Curcumin enhances the mitomycin C-induced cytotoxicity via downregulation of
MKK1/2
–ERK1/2-mediated Rad51 expression in non-small cell lung cancer cells
Jen-Chung Ko
a,b,c, Min-Shao Tsai
d, Shao-Hsing Weng
d, Ya-Hsun Kuo
d, Yu-Fan Chiu
d, Yun-Wei Lin
d,⁎
a
Department of Internal Medicine, Hsinchu Hospital, Department of Health, The Executive Yuan, Taiwan
bDepartment of Nursing, Yuanpei University, HsinChu, Taiwan; Graduate Institute of Technology Law, National Chiao Tung University, Taiwan c
Graduate Institute of Technology Law, National Chiao Tung University, Taiwan
d
Molecular Oncology Laboratory, Department of Biochemical Science and Technology, National Chiayi University, Chiayi, Taiwan
a b s t r a c t
a r t i c l e i n f o
Article history: Received 1 June 2011 Revised 6 July 2011 Accepted 15 July 2011 Available online 24 July 2011 Keywords:Curcumin Rad51 Mitomycin C Cytotoxicity
Non-small cell lung cancer
Curcumin (diferuloylmethane), a major active component of turmeric (Curcuma longa), has been reported to suppress the proliferation of a wide variety of tumor cells. Rad51 is a key protein in the homologous recombination (HR) pathway of DNA double-strand break repair, and HR represents a novel target for cancer therapy. A high expression of Rad51 has been reported in chemo- or radio-resistant carcinomas. Therefore, in the current study, we will examine whether curcumin could enhance the effects of mitomycin C (MMC), a DNA interstrand cross-linking agent, to induce cytotoxicity by decreasing Rad51 expression. Exposure of two human non-small lung cancer (NSCLC) cell lines (A549 and H1975) to curcumin could suppress MMC-induced MKK1/2–ERK1/2 signal activation and Rad51 protein expression. Enhancement of ERK1/2 activation by constitutively active MKK1/2 (MKK1/2-CA) increased Rad51 protein levels in curcumin and MMC co-treated human lung cells. Moreover, the synergistic cytotoxic effect induced by curcumin combined with MMC was decreased by MKK1-CA-mediated enhancement of ERK1/2 activation by a significant degree. In contrast, MKK1/2 inhibitor, U0126 was shown to augment the cytotoxicity of curcumin and MMC through downregulation of ERK1/2 activation and Rad51 expression. Depletion of endogenous Rad51 expression by siRad51 RNA transfection significantly enhanced MMC and/or curcumin induced cell death and cell growth inhibition. In contrast, an overexpression of Rad51 protected lung cancer cells from synergistic cytotoxic effects induced by curcumin and MMC. We concluded that Rad51 inhibition may be an additional action mechanism for enhancing the chemosensitization of MMC by curcumin in NSCLC.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Lung cancer is a leading cause of morbidity and mortality worldwide. Lung cancer is clinically classified into two histological types, non-small cell lung cancer (NSCLC) and small cell lung cancer (SCLC) (Bhattacharjee et al., 2001). NSCLC is the major type of lung cancers, accounting for approximately 85% of all cases (Landis et al., 1999). The current therapeutic option for advanced NSCLC patients is chemotherapy, and the use of cytotoxic chemotherapy improves median overall survival rate compared with best supportive care (Pfister et al., 2004; Schiller et al., 2002). Mitomycin C (MMC), an anticancer drug, is typically used as afirst or second line regimen to treat NSCLC, and is often combined with other chemotherapeutic agents for advanced NSCLC treatment (Babiak et al., 2007; Booton et al., 2006). MMC causes intrastrand DNA cross-links that
lead to collapsing replication forks and generation of DNA double-strand breaks that are typically repaired by homologous recombination repair (HRR) (Dronkert and Kanaar, 2001; Helleday et al., 2008; Warren and Hamilton, 1996). The therapeutic value of MMC depends on the capability of the cells to remove DNA damage (McHugh et al., 2001).
