Anti-tumor and anti-inflammatory properties of ankaflavin and monaphilone A from Monascus purpureus NTU 568
Li-Chuan Hsu, †§Ya-Wen Hsu, †§Yu-Han Liang, †§ Yao-Haur Kuo*, §‡ and Tzu-Ming
Pan*,†
Department of Biochemical Science & Technology, College of Life Science, National
Taiwan University, Taipei, 106, Taiwan, R.O.C.
National Research Institute of Chinese Medicine, Taipei, 102, Taiwan, R. O. C.
Graduate Institute of Integrated Medicine, China Medical University, Taichung, 404,
Taiwan, R. O. C.
National Research Institute of Chinese Medicine, Taipei, 102, Taiwan, R. O. C.
* Corresponding Author: Tzu-Ming Pan, Professor; Tel: +886-2-3366-4519 ext 10;
fax: +886-2-3366-3838; e-mail: [email protected], Yao-Haur Kuo, Professor; Tel:
+886-2-2820-1999 ext. 7061; Fax: +886-2-2823-6150; E-mail: [email protected]; †
Institute of Microbiology and Biochemistry, National Taiwan University §
National Research Institute of Chinese Medicine ‡
ABSTRACT
An azaphilonidal derivative monaphilone A (MA), was recently isolated from the fermented products of Monascus purpureus NTU 568 by our laboratory. We report here the exploration of apoptosis-related and anti-inflammatory properties of MA and ankaflavin (AK) by some experiments about inducing death of human laryngeal carcinoma cell line HEp-2 and reducing inflammatory responses on murine macrophage RAW 264.7 cells. We employed ssDNA ELISA kit to investigate the nuclear changes of early apoptosis induced by AK and MA on HEp-2 cells, and used Western blot and enzyme activity assay to demonstrate the activation of caspase-3, -8 and -9 by MA and AK. Our studies revealed that AK and MA may decrease lipopolysaccharide (LPS) -induced inflammatory responses, including nitrite productions and expressions of inducible nitric oxide synthase (iNOS) and cyclooxygenase 2 (COX-2) in RAW 264.7 cells. All evidences support that azaphilonidal derivatives from Monascus purpureus NTU 568, such as AK and MA, are suitable for the development of chemotherapy or chemopreventive agents.
INTRODUCTION
Monascus species has traditionally been used to make food more colorful and easy
to preserve in Asian countries for thousands of years. Using rice as fermented
substrate, its product, red mold rice (RMR), has been proved to possess various
biological functions. For example, RMR exhibited hypolipidemic effects in a
hyperlipidemia hamster model (1), antifatigue activities to Wistar rats through
swimming exercise (2), neuroprotective properties against Alzheimer’s risk factors in Aβ40-infused rats (3), and preventive ability for obesity in a high-fat diet rat model (4), etc.
Some bioactive components from Monascus species have been identified and
characterized. A family of monacolins possessed HMG-CoA reductase inhibitory
activity (5), and monacolin K was the marker monacolin (6). Other bioactive secondary metabolites, including γ-amino butyric acid (GABA) and dimerumic acid, were considered to possess hypotensive (7, 8) and antioxidant activities respectively
(9). Furthermore, yellow pigments in Monascus sp. were demonstrated to possess
cytotoxic and anti-inflammatory effects (10, 11). Additionally, several
Monascus-pigment derivatives produced by fermentation with various amino acids
(12) showed different kinds of usages, including anti-microbial activity (13),
Carcinogenesis was involving in complicated steps, possibly resulted from chronic
inflammatory stimuli in the beginning, and leading to uncontrolled growth of tumor
cells in the end. It was hoped that some nature products could exert anti-tumor effect
by apoptosis-related mechanism or reduction of inflammatory responses. Our group
has found that fermented products from Monascus purpureus NTU 568 showed
anti-tumor and anti-inflammatory effects. For example, RMR extracts from Monascus
purpureus NTU 568 significantly reduced tumor progression of Lewis lung carcinoma
bearing mice (16) and also mitigated oral carcinogenesis through anti-inflammatory
responses in a hamster model (17). Recently, three new azaphilone pigments were
isolated from Monascus purpureus NTU 568 and reported to be cytotoxic to cancer
cell lines (18), and monaphilone A (MA) was the most cytotoxic compound. Here, we
report the exploration of apoptosis-related and anti-inflammatory mechanisms of MA
and ankaflavin (AK). To the purpose, we designed some experiments including of
induction of apoptosis in human laryngeal carcinoma cell line HEp-2 and reduction of
inflammatory responses on murine macrophage RAW 264.7 cells.
MATERIALS AND METHODS
General Experimental Procedures. NMR spectra were run on a Brucker NMR
NMR spectrometers (Varian Gemini 200 MHz, Varian Inc., Palo Alto, CA, USA)
using d6-acetone as solvent. HPLC separations were performed on a Shimadzu
LC-6AD series apparatus with a SPD-6AV UV detector, equipped with a 250 x 20
mm i.d. preparative Cosmosil AR-II column (Nacalai Tesque, Inc., Kyoto, Japan).
Reagents. Methanol and acetonitrile (HPLC grade), acetone, ethyl acetate,
n-hexane and methanol (analytical grade) were purchased from ECHO (Miaoli,
Taiwan). Trifluroacetic acid (TFA), anisaldehyde and sulphyric acid were purchased
from Merck. Fetal bovine serum (FBS), minimum essential medium (MEM),
Dulbecco’s minimum essential medium (DMEM), phosphate buffered saline (PBS)
and trypan blue were purchased from Biological Industries (Kibbutz Beit-Haemek,
North District, Isreal). Other chemicals, such as lipopolysaccharides (LPS, from
Escherichia coli O55:B5), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium
bromide (MTT) and dimethyl sulfoxide (DMSO) were obtained from Sigma (St.
