby oraladministrationofDeEA(10g/g)(Fig.6A).Theaveragetumor sizeonday31inthecontrolgroupwas229.46mm3,whereasthe
averagetumorsizeintheDeEA-treatedgroupwas102.75mm3
(Fig.6B).ThetumorsizewassignificantlylowerintheDeEA-treated groupas compared tothecontrol group(p<0.05) withno sig-nificantdifferenceinthemice bodyweights. Attheend of the treatment,thetumortissuewasisolatedandweighed(Fig.6C). The mean of tumor weights was obviously less in the DeEA-treatedgroupcomparedtothecontrolgroup(p<0.05)(Fig.6D). Thewesternblottinganalysisoftumortissueindicatedthat the levelofPARPwasdecreasedandthephosphorylationofH2A.Xwas increasedinDeEA-treatedgroupcomparedtothecontrolgroup (Fig.6E).
Conclusion
Inthisstudy,weillustratedtheapoptoticandtheDNA damag-ingactivitiesofthetriterpenoid-richfraction(FEA)anditsmajor componentsobtainedfromtheethanolicextractofA.camphorata. FEAinducedPARPcleavageandcaspase3activationindicatingits apoptoticactivity.ItalsoinducedthephosphorylationofH2A.X as a marker of DNA double strandbreak. From the active FEA fraction, dehydroeburicoicacid(DeEA)wasisolated asthelead most active component. DeEAinduced G2/M phase arrest in a dose dependent manner in HL 60 cells. It also induced apop-toticandDNAdamagingmarkersrevealingitsroleasthepotent cytotoxicconstituentofFEA.AdditionallyitinhibitedtopoII sug-gestingthepossibleroleofthisenzymeinthecytotoxicactivity ofDeEA.TheantitumoreffectofDeEAwasfurtherdemonstrated inreducingtumorweightandsizeinxenograftanimalmodel.The presentfindingsnotonlysupporttheethnopharmacologicaluse ofA.camphoratabutalsodivulgetheantitumoractivityof dehy-droeburicoicacidasthemajorleadcompoundofthistreasured mushroom.
Conflictofinterest
Theauthorsdeclarethattheyhavenoconflictsofinterest.
Acknowledgments
ThisworkwassupportedbytheNationalScienceCouncilgrants (NSC98-2324-B-037-001and98-2321-B-037-060)andthe Depart-mentofHealth,ExecutiveYuan,Taiwan(DOH99-TD-C-111-002). ThisworkwasalsosupportedbytheAcademic-Industrial Collab-orative ProjectofKaohsiung Medical Universityand YungPeng BiotechnologyCo.,Ltd.,Kaohsiung(S099009and100A502).
AppendixA. Supplementarydata
Supplementarydataassociatedwiththisarticlecanbefound,in theonlineversion,atdoi:10.1016/j.phymed.2012.03.014.
References
Ao,Z.H.,Xu,Z.H.,Lu,Z.M.,Xu,H.Y.,Zhang,X.M.,Dou,W.F.,2009.Niuchangchih (Antrodiacamphorata)anditspotentialintreatingliverdiseases.Journalof Ethnopharmacology121,194–212.
Chen,C.H.,Yang,S.W.,Shen,Y.C.,1995.NewsteroidacidsfromAntrodiacinnamomea, afungalparasiteofCinnamomummicranthum.JournalofNaturalProducts58, 1655–1661.
Darzynkiewicz,Z.,Halicka,D.H.,Tanaka,T.,2009.CytometricassessmentofDNA damageinducedbyDNAtopoisomeraseinhibitors.MethodsinMolecular Biol-ogy582,145–153.
Das,C.M.,Aguilera,D.,Vasquez,H.,Prasad,P.,Zhang,M.,Wolff,J.E.,Gopalakrishnan, V.,2007.Valproicacidinducesp21andtopoisomerase-II(alpha/beta) expres-sionandsynergisticallyenhancesetoposidecytotoxicityinhumanglioblastoma celllines.JournalofNeuro-Oncology85,159–170.
Deng,J.Y.,Chen,S.J.,Jow,G.M.,Hsueh,C.W.,Jeng,C.J.,2009.Dehydroeburicoicacid inducescalcium-andcalpain-dependentnecrosisinhumanU87MG glioblas-tomas.ChemicalResearchinToxicology22,1817–1826.