Mammalian Rad51, the key protein in the HRR pathway, is a strand transferase that forms a nucleoproteinfilament by polymerizing onto single-stranded DNA at the processed DNA break. The nucleoprotein filament promotes DNA strand exchange with the undamaged homologous chromatid (Daboussi et al., 2002; Richardson, 2005; San Filippo et al., 2008). A high level of Rad51 expression is reported for a variety of tumor cell lines (Richardson, 2005) and leads to a worse clinical outcome in lung cancer (Qiao et al., 2005; Takenaka et al., 2007). Elevated Rad51 levels can result in an increased number of chromosomal recombination and rearrangement events, and corre-late with resistance to chemotherapeutic agents in mammalian cells (Arnaudeau et al., 1999; Richardson, 2005; Vispe et al., 1998). Our previous studies have indicated that downregulation of Rad51 expression could increase the chemosensitivity to MMC in NSCLC
Toxicology and Applied Pharmacology 255 (2011) 327–338
⁎ Corresponding author at: Department of Biochemical Science and Technology, National Chiayi University, Chiayi 600, Taiwan. Fax: + 886 5 271 7780.
E-mail address:[email protected](Y.-W. Lin).
0041-008X/$– see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.taap.2011.07.012
Contents lists available atScienceDirect
Toxicology and Applied Pharmacology
j o u r n a l h o m e p a g e : w w w. e l s ev i e r. c o m / l o c a t e / y ta a pcells (Ko et al., 2008). Therefore, overexpression of Rad51 is involved in tumor progression by promoting genome instability and alternative HRR pathway (Richardson et al., 2004).
Curcumin (diferuloylmethane), present in Curcuma longa, has been shown to inhibit cell proliferation in a wide variety of human cancer cell lines in vitro, including inhibition of NSCLC cell prolifer-ation (Aggarwal et al., 2003). For example, the IC50 value at 24 h exposure of curcumin is 40μM for human NSCLC cell line (H1299) (Radhakrishna Pillai et al., 2004). However, the IC50 of curcumin in human lung adenocarcinoma epithelial cell line A549 is 93μM at 24 h treatment (Chen et al., 2010). Moreover, curcumin is known as a potent inhibitor of NF-κB, MAPK family and AP-1 signaling pathways (Chen and Tan, 1998; Collett and Campbell, 2004; Han et al., 2002; Singh and Aggarwal, 1995). The IC50 (50% inhibition concentration) of curcumin is between 5 and 10μM for JNK activation and is 20 μM for ERK1/2 activation (Chen and Tan, 1998). The sensitizing effect of curcumin on cancer cell apoptosis has been reported for several chemotherapeutic agents (Du et al., 2006; Weir et al., 2007). For example, curcumin sensitizes ovarian cancer cells to cisplatin and enhances chemotherapeutic treatment (Chan et al., 2003). However, there is no detailed mechanism to address curcumin's effect on MMC-induced cytotoxicity in human NSCLC cells.
In this study, we hypothesized that Rad51 inhibition by curcumin may be an additional action mechanism for enhancing the chemosensitization of MMC in NSCLC. The results demonstrated that curcumin could downregulate MMC-induced Rad51 expression to decrease cell viability in NSCLC cell lines. Further investigation revealed that curcumin-decreased Rad51 expression was through inactivation of MKK1/2–ERK1/2 in human lung cancer cells. These data suggest that at least one mechanism underlying the mecha-nism of action of curcumin is to mediate the dowregulation of MKK1/2–ERK1/2-Rad51, which contributes to the enhancement of the sensitivity of MMC to NSCLC.
Methods
Cell lines and culture. Human lung squamous cell carcinoma A549, and lung carcinoma H1975 cell lines were obtained from the American Type Culture Collection (Manassas, VA) and cultured at 37 °C in a humidified atmosphere containing 5% CO2in RPMI-1640 complete
medium supplemented with sodium bicarbonate (2.2%, w/v), L -glutamine (0.03%, w/v), penicillin (100 units/mL), streptomycin (100μg/mL), and fetal calf serum (10%).
Drugs and reagents. Curcumin and MMC were obtained from Sigma-Aldrich (St. Louis, MO). N-acetyl-Leu-Leu-norleucinal (ALLN), MG132, and U0126 were purchased from Calbiochem–Novabiochem (San Diego, CA). Curcumin, ALLN, MG132, and U0126 were dissolved in dimethyl sulfoxide (DMSO).