Louis, MO, USA).
Extraction and Purification. Preparations of AK and MA were followed the
experimental procedures from our previous publication (18). Briefly described, the
dried RMR was extracted three times with acetone. After filtration and concentration,
the residue was chromatographyed on silica gel column and Sephadex (LH-20) gel
high-performance liquid chromatography (HPLC) to obtain AK and MA
Cell Lines and Culture. Human laryngeal carcinoma cell line HEp-2 and murine
macrophage cell line RAW 264.7 were obtained from Bioresources Collection and
Research Center (Hsinchu, Taiwan). HEp-2 cells were maintained in MEM (5% FBS)
in a humified incubator with 5% CO2 at 37oC, and RAW 264.7 cells were maintained
in DMEM (5% FBS).
Cytotoxicity Assay. Cytotoxicity assay was modified from the reported methods
(19, 20). HEp-2 cells (3 x 103 per well) were seeded with 180 μL of MEM in 96-well plates. After 4 hr, 20 μL of test agents dissolved in PBS solution were added at final concentrations of 5, 10, 25 and 50 μg/mL and incubated in a 37o
C incubator with 5%
CO2. Culturing for 24, 48 and 72 hr, 20 μL of MTT solution (2 mg/mL) was added to
each well and incubated for 4 hr to make cellular conversion of a tetrazolium salt into a formazan product. Then the supernatant was removed and 200 μL of DMSO was added to dissolve the formazan. Finally, the formazan can be detected by
spectrophotometry in the absorbance at 570 nm and provided a relative estimate of
cell viability. The same test agents were treated on RAW 264.7 cells with similar
procedures. However, RAW 264.7 cells were seeded for 2 x 105 per well and maintained with 500 μL of DMEM in 24-well plates. Test agents were added at final concentrations of 5 and 10 μg/mL.
Nitrite Production Assay. RAW 264.7 cells (2 x 105 per well) were seeded and maintained with 500 μL of DMEM in 24-well plates. After 12 hr, cells were treated with LPS (1 μg/mL) and test agents (10 μg/mL) dissolved in DMEM. After 24 hr of incubation, determination of the nitrite levels in supernatants was performed by Griess
reagent kit (Promega, Madison, WI, USA), and adapted from the reported methods
(21).
Early Detection of Apoptosis. This assay was designed for detection of early
apoptosis by using ssDNA apoptosis ELISA kit (Millipore, Billerica, MA, USA), and
adapted from the reported methods (22). HEp-2 cells (about 10000 cells per well)
were transferred into a 96-well microplate and treated with camptothecin (CPT, 0.2 μg/mL), AK (50 μg/mL) and MA (50 μg/mL) in 200 μL of MEM (5% FBS) for 12 hr. As positive control and negative control, 100 μL of ssDNA (0.3 μg/mL) and S1 nuclease were added separately. For all conditions, antibody mixture (recognized for
ssDNA) and 2,2'-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid) (ABTS solution)
were added. Finally, we easily detect early-apoptotic cells by measuring the
absorbance at 405 nm in a standard microplated system.
Western Blot Analysis. Cells (about 5 x 105 for HEp-2 and 8 x 106 for RAW 264.7) were seeded with 10 mL of media in a 75 cm2 flask. After 12 hr, 10 mL of test agents
harvested and extracted by RIPA lysis buffer (Millipore, Bellerica, MA, USA) with
1% protease inhibitor (Sigma, St. Louis, MO, USA). The cell lysates were analyzed
with primary antibodies, including of caspase-3 antybody (Novus biologicals,
Littleton, CO, USA), β-actin antibody (Epitomics, Burlingame, CA, USA), inducible nitric oxide synthase (iNOS) and cyclooxygenase 2 (COX-2) polyclonal antibody
(Cayman Chemical, Ann Arbor, MI, USA). The anti-rabbit or anti-mouse secondary
horseradish peroxidase antibodies (Jackson ImmunoResearch, West Grove, PA, USA)
were further added. Finally, the detection was performed using the Western lightning
chemiluminescence reagent (PerkinElmer Life Sciences, Waltham, MA, USA).
Caspase Activity Assay. Cells (about 5 x 105 for HEp-2 and 8 x 106 for RAW 264.7) were seeded with 10 mL of media in a 75 cm2 flask. After 12 hr, 10 mL of test agents
dissolved in media were replaced. After 12 and 24 hr of incubation, the cells were
harvested and tested for caspase-8 and caspase-9 activities respectively by using
colorimetric assay kit (BioVision, Linda Vista Avenue, CA, USA). Caspase activity
was determined according to the manufacturer’s protocol.