Dowdy,S.C.,Jiang,S.,Zhou,X.C.,Hou,X.,Jin,F.,Podratz,K.C.,Jiang,S.W.,2006.Histone deacetylaseinhibitorsandpaclitaxelcausesynergisticeffectsonapoptosisand microtubulestabilizationinpapillaryserousendometrialcancercells.Molecular CancerTherapeutics5,2767–2776.
Geethangili,M.,Tzeng,Y.M.,2009.ReviewofpharmacologicaleffectsofAntrodia camphorataanditsbioactivecompounds.EvidenceBasedComplementaryand AlternativeMedicine2011,1–17.
Giri,R.,Goodell,J.R.,Xing,C.,Benoit,A.,Kaur,H.,Hiasa,H.,Ferguson,D.M.,2010. Synthesisandcancercellcytotoxicityofsubstitutedxanthenes.Bioorganicand MedicinalChemistry18,1456–1463.
Huang,C.H.,Chang,Y.Y.,Liu,C.W.,Kang,W.Y.,Lin,Y.L.,Chang,H.C.,Chen,Y.C.,2010. FruitingbodyofNiuchangchih(Antrodiacamphorata)protectsliversagainst chronicalcoholconsumptiondamage.JournalofAgriculturalandFood Chem-istry58,3859–3866.
Kim,S.,Jun,D.H.,Kim,H.J.,Jeong,K.C.,Lee,C.H.,2011.Developmentofahigh-content screeningmethodforchemicalsmodulatingDNAdamageresponse.Journalof BiomolecularScreening16,259–265.
Kuo,L.J.,Yang,L.X.,2008.Gamma-H2AX—anovelbiomarkerforDNAdouble-strand breaks.InVivo22,305–309.
Lu,M.C.,Du,Y.C.,Chuu,J.J.,Hwang,S.L.,Hsieh,P.C.,Hung,C.S.,Chang,F.R.,Wu,Y.C., 2009a.ActiveextractsofwildfruitingbodiesofAntrodiacamphorata(EEAC) induceleukemiaHL60cellsapoptosispartiallythroughhistone hypoacetyla-tionandsynergisticallypromoteanticancereffectoftrichostatinA.Archivesof Toxicology83,121–129.
Lu,M.C.,Hwang,S.L.,Chang,F.R.,Chen,Y.H.,Chang,T.T.,Hung,C.S.,Wang,C.L., Chu,Y.H.,Pan,S.H.,Wu,Y.C.,2009b.ImmunostimulatoryeffectofAntrodia cam-phorataextractonfunctionalmaturationofdendriticcells.FoodChemistry113, 1049–1057.
Mah,L.J.,El-Osta,A.,Karagiannis,T.C.,2010.␥H2AX:asensitivemolecularmarker ofDNAdamageandrepair.Leukemia24,679–686.
Mizushina,Y.,Akihisa,T.,Ukiya,M.,Murakami,C.,Kuriyama,I.,Xu,X.,Yoshida,H., Sakaguchi,K.,2004.AnovelDNAtopoisomeraseinhibitor:dehydroebriconic acid,oneofthelanostane-typetriterpeneacidsfromPoriacocos.CancerScience 95,354–360.
Shen,C.C.,Kuo,Y.C.,Huang,R.L.,Lin,L.C.,Don,M.J.,Chang,T.T.,Chou,C.J.,2003.New ergostaneandlanostanefromAntrodiacamphorata.JournalofChineseMedicine 14,247–258.
Yang,H.L.,Hseu,Y.C.,Chen,J.Y.,Yech,Y.J.,Lu,F.J.,Wang,H.H.,Lin,P.S.,Wang,B.C., 2006.Antrodiacamphoratainsubmergedcultureprotectslowdensity lipopro-teinsagainstoxidativemodification.AmericanJournalofChineseMedicine34, 217–231.
Yeh,C.T.,Rao,Y.K.,Yao,C.J.,Yeh,C.F.,Li,C.H.,Chuang,S.E.,Luong,J.H.,Lai,G.M.,Tzeng, Y.M.,2009.CytotoxictriterpenesfromAntrodiacamphorataandtheirmodeof actioninHT-29humancoloncancercells.CancerLetters285,73–79.