Western blot analysis. After different treatments, equal amounts of proteins from each set of experiments were subjected to Western blot analysis as previously described (Ko et al., 2008). The specific phospho-ERK1/2 (Thr202/Tyr204) and phospho-MKK1/2 (Ser217/Ser221) anti-bodies were purchased from Cell Signaling (Beverly, MA). Rabbit polyclonal antibodies against Rad51 (H-92) (sc-8349), ERK2 (K-23) (sc-153), ubiquitin (P4D1) (sc-8017), and Actin (I-19) (sc-1616) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA).
Immunoprecipitation. After treatment, an equal amount of proteins was immunoprecipitated using an anti-ubiquitin antibody and collected with protein G-Sepharose beads (GE Healthcare). The immunoprecipitate was then washed three times with cold lysis buffer and subjected to immunoblot analysis.
Transfection with small interfering RNA (siRNA). Plasmids transfection of MKK1-CA (a constitutively active form of MKK1,ΔN3/S218E/S222D) and MKK2-CA (a constitutively active form of MKK2,ΔN4/S222E/S226D) was achieved as previously described (Ko et al., 2008). Exponentially growing human lung cancer cells (106) were plated for 18 h, and then
the MKK1/2-CA expression vectors were transfected into cells using Lipofectamine (Invitrogen) for 24 h before combined treatment with curcumin and MMC in RPMI-1640 complete medium.
Quantitative real-time PCR. PCRs were performed using an ABI Prism 7900HT according to the manufacturer's instructions. Amplification of specific PCR products was detected using the SYBR Green PCR Master Mix (Applied Biosystems). For each sample, the data were normalized to the housekeeping gene Glyceraldehyde 3-phosphate dehydrogenase (gapdh). The designed primers in this study were: rad51 forward primer, 5′-CCCATTTCACGGTTAGAGCAG-3′; rad51 reverse primer, 5′-TAGCTCCTTCTTTGGCGCATA-3′; gapdh forward primer, 5′-CAT-GAGAAGTATGACAACAGCCT-3′; gapdh reverse primer, 5′-AGTCCTTCCACGATACCAAAGT-3′. Analysis was performed using the comparative Ct value method.
Reverse transcription-PCR (RT-PCR). Total RNA was extracted from drug-treated NSCLC cells with TRIzol reagent (Invitrogen, Carlsbad, CA). cDNA was synthesized by using random hexamers following the Moloney murine leukemia virus reverse transcriptase cDNA synthesis system (Invitrogen). Thefinal cDNA was used for the subsequent PCRs (Ko et al., 2009).
Measurement of cell viability. In vitro MTS assay was carried out essentially according to the procedure described (Tsai et al., 2010). Briefly, cells were cultured at 5000 per well in 96-well tissue culture plates. To assess cell viability, drugs were added after plating. At the end of the culture period, 20μL of MTS solution (CellTiter 96 Aqueous One Solution Cell Proliferation Assay; Promega, Madison, WI, USA) was added; the cells were incubated for a further 2 h, and the absorbance was measured at 490 nm using an ELISA plate reader (Biorad Technologies, Hercules, CA).
Trypan blue dye exclusion assay. Cells were treated with curcumin and/or MMC for 24 h. In each preparation, the cell death of A549 or H1975 cells was determined by trypan blue dye exclusion assay. This dye can be excluded from living cells, but can penetrate dead cells. The proportion of dead cells was determined using a hemocytometer to count the number of cells stained with trypan blue.
Colony-forming ability assay. Immediately after treating the drugs, cells were washed with phosphate-buffered saline and trypsinized for the determination of cell numbers. The cells were plated at a density of 500–5000 cells on a 60 mm diameter Petri dish in triplicate for each treatment. The cells were cultured for 10–14 days, and the cell colonies were stained with 1% crystal violet solution in 30% ethanol. Cytotoxicity was determined by the number of colonies in the treated cells divided by the number of colonies in the untreated control. Combination index analysis. The cytotoxicity induced by combined treatment with curcumin and MMC was compared with the cytotoxicity induced by each drug using the combination index (CI), where CIb0.9, CI=0.9 to 1.1, and CIN1.1 indicated synergistic, additive, and antagonistic effects, respectively. Combination index analysis was performed using CalcuSyn software (Biosoft, Oxford, UK). The CI values at fraction affected (FA) of 0.5 were averaged for each experiment, and the value was used to calculate the mean between three independent experiments.
Statistical analyses. For each protocol, three or four independent experiments were performed. Results were expressed as the mean ±
standard error of the mean (SEM). Statistical calculations were performed using SigmaPlot 2000 (Systat Software, San Jose, CA). Differences in measured variables between experimental and control groups were assessed by unpaired t-test. Pb0.05 was considered statistically significant.