Statistical Analysis. Data were presented as mean ± standard deviation (n = 3). The
statistical comparisons were performed by one-way analysis of variance (ANOVA)
RESULTS
Identification of MA and AK from RMR. We purified these two compounds by
preparative HPLC twice in the final step, and identified them according to the
following MS and NMR data. Monaphilone A: ESIMS m/z 383 [M + Na]+. 1H NMR
(d6-acetone, 400 MHz) : 0.87 (3H, t, J = 6.8, H-21), 1.15 (3H, s, H-12), 1.27 (8H, m, H-17 -20), 1.54 (2H, m, H-16), 1.82 (3H, d, J = 7.2, H-11), 2.21 (1H, m, H-5a), 2.47 (1H, m, H-13a), 2.49 (3H, m, H-5b, H-15), 2.54 (1H, m, H-6), 2.91 (1H, d, J = 15.2, H-13b), 4.71 (1H, d, J = 12.4, H-1a), 4.88 (1H, d, J = 12.4, H-1b), 5.41 (1H, s, H-4), 6.01 (1H, d, J = 15.2, H-9), 6.39 (1H, dq, J = 15.2, 7.2, H-10); 13C NMR (d6-acetone, 100 MHz) δ: 14.2 (C-21), 18.3 (C-11), 20.0 (C-12), 23.2 (C-20), 24.4 (C-16), 29.9 (C-17, C-19), 32.4 (C-18), 32.9 (C-5), 40.3 (C-6), 42.4 (C-13), 43.4 (C-15), 64.3 (C-1), 74.8 (C-7), 104.3 (C-4), 114.2 (C-8a), 125.7 (C-9), 134.1 (C-10),
152.4 (C-4a), 160.3 (C-3), 198.6 (C-8), 209.8 (C-14). Ankaflavin: ESIMS m/z 387
[M + H]+. 1H NMR (d6-acetone, 400 MHz) δ: 0.87 (3H, t, J = 6.8, H-21), 1.29 (8H, m, H-17 ~ 20), 1.45 (3H, s, H-12), 1.60 (2H, m, H-16), 1.82 (3H, d, J = 7.2, H-11), 2.66 (2H, m, H-5), 2.70 (1H, m, H-15a), 2.92 (1H, m, H-15b), 3.15 (1H, m, H-6), 4.27 (1H, d, J = 13.2, H-13), 4.68 (1H, d, J = 12.4, H-1a), 4.90 (1H, d, J = 12.4, H-1b), 5.50 (1H, s, H-4), 6.01 (1H, d, J = 15.6, H-9), 6.41 (1H, dt, J = 15.6, 7.2, H-10); 13C NMR (d6-acetone, 100 MHz) δ: 14.2 (C-21), 17.7 (C-12), 18.3 (C-11), 23.2 (C-20),
23.6 (C-16), 29.2 (C-5), 29.9 (C-17, C-19), 32.3 (C-18), 43.4 (C-15), 44.4 (C-6), 55.3
(C-13), 64.2 (C-1), 84.0 (C-7), 104.6 (C-4), 115.1 (C-8a), 125.5 (C-9), 134.6 (C-10),
151.6 (C-4a), 160.2 (C-3), 171.3 (C-13a), 190.6 (C-8), 203.6 (C-14). Both of them
were yellow pigments and structurally similar to each other. They were azaphilones
containing the same alkyl group, but MA was decarboxylated and resulting a breakage
on the ester bond. The purity of MA and AK was confirmed by HPLC chromatogram
(Figure 1).
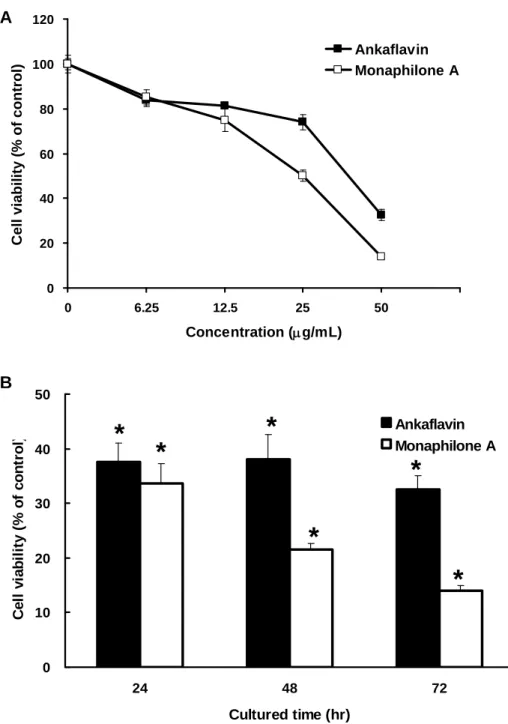
Cytotoxicity of MA and AK on HEp-2 Cells. To study the inhibitory effects on
cell viability of HEp-2 (human laryngeal carcinoma) cell lines by MA and AK, we
utilized MTT assay for a three-day course (Figure 2). Both of the two azaphilone
derivatives possessed dose-dependent and moderated cytotoxic activity against HEp-2
cells. The structure of MA was similar as AK, whereas it had better cytotoxic activity
(IC50 = 20.97 ± 2.27 μg/mL) against HEp-2 cells, than that of AK (IC50 = 31.62 ± 2.51 μg/mL). The increase of cytotoxicity against human laryngeal carcinoma cell line was possibly resulted from the structural changes between C-12 and C-14. The results also
suggested that MA and AK were suitable for apoptotic approach for the next step.
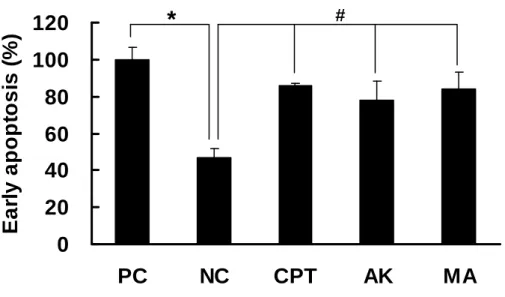
Induction of Early Apoptosis by MA and AK on HEp-2 Cells. Since MA
possessed moderate cytotoxicity against HEp-2 cells, we have examined whether MA induced apoptosis or necrosis. After HEp-2 cells treating with CPT (0.2 μg/mL,
reference compound for cytotoxity), AK (50 μg/mL) and MA (50 μg/mL) for 12 hr, the occurrences of early apoptosis were analyzed by ssDNA apoptosis ELISA kit
(Figure 3). Comparing with the negative control, both of the two azaphilones
exhibited significant increment of ssDNA formation, which was regarded as early
apoptosis. The results confirmed that it was apoptosis, not necrosis, induced by the
treatment of MA and AK.