Results
MMC induces MKK1/2–ERK1/2 signal activation and Rad51 protein expression in A549 and H1975 cells
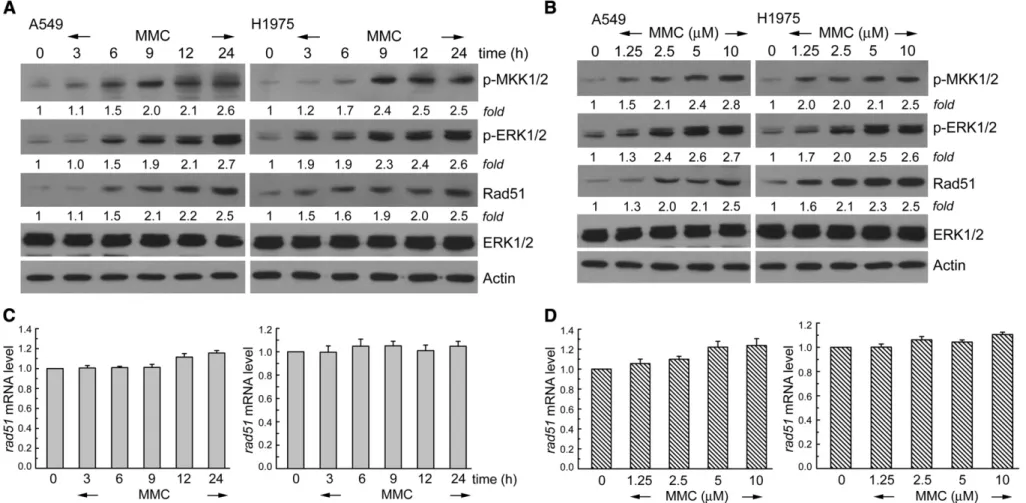
Human NSCLC cell lines, A549 and H1975, were exposed to various concentrations of MMC for 3–24 h. Phosphorylation of MKK1/2–ERK1/2 was determined by Western blot analysis using antibodies specific to phospho-MKK1/2 and phospho-ERK1/2. InFigs. 1A and B, exposure to MMC increased the phosphorylated MKK1/2 and ERK1/2 levels in time and dose-dependent manners, whereas endogenous ERK1/2 protein levels did not change in each treatment. Interestingly, it was found that MMC treatment could increase the protein level of Rad51 in A549 and H1975 cells (Figs. 1A and B). To elucidate whether the observed MMC stimulation of Rad51 protein expression occurred at the transcriptional level, MMC (10μM) was added to A549 or H1975 cells for 3–24 h. Total RNA was isolated and subjected to real-time PCR analysis for rad51 transcripts. As shown in Figs. 1C and D, MMC treatment did not significantly affect rad51 mRNA level in A549 or H1975 cells. Therefore, MMC at low doses of 1.25–10 μM could increase Rad51 protein, but not mRNA level in A549 and H1975 cells.
Curcumin suppresses MMC-elicited phosphorylated MKK1/2–ERK1/2, and Rad51 protein levels
To elucidate whether the observed MMC stimulation of Rad51 protein expression could be affected by curcumin combination, the various concentrations of MMC were co-added with curcumin to A549 or H1975 cells for 24 h. As shown inFig. 2A, curcumin was found to decrease the MMC-elicited upregulation of Rad51 protein expression and phosphorylation of MKK1/2–ERK1/2 in A549 and H1975 cells. In
Fig. 2B, curcumin treatment alone could also induce the down-regulation of Rad51 protein and MKK1/2–ERK1/2 activation in A549 and H1975 cells in a dose-dependent manner. Furthermore, we examined the changes in mRNA levels of rad51 after treatment with curcumin and MMC combination by using real-time PCR and RT-PCR analyses. InFigs. 2C and D, it could be seen that the combination of MMC and curcumin produced significantly lower rad51 mRNA levels relative to MMC alone. Taken together, these results indicated that curcumin-decreased MMC-elicited ERK1/2 activation as well as protein and mRNA expression of Rad51.