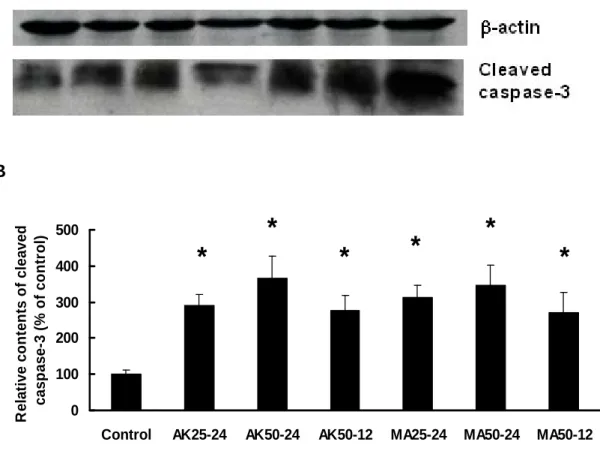
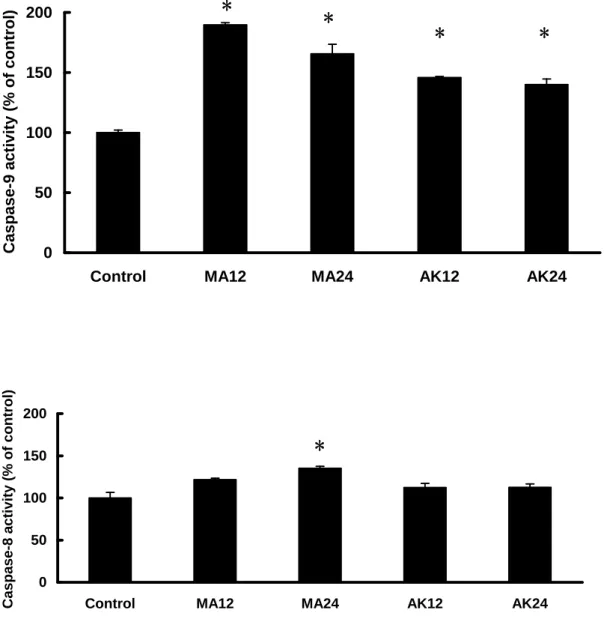
Caspase Activation of MA and AK on HEp-2 Cells. HEp-2 cells were treated
with 25 and 50 μg/mL of MA and AK for 12 and 24 hr, and further analyzed for the cleaved caspase-3 by Western blot (Figure 4) and enzyme activity of caspase-8 and -9 by colorimetric assay kit (Figure 5). Treatment of MA and AK (25 and 50 μg/mL) exhibited remarkable contents of cleaved caspase-3, which were regarded as caspase-3 activation resulted from apoptosis. As to caspase-9 activation, treatment of MA (50 μg/mL) and AK (50 μg/mL) exhibited significant increases of caspase-9 activity. However, caspase-8 activation was quite different between MA and AK. Comparing with all conditions, only treatment of MA (50 μg/mL, 24 hr) exhibited significant increase of caspase-8. Thus, MA was demonstrated to induce apoptosis through both of caspase-8 and -9 activations, but AK was merely through caspase-9 activation.
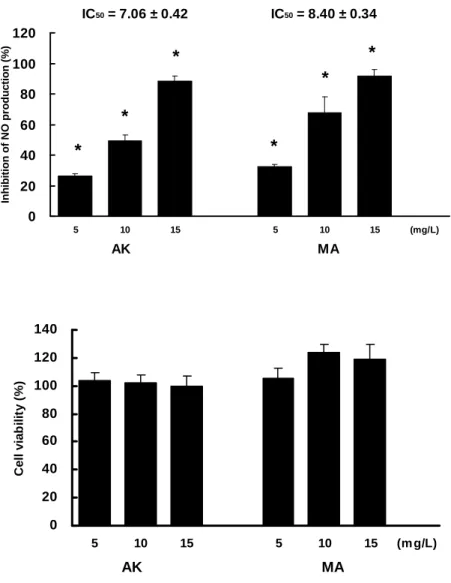
Inhibitory Effect of MA and AK on LPS-induced NO Production in RAW 264.7 Cells. To study the anti-inflammatory effects of MA and AK on LPS-stimulated
RAW 264.7 cells, we first utilized Griess reagent and MTT assay to estimate the NO
azaphilone derivatives reduced NO production in a dose-dependent manner, but did
not alternate cell viability obviously. The anti-NO activity of MA (IC50 = 7.06 ± 0.42 μg/mL) was slightly better than that of AK (IC50 = 8.40 ± 0.34 μg/mL). The results suggested that MA and AK were good anti-inflammatory agents, which was suitable for further anti-inflammatory studies at dosage of 10 μg/mL.
Suppression of LPS-induced iNOS/COX-2 Expression by MA and AK. RAW
264.7 cells were treated with 1 μg/mL of LPS alone, or in combinations with 10 μg/mL of MA and AK for 24 hr. The alternation of iNOS and COX-2 expression were detected by Western blot (Figure 7). Treatment of MA and AK exhibited a remarkable
decrease of iNOS expression, which was regarded as a key enzyme directly
responsible for NO production. As mention to COX-2, treatment of MA or AK
showed only a slight decrease of protein expression. Nevertheless, MA inhibited
iNOS and COX-2 expressions obviously compared with AK.
DISCUSSION
RMR was recognized to posses various biological functions, but in most cases, the
mechanisms remained unclear. In order to explore the mechanisms for each biological
function, the preparation of large-scale bioactive components were needed. [An
available as lovastatin.] Generally speaking, the study of anti-tumor properties of
RMR were started from the extracts, and then narrowed down to its major
components. For example, treatments of RMR extract not only mitigated oral
carcinogenesis in hamster (17), but also prevented from neovascularization and
intravasation of malignant cell by chicken embryo model (23). One of the major
anti-tumor components from RMR was MK, which inhibited metastasis and tumor
progression of Lewis lung carcinoma cell (16). Similarly, treating on colon cancer cell
line Caco-2, MK induced apoptosis (24) and exerted some proteomic changes (25). As
to the study of Monascus pigments, treatment of pigment-rich fraction from RMR also
induced apoptosis on colon cancer cells (26). Besides, a scarce but critical paper,
reported about the purification and treatment of AK to induce apoptosis on human
liver cancer cell line Hep G2 (27). In our recent research, we have successfully
developed the procedures of purifying a series of yellow pigment derivatives from the
fermented products of M. purpureus NTU 568, and further proved their cytotoxicity to
cancer cells, but not to normal cells (18). The above experiences are useful for us to
execute large-scale preparation of related known or novel derivatives, and investigate
the interesting biological mechanisms, such as apoptosis-related and
anti-inflammatory properties.
apoptotic mechanisms, including caspase-9 mediated intrinsic pathway and caspase-8
mediated extrinsic pathway (28, 29). Our first results were that AK and MA induced
caspase-3 activations, which were recognized a down-steam consequence of apoptosis
(Figure 4). This outcome was consistent with a reported data, which showed AK
increased the amounts of apoptotic cells with cycle cell analysis (27). Further study of
up-stream events for caspase-3, we also demonstrated AK and MA induced
casepase-9 activations through the intrinsic pathway. However, the situation of MA
was much more complicated. We found MA activated not only caspase-9 but also
caspase-8. Generally, caspase-8 was indicated to activate caspase-9 through t-Bid (30,
31), but our data showed that MA-induced activations of caspase-9 were earlier and
stronger than that of caspase-8 (Figure 5). This result suggested that apoptosis
induced by MA was major through caspase-9 mediated intrinsic pathway, but minor
through caspase-8 mediated extrinsic pathway. In this case, the activations of
caspase-9 were not amplified by caspase-8. Thus, the activations of caspase-8 were
possibly triggered by clustering of death receptors (32, 33), but not a direct binding of
ligand to death receptor.
Moreover, we utilized the isolated AK and MA to explore their anti-inflammatory
properties, such as inhibition of pro-inflammatory factors or enzymes. As shown in
MA. This result was consistent with our previous published paper, which describing
that RMR extracts significantly decreased the NO production and pro-inflammatory
cytokines (17). Further exploring the up-stream events for NO production, we
analyzed the protein expressions of iNOS and COX-2. Therefore, we found that the
expression of iNOS was severely decreased with the treatment of AK and MA,
whereas the expression of COX-2 was just slightly decreased with the same treatment.
This result was consistent with our previous report; however in which experiments
were performed by treating with RMR extracts, not purified component (17). The
evidences implied that the anti-inflammatory properties of AK, MA and RMR extracts
were major resulted from inhibition of iNOS, but minor from that of COX-2. It was
possible that these yellow pigment derivatives inhibited iNOS and then reduced some
NO-mediated responses, such as NO-stimulated COX-2 expression via p38-dependent
pathway (34) or blockage of the self-deactivation of COX-2 (35).
Previous studies in our group showed that RMR extracts or red mold dioscorea
(RMD) extracts fermented from M. purpureus NTU 568 might prevent the
DMBA-induced oral carcinogenesis in a hamster model (17, 36). Studies from other
group also showed that monascin possessed anti-tumor-initiating effects on mice skin
(11). These studies have indicated that the anti-tumor properties of fermented products
pigments. We have also investigated caspase activations and iNOS/COX-2 inhibitions
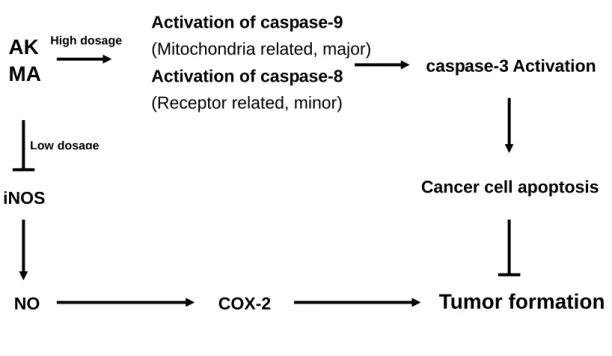
with treatments of yellow pigment derivates. In a conclusion, we purposed that yellow
pigment derivates, such as AK and MA, might inhibit tumor formation by inducing
apoptosis of cancer cells or through anti-inflammatory effects (Figure 8). These
results strongly implied that RMR or RMD fermented from M. purpureus NTU 568
were potential candidates for tumor prevention due to the fermented products of M.
purpureus NTU 568 containing available amount of yellow pigments.
ACKNOWLEDGMENT
The research grant was supported by the National Science Council (NSC) and
National Research Institute of Chinese Medicine (NRICM), Taiwan, the Republic of
China.
LITERATURE CITED
(1) Lee, C. L.; Tsai, T. Y.; Wang, J. J.; Pan, T. M. In vivo hypolipidemic effects and
safety of low dosage Monascus powder in a hamster model of hyperlipidermia.
Appl. Microbiol. Biotechnol. 2006, 70, 533-540.
(2) Wang, J. J.; Shieh, M. J.; Kuo, S. L.; Lee, C. L.; Pan, T. M. Effect of red mold rice
exercise. Appl. Microbiol. Biotechnol. 2006, 70, 247-253.
(3) Lee, C. L.; Kuo, T. F.; Wu, C. L.; Wang, J. J.; Pan, T. M. Red mold rice promotes
neuroprotective sAPPalpha secretion instead of Alzheimer’s risk factors and
amyloid beta expression in hyperlipidemic Aβ40-infused rats. J. Agric. Food
Chem. 2010, 58, 2230-2238.
(4) Chen, W. P.; Ho, B. Y.; Lee, C. L.; Lee, C. H.; Pan, T. M. Red mold rice prevents
the development of obesity, dyslipidemia and hyperinsulinemia induced by
high-fat diet. Int. J. Obes. 2008, 32, 1694-1704.