26S proteasome inhibitors, MG132 or ALLN, restores the curcumin and MMC combination-induced downregulation protein expression of Rad51 To investigate the role of proteasome-mediated degradation of Rad51 induced by curcumin and MMC, the 26S proteasome inhibitors, MG132 or ALLN, was added for the final 6 h before harvesting curcumin and MMC co-exposed A549 or H1975 cells. As shown in
Fig. 3A, both MG132 and ALLN restored the decreased Rad51 protein levels induced by co-treatment with curcumin and MMC. These results revealed that curcumin-decreased Rad51 protein level in cells treated with MMC was through the 26S proteasome-mediated proteolysis mechanism. In addition, we examined the levels of ubiquitin conjugates on Rad51. Curcumin co-treatment significantly increased the levels of ubiquitin conjugated-Rad51 in MMC-treated NSCLC cell lines, which were compared with MMC alone (Fig. 3B). The results revealed that decreased Rad51 protein level induced by curcumin was through the degradation of Rad51 by the ubiquitin-mediated 26S proteasome pathway in MMC-treated NSCLC cells.
Influence of ERK1/2 activation on the regulation of Rad51 mRNA and protein expression in curcumin and MMC co-treated NSCLC cells
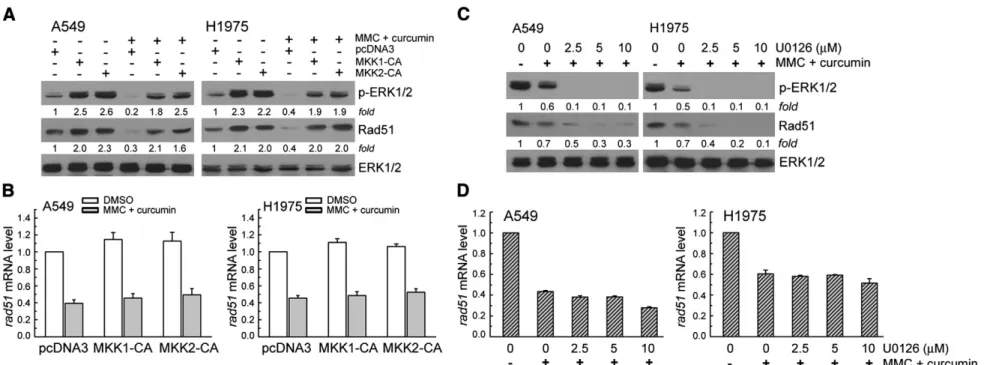
Next, we examined whether the ERK1/2 signaling pathway was involved in regulation of Rad51 protein expression in curcumin and MMC co-exposed A549 or H1975 cells. The cells were transient transfected with a plasmid carrying MKK1-CA or MKK2-CA, a constitutively active form of MKK1/2. Consistent with our previous studies, MKK1-CA or MKK2-CA vectors transfection could increase cellular ERK1/2 phosphorylation and Rad51 protein levels (Fig. 4A). In addition, expression of MKK1/2-CA restored the decrease of Rad51 protein and phosphorylation of ERK1/2 in cells co-treated with curcumin and MMC (Fig. 4A). Furthermore, the mRNA levels of Rad51 affected by MKK1-ERK1/2 signaling pathway was investigated by real-time PCR. InFig. 4B, MKK1-CA vector transfection did not affect Rad51 mRNA in curcumin and MMC co-treated-A549 or H1975 cells as compared to pcDNA3 transfected cells; therefore, ERK1/2 signaling pathway did not affect rad51 transcript in this condition. Moreover, various concentrations of the MKK1/2 inhibitor U0126 (2.5–10 μM) were co-added in curcumin and MMC-treated A549 or H1975 cells for 24 h. As shown inFig. 4C, ERK1/2 phosphorylation and Rad51 protein expression were significantly further decreased by curcumin and MMC in U0126 co-exposed lung cancer cells compared with DMSO control. In addition, U0126 treatment could demonstrate a slightly further downregulation for rad51 mRNA expression in curcumin and MMC co-treated-A549 and H1975 cells, which were analyzed by real-time PCR (Fig. 4D). In summary, inhibition of ERK1/2 activity by curcumin reduced Rad51 protein expression in MMC-exposed NSCLC cells.
Curcumin potentiates cytotoxic effect induced by MMC in NSCLC cells Next, we investigated the effect of a combination of curcumin and MMC on cell viability. A549 and H1975 cells were treated with curcumin (10–40 μM), MMC (2.5–10 μM) or the two-drug combina-tion for 24 h. The viable cells were evaluated by MTS and trypan blue exclusion assay. InFigs. 5A and C, curcumin plus MMC treatment for 24 h resulted in more loss of cell viability than either curcumin or MMC alone in A549 and H1975 cells. Using drug combination effect analysis, the mean values of the combination index (CI) indicated that the curcumin–MMC combination has a synergistic effect (Fig. 5B). Moreover, NSCLC (A549, and H1975) cancer cells were exposed to curcumin and/or MMC and their proliferation was determined over 1– 4 days after exposure to the drugs. Curcumin (20μM) or MMC (1 μM) alone has an antiproliferative effect on NSCLC. However, the combination of curcumin and MMC together was more effective than either drug alone (Fig. 5D). In summary, synergism using a combination of MMC and curcumin was observed in human lung cancer cell lines.
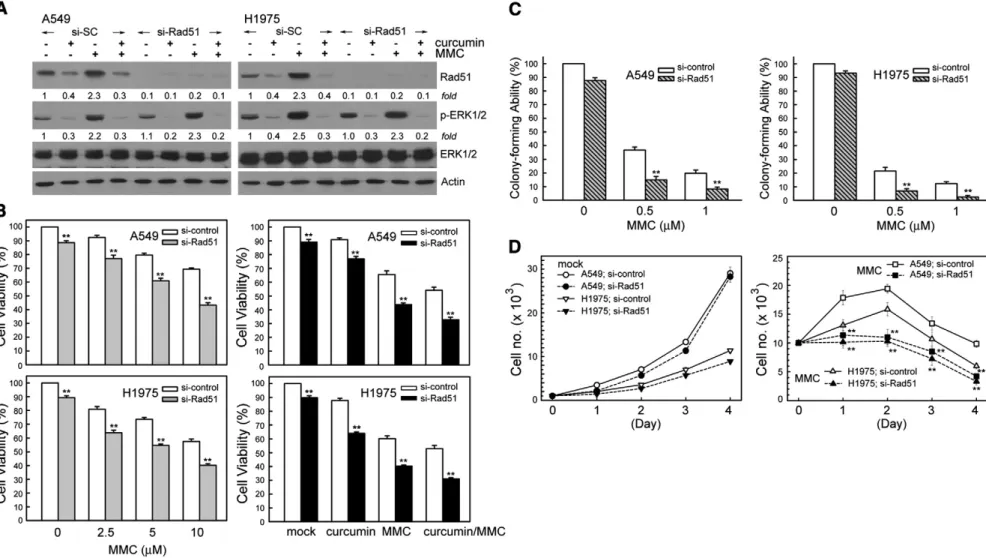
Knockdown Rad51 enhances the cytotoxic effect of MMC in combination with curcumin
To verify the involvement of Rad51 in the cytotoxicity induced by curcumin or/and MMC, the Rad51 protein expression was knocked down using specific siRNA duplexes. As shown inFig. 6A, Rad51 siRNA resulted in reduction of Rad51 protein level in curcumin and/or MMC-treated A549 and H1975 cells. In contrast, phospho-ERK1/2 protein levels were not changed by similar treatment (Fig. 6A). Suppression of Rad51 protein expression by si-Rad51 RNA resulted in A549 or H1975 cells enhancing the cytotoxicity caused by MMC or/and curcumin significantly as compared to si-control transfected cells. This was assessed using MTS assay (Fig. 6B). Given the effect of curcumin on Rad51 expression, we next performed colony-forming assays to investigate whether modifying Rad51 would affect long-term clono-genic cell survival based on inhibited Rad51 expression. We observed
329 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327–338
Fig. 1. MMC increases phospho-MKK1/2, phospho-ERK1/2, and Rad51 protein levels in A549 and H1975 cells. (A) A549 or H1975 cells were cultured in complete medium for 18 h and then exposed to MMC (10μM) for 3–24 h in complete medium. (B) Cells were exposed to various concentrations of MMC (1.25–10 μM) for 24 h. After treatment, cell extracts were examined by Western blot for the determination of phospho-MKK1/2, phospho-ERK1/2, Rad51, and ERK1/2 protein levels. (C and D) After the treatment as A and B, total RNA was isolated and subjected to real-time PCR for rad51.