(5) Li, Y. G.; Zhang, F.; Wang, Z. T.; Hu, Z. B. Identification and chemical profiling of
monacolins in red yeast rice using high-performance liquid chromatography with
photodiode array detector and mass spectrometry. J. Pharm. Biomed. Anal. 2004,
35, 1101-1112.
(6) Journoud, M.; Jones, P. J. Red yeast rice: a new hypolipidemic drug. Life Sci.
2004, 74, 2675-2683.
(7) Tsuji, K.; Ichikawa, T.; Tanabe, N.; Obata, H.; Abe, S.; Tarui, S.; Nakagawa, Y.
Extraction of hypotensive substance from wheat beni-koji. Nippon Shokuhin
Kogyo Gakkaishi 1992, 39, 913-918.
(8) Wu, C. L.; Lee, C. L.; Pan, T. M. Red mold dioscorea has a greater
hypertensive rats. J. Agric. Food Chem. 2009, 57, 5035-5041.
(9) Aniya, Y.; Yokomakura, T.; Yonamine, M.; Shimada, K.; Nagamine, T.;
Shimabukuro, M.; Gibo, H. Screening of antioxidant action of various molds and
protection of Monascus anka against experimentally induced liver injuries of rats.
Gen. Pharmacol. 1999, 32, 225-231.
(10) Akihisa, T.; Tokuda, H.; Yasukawa, K.; Ukiya, M.; Kiyota, A.; Sakamoto, N.;
Suzuki, T.; Tanabe, N.; Nishino, H. Azaphilones, furanoisophthalides, and amino
acids from the extracts of Monascus pilosus-fremented rice (red-mold rice) and
their chemopreventive effects. J. Agric. Food Chem. 2005, 53, 562-565.
(11) Akihisa, T.; Tokuda, H.; Ukiya, M.; Kiyota, A.; Yasukawa, K.; Sakamoto, N.;
Kimura, Y.; Suzuki, T.; Takahasu, J.; Nishino, H. Anti- tumor-initiating effects of
monascin, an azaphilonoid pigment from the extract of Monascus pilosus
fermented rice (red-mold rice). Chem. Biodivers. 2005, 2, 1305-1309.
(12) Jung, H.; Kim, C.; Kim, K.; Shin, C. S. Color characteristics of Monascus
pigments derived by fermentation with various amino acids. J. Agric. Food Chem.
2003, 51, 1302-1306.
(13) Kim, C.; Jung, H.; Kim, Y. O.; Shin, C. S. Antimicrobial activities of amino acid
derivatives of Monascus pigments. FEMS Microbio. Lett. 2006; 264:117-124.
Development of lipase inhibitors from various derivatives of Monascus pigment
produced by Monascus fermentation. Food Chemistry 2007, 101, 357-364.
(15) Jeun, J.; Jung, H.; Kim, J. H.; Kim, Y. O.; Youn, S. H.; Shin, C. S. Effect of
Monascus pigment threonine derivative on regulation of the cholesterol level in
mice. Food Chemistry 2008, 107, 1078-1085.
(16) Ho, B. Y.; Pan, T. M. The Monascus metabolite monacolin K reduces tumor
progression and metastasis of Lewis lung carcinoma cells. J. Agric. Food Chem.
2009, 57, 8258-8265.
(17) Tsai, R. L.; Ho, B. Y.; Pan, T. M. Red mold rice mitigates oral carcinogenesis in
7,12-dimethyl-1,2-benz[a]anthracene-induced oral carcinogenesis in hamster. Evid.
Based Complement Alternat. Med. 2009, Advance access published on December
17.
(18) Hsu, Y. W.; Hsu, L. C.; Liang, Y. H.; Kuo, Y. H.; Pan, T. M. Monaphilones A-C,
three new antiproliferative azaphilone derivatives from Monascus purpureus. J.
Agric. Food Chem. 2010, 58, 8211-8216.
(19) Edmondson, J. M.; Armstrong, L. S.; Martinez, A. O. A rapid and simple
MTT-based spectrophotometric assay for determining drug sensitivity in
monolayer cultures. Methods Cell Sci. 1988, 11, 15-17.
Methods Mol. Biol. 1998, 79, 179-183.
(21) Fiddler, R. M. Collaborative study of modified AOAC method of analysis for
nitrite in meat and meat products. J. AOAC. 1977, 60, 594-599.
(22) Frankfurt, O. S.; Krishan, A. Enzyme-linked-immunosorbent assay (ELISA) for
the specific detection of apoptotic cells and its application to rapid drug screening.
J. Immunol. Methods. 2001, 153, 133-143.
(23) Ho, B. Y.; Wu, Y. M.; Hsu, Y. W.; Hsu, L. C.; Kuo, Y. H.; Chang, K. J.; Pan, T. M.
Effects of Monascus-fermented rice extract on malignant cell-associated
neovascularization and intravasation determined using the chicken embryo
chorioallantoic membrane model. Integr. Cancer Ther. 2010, 2, 204-212.
(24) Lin, W. Y.; Hsu, W. Y.; Hish, C. H.;Pan, T. M. Proteome changes in Caco-2 cells
treated with Monascus-fermented red mold rice extract. J. Agric. Food Chem.
2007, 55, 8987-8994.
(25) Lin, W. Y.; Song, C. Y.; Pan, T. M. Proteomic analysis of Caco-2 cells treated
with monacolin K. J. Agric. Food Chem. 2006, 54, 6192-6200.
(26) Hong, M. Y.; Seeram, N. P.; Zhang, Y.; Heber, D. Anticancer effects of Chinese
red yeast rice versus monacolin K alone on colon cancer cells. J. Nutr. Biochem.
2008, 19, 448-458.