330 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327 – 338
Fig. 2. Curcumin decreases basal or MMC-induced phospho-MKK1/2–ERK1/2 and Rad51 protein levels in human lung cancer cells. (A) A549 or H1975 cells were cultured in complete medium for 18 h and then exposed to MMC (2.5–10 μM) and curcumin (20μM) for 24 h in complete medium. (B) Cells were treated with curcumin (5–40 μM) for 24 h. After treatment, cell extracts were examined by Western blot for the determination of phospho-ERK1/2, phospho-MKK1/2, Rad51, and ERK1/2 protein levels. (C and D) After the treatment as (A), total RNA was isolated and subjected to RT-PCR and real-time PCR for Rad51.
331 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327 – 338
decreased cell survival by ~21% and by ~ 14% in 0.5μM MMC-exposed A549 or H1975 cells following Rad51 siRNA treatment repeatedly (Fig. 6C). In trypan blue exclusion experiments, we observed that specific downregulation of Rad51 by siRNA enhanced MMC-induced NSCLC cell growth inhibition (Fig. 6D).
Curcumin and/or MMC-induced synergistic cytotoxic effect is abrogated in lung cancer cells with Rad51 overexpression
To strengthen the protective role for Rad51 from cytotoxicity induced by MMC and curcumin, a Flag-Rad51 vector was transfected into A549 or H1975 cells. Then these cells were incubated with MMC in the presence or absence of curcumin. Cell viability was analyzed using the MTS assay (Figs. 7A and B). Treatment of H1975 cells with MMC (10μM) and curcumin (40 μM) decreased cell viability by ~45%. Interestingly, the induction of cytotoxicity by MMC and curcumin combination was significantly reduced (by ~30%) following the overexpression of Rad51 (Fig. 7B). Therefore, Rad51 protected NSCLC cells from the cytotoxicity induced by MMC and curcumin. Influence of MKK1/2–ERK1/2 signal on cytotoxicity induced by curcumin and/or MMC treatment
To evaluate the effects of ERK1/2 on synergistic cytotoxicity induced by curcumin–MMC co-treatment, A549 or H1975 cells were transfected with MKK1/2-CA vectors followed by treatment with curcumin and MMC prior to assessment of MTS assay. Transfection of MKK1/2-CA vector could enhance the cell survival and ERK1/2 activity that was previously suppressed by co-treatment with curcumin and MMC (Fig. 7C andFig. 4A). On the contrary, U0126 (an MKK1/2
inhibitor) markedly enhanced the cytotoxicity in NSCLC cells treated with MMC and curcumin (Fig. 7D). Taken together, these results have shown that the synergistic cytotoxic effect from curcumin and MMC was mainly due to the suppression of ERK1/2 activation.
Discussion
Our results demonstrated for thefirst time that curcumin could decrease the expression of Rad51 upregulated by the MMC in NSCLC cell lines. In mammalian cells, Rad51 is the key recombination protein promoting the pairing and exchange of strands between homologous DNA molecules during HRR (Richardson, 2005). Rad51 interacts with many accessory proteins and is involved in the repair of DNA cross-links that are caused by MMC. It has been shown that chronic hypoxia can decrease the levels of Rad51, leading to reduced levels of HR and increased sensitivity to DNA cross-linking agents in hypoxic tumors (Bristow and Hill, 2008; Chan et al., 2008). Defects in Rad51C expression result in increased cytotoxicity and chromosomal aberra-tions induced by MMC (Godthelp et al., 2002). Increased levels of Rad51 correlate with increased erroneous recombination and resis-tance to DNA-damaging agents in tumor cells (Klein, 2008). Rad51 overexpression leads to a worse clinical outcome in lung cancer (Qiao et al., 2005). In this study, Rad51 downregulation enhanced curcumin and/or MMC-induced cytotoxicity in human lung cancer cells; in contrast, Rad51 overexpression rescued the cytotoxic effect. There-fore, curcumin enhanced MMC-induced cytotoxicity via downregula-tion of Rad51 expression in NSCLC cell lines.