Monascus-fermented red rice exhibits selective cytotoxic effect and induces cell
death on Hep G2 cells. J. Agric. Food Chem. 2005, 53, 1949-1954.
(28) Budihardjo, I.; Oliver, H.; Lutter, M.; Luo, X.; Wang, X. Biochemical pathways
of caspase activation during apoptosis. Annu. Rev. Cell Devel. Biol. 1999, 15,
269-290.
(29) Hengartner, M. O. The biochemistry of apoptosis. Nature 2000, 407, 770-776.
(30) Eskes, R.; Desagher, S.; Antonsson, B.; Martinou, J. Bid induces the
oligomerization and insertion of Bax into the outer mitochondrial membrane. Mol.
Cell Biol. 2000, 20, 929-935.
(31) Desagher, S.; Osen-Sand, A.; Nichols, A.; Eskes, R.; Montessuit, S.; Lauper, S.;
Maundrell, K.; Antonsson, B.; Martinou. J. Bid-induced conformational change of
Bax is responsible for mitochondrial cytochrome c release during apoptosis. J.
Cell Biol. 1999, 144, 891-901.
(32) Stel, A. J.; Ten Cate, B.; Jacobs, S.; Kok, J. W.; Spierings, D. C.; Dondorff, M.;
Helfrich, W.; Kluin-Nelemans, H. C.; de Leij, L. F.; Withoff, S.; Kroesen, B. J. Fas
receptor clustering and involvement of the death receptor pathway in
rituximab-mediated apoptosis with concomitant sensitization of lymphoma B cells
to Fas-induced apoptosis. J. Immunol. 2007, 178, 2287-2295.
Robinson, H.; Salvesen, G. S.; Schwarzenbacher, R.; Riedl, S. J. The Fas-FADD
death domain complex structure unravels signalling by receptor clustering. Nature,
2009, 457, 1019-1022.
(34) Cheng, H. F.; Zhang, M. Z.; Harris, R. C. Nitric oxide stimulates
cyclooxygenase-2 in cultured cTAL cells through a p38-dependent pathway. Am. J.
Physiol. Renal Physiol. 2006, 290, 1391-1397.
(35) Egan, R. W.; Paxton, J.; Kuehl, F. A. Mechanism for irreversible
self-deactivation of prostaglandin synthetase. J. Biol. Chem. 1976, 251,
7329-7335.
(36) Hsu, W. H.; Lee, B. H.; Pan, T. M. Protection of Monascus-fermented dioscorea
against DMBA-induced oral injury in hamster by anti-inflammatory and
Figure legends
Figure 1. (A) Structures of monaphilone A (MA) and ankaflavin (AK). (B) HPLC
analysis and UV spectrum of purified MA and AK showed their purities were more
than 98%. Purified MA and AK were detected at 236 nm.
Figure 2. Inhibition of HEp-2 cell viability by AK and MA. (A) HEp-2 cells were
treated with test agents for 72 hr. (B) HEp-2 cells were treated with 50 μg/mL of test agents for 24, 48 and 72 hr. Data were expressed as means ± SD (n = 3). *,
significantly different (p < 0.05) versus the negative control (without any treatment).
Figure 3. Detection of early apoptosis of HEp-2 cells treated with AK and MA by
ssDNA ELISA kit. HEp-2 cells were treated with CPT (0.2 μg/mL, reference compound for cytotoxity), AK (50 μg/mL) and MA (50 μg/mL) for 12 hr. (PC, positive control (0.3 μg/mL of ssDNA); NC, negative control; CPT, camptothecin) Data were expressed as means ± SD (n = 3). *, significantly different (p < 0.05)
versus the positive control. #, significantly different (p < 0.05) versus the negative
control.
caspase-3 and β-actin were detected by Western blot. HEp-2 cells were treated with 25 or 50 μg/mL of test agents for 12 or 24 hours. (A) From the left side: Lane 1, control; lane 2, AK 25 μg/mL for 24 hr; lane 3, AK 50 μg/mL for 24 hr; lane 4, AK 50 μg/mL for 12 hr; lane 5, MA 25 μg/mL for 24 hr; lane 6, MA 50 μg/mL for 24 hr; lane 7, MA 50 μg/mL for 12 hr. (B) Quantification of cleaved caspase-3 presented above. Data were expressed as means ± SD (n = 3). *, significantly different (p < 0.01)
versus the control (without any treatment).
Figure 5. The effects of MA and AK on caspase-9 and -8 activities. HEp-2 cells were
treated with 50 μg/mL of test agents for 12 or 24 hours. From the left side were: control; MA, 12 hr; MA, 24 hr; AK, 12 hr; AK, 24 hr. Data were expressed as means
± SD (n = 3). *, significantly different (p < 0.01) versus the control (without any
treatment).
Figure 6. AK and MA suppressed LPS-induced NO production on RAW 264.7 cells.
Cells were treated with LPS (1 μg/mL) and in combinations with AK (10 μg/mL) or MA (10 μg/mL) for 24 hr. Data were expressed as means ± SD (n = 3). *, significantly different (p < 0.05) versus the negative control (treatment only with LPS).
Figure 7. AK and MA suppressed LPS-induced inflammatory iNOS/COX-2
expression on RAW 264.7 cells. Cells were treated with test agents for 24 hr and
detected for iNOS/COX-2 by Western blot. (A) From the left side: Lane 1, control; lane 2, LPS (1 μg/mL); lane 3, LPS (1 μg/mL) and AK (10 μg/mL); lane 4, LPS (1 μg/mL) and MA (10 μg/mL). (B) Quantification of iNOS/COX-2 expression presented above. Data were expressed as means ± SD (n = 3). *, significantly different
(p < 0.05); #, significantly different (p < 0.01).
Figure 8. The correlations of AK and MA inhibited tumor formation via apoptosis of
Monaphilone A Ankaflavin
Figure 1. (A) Structures of monaphilone A (MA) and ankaflavin (AK). (B) HPLC
analysis and UV spectrum of purified MA and AK showed their purities were more
than 98%. Purified MA and AK were detected at 236 nm. Minutes 0 5 10 15 20 25 30 mA U 0 200 400 0 2 4 DAD-236 nm Monaphilone B Monaphilone B30-70(0-20min)ACN.met.dat Retention Time Area Minutes 0 5 10 15 20 25 30 mA U 0 200 400 0 20 40 DAD-236 nm Ankaflavin Ankaflavin30-70(0-20min)ACN.met.dat Retention Time Area 1 2 O O CH3 O CH3 O H C H3 3 4 4a 5 6 9 7 14 8 8a 11 12 10 13 13a 1 2 3 4 4a 5 6 9 7 14 8 8a 11 12 10 13 O O CH3 C H3 O O CH3 O A B
0 20 40 60 80 100 120 0 6.25 12.5 25 50 Concentration (μg/mL) Cell viabil it y (% of contr o l) Ankaflavin Monaphilone A
Figure 2. Inhibition of HEp-2 cell viability by AK and MA. (A) HEp-2 cells were
treated with test agents for 72 hr. (B) HEp-2 cells were treated with 50 μg/mL of test agents for 24, 48 and 72 hr. Data were expressed as means ± SD (n = 3). *,
significantly different (p < 0.05) versus the negative control (without any treatment).
0 10 20 30 40 50 24 48 72 Cultured time (hr) Cel l v iab il it y ( % of co nt ro l) Ankaflavin Monaphilone A
*
*
*
*
*
*
A B
0
20
40
60
80
100
120
PC
NC
CPT
AK
MA
E
a
rl
y apopt
osi
s
(
%
)
*
#Figure 3. Detection of early apoptosis of HEp-2 cells treated with AK and MA by
ssDNA ELISA kit. HEp-2 cells were treated with CPT (0.2 μg/mL, reference compound for cytotoxity), AK (50 μg/mL) and MA (50 μg/mL) for 12 hr. (PC, positive control (0.3 μg/mL of ssDNA); NC, negative control; CPT, camptothecin) Data were expressed as means ± SD (n = 3). *, significantly different (p < 0.05)
versus the positive control. #, significantly different (p < 0.05) versus the negative
A
B
Figure 4. The effects of MA and AK on caspase-3 activation in HEp-2 cells. Cleaved
caspase-3 and β-actin were detected by Western blot. HEp-2 cells were treated with 25 or 50 μg/mL of test agents for 12 or 24 hours. (A) From the left side: Lane 1, control; lane 2, AK 25 μg/mL for 24 hr; lane 3, AK 50 μg/mL for 24 hr; lane 4, AK 50 μg/mL for 12 hr; lane 5, MA 25 μg/mL for 24 hr; lane 6, MA 50 μg/mL for 24 hr; lane 7, MA 50 μg/mL for 12 hr. (B) Quantification of cleaved caspase-3 presented above. Data were expressed as means ± SD (n = 3). *, significantly different (p < 0.01)
versus the control (without any treatment).
0 100 200 300 400 500
Control AK25-24 AK50-24 AK50-12 MA25-24 MA50-24 MA50-12
R e la ti v e c o nte n ts o f c le a v e d c a s p a s e -3 (% of c o ntr ol )
*
*
*
*
*
*
A 0 50 100 150 200
Control MA12 MA24 AK12 AK24
Caspase-9 ac ti vi ty (% of control )
*
*
*
*
0 50 100 150 200Control MA12 MA24 AK12 AK24
Caspase-8 a
c
tivity (% of control)
*
Figure 5. The effects of MA and AK on caspase-9 and -8 activities. HEp-2 cells were
treated with 50 μg/mL of test agents for 12 or 24 hours. From the left side were: control; MA, 12 hr; MA, 24 hr; AK, 12 hr; AK, 24 hr. Data were expressed as means
0 20 40 60 80 100 120 140 5 10 15 5 10 15 (m g/L) AK MA C e ll viab il it y (% )
Figure 6. AK and MA suppressed LPS-induced NO production on RAW 264.7 cells.
Cells were treated with LPS (1 μg/mL) and in combinations with AK (10 μg/mL) or MA (10 μg/mL) for 24 hr. Data were expressed as means ± SD (n = 3). *, significantly different (p < 0.05) versus the negative control (treatment only with LPS).
IC50 = 7.06 ± 0.42 IC50 = 8.40 ± 0.34 0 20 40 60 80 100 120 5 10 15 5 10 15 (mg/L) AK MA Inhib it ion of N O pr oduc ti on (% ) * * * * * *
B
0 20 40 60 80 100 120 AK MA Rel a ti ve co nt ent s ( % ) iNOS COX-2*
#Figure 7. AK and MA suppressed LPS-induced inflammatory iNOS/COX-2
expression on RAW 264.7 cells. Cells were treated with test agents for 24 hr and
detected for iNOS/COX-2 by Western blot. (A) From the left side: Lane 1, control; lane 2, LPS (1 μg/mL); lane 3, LPS (1 μg/mL) and AK (10 μg/mL); lane 4, LPS (1 μg/mL) and MA (10 μg/mL). (B) Quantification of iNOS/COX-2 expression presented above. Data were expressed as means ± SD (n = 3). *, significantly different
β-actin
iNOS
COX-2
Figure 8. The correlations of AK and MA inhibited tumor formation via apoptosis of
cancer cells or via suppression of inflammations.
AK
MA
iNOS
NO COX-2
Tumor formation
Cancer cell apoptosis Activation of caspase-9
(Mitochondria related, major)
Activation of caspase-8
(Receptor related, minor)
caspase-3 Activation
High dosage