Previous studies indicated that caffeine, which inhibits the Ataxia-telangiectasia mutated (ATM) and ATM- and Rad3-related (ATR) kinases, can abrogate the appearance of Rad51 nuclear foci (Mladenov Fig. 3. 26S proteasome inhibitors restore the decrease Rad51 protein levels in curcumin and MMC-exposed NSCLC cells. (A) Curcumin (40μM) and MMC (2.5 μM) was co-added to A549 or H1975 cells for 18 h. Cells were then co-treated with MG132 (10μM) or ALLN (10 μM) for 6 h. Whole cell extracts were collected for Western blot analysis. (B) As described above, MMC and/or curcumin treated cells were incubated with MG132 (10μM) for 6 h before collection of whole cell extracts. Equal amounts of proteins in each cell extract were subjected to immunoprecipitation (IP) using an anti-ubiquitin or anti-Rad51 antibody. The immunoprecipitates were analyzed by immunoblot (IB) analysis using an anti-Rad51 or anti-ubiquitin antibody.
Fig. 4. Overexpression of MKK1/2-CA restores curcumin-suppressed ERK1/2 activation and Rad51 expression induced by MMC. (A) MKK1-CA expression vectors were transfected into A549 or H1975 cells using lipofectamine. After expression for 24 h, the cells were treated with curcumin (40μM) and MMC (2.5 μM) as described in the legend ofFig. 2. (B) After the treatment described above, total RNA was isolated and subjected to real-time PCR for rad51. (C) The cells were pretreated with various concentrations of MKK1/2 inhibitor U0126 (2.5–10 μM) for 1 h, and then co-treated with MMC (2.5 μM) and curcumin (10 μM) for 24 h. Western blot analysis was performed using specific antibodies against Rad51, phopho-ERK1/2, and ERK1/2. (D) After the treatment described above, total RNA was isolated and subjected to real-time PCR for rad51.
333 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327 – 338
Fig. 5. Curcumin co-treated with MMC synergistically enhances cytotoxicity. (A) Left panel, MMC (2.5–10 μM) and curcumin (10 μM) were co-added to A549 or H1975 cells for 24 h. Right panel, cells were treated with different concentrations of curcumin and MMC (2.5μM) for 24 h. Cytotoxicity was determined by assessment of MTS assay. (B) The mean CI values of curcumin–MMC combination. CI values were averaged for each experiment, and that value was used to calculate the mean between experiments, as described underMethods. Points and columns, mean values obtained from three independent experiments; bars, S.E. (C) At the end of treatment as A, unattached and attached cells were collected and stained with trypan blue dye. The numbers of stained cells (dead) were manually counted. Columns, percentage of trypan blue-positive cells, representing a population of dead cells; bar, standard error (SE) from three independent experiments. The ** respectively denote pb0.01 using Student's t-test for the comparison between the cells treated with curcumin/MMC alone or MMC-curcumin co-treatment. (D) Cells were treated with MMC (1 μM) and/or curcumin (20 μM) for 1– 4 days, after which viable cells were determined by MTS assay. **pb0.01 using Student's t-test for comparison between cells treated with curcumin or MMC alone or with curcumin–MMC co-treatment.
334 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327 – 338
Fig. 6. Knockdown of Rad51 expression by siRNA transfection enhances cytotoxic effects induced by MMC or/and curcumin. (A) A549 or H1975 cells were transfected with siRNA duplexes (200 nM) specific to Rad51 or scrambled (control) in complete medium for 24 h prior to treatment with MMC (10μM) or/and curcumin (20 μM) in complete medium for 24 h. Whole cell extracts were collected for Western blot analysis using specific antibodies against Rad51, phospho-ERK1/2, ERK1/2, and actin. (B and C) After treatment described above, cytotoxicity was determined by MTS and colony-forming ability assay. (D) After the cells were transfected with si-Rad51 or si-scrambled RNA, the cells (104cells/dish) were
seeded in 60-mm dishes. After 18 h, the cells were treated with or without MMC (1μM) for 1–4 days. The surviving cells were counted after staining with trypan blue for each of these days, in triplicate. The results (mean±SEM) were from four independent experiments. ** denotes pb0.01, using the Student's t-test for the comparison between cells treated with MMC in si-Rad51 RNA or si-scrambled RNA transfected cells.
335 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327 – 338
Fig. 7. The influence of the MKK1/2–ERK1/2-Rad51 signaling pathway on cell viability in the NSCLC cells treated with MMC and/or curcumin. (A) Cells were transfected with the Flag-Rad51 vector for 24 h, and then the cells were treated with MMC (10μM) and/or curcumin (40 μM) for 24 h. Whole cell extracts were collected for Western blot analysis using specific antibodies against Rad51, phospho-ERK1/2, and ERK1/2. (B) The cytotoxicity was determined by MTS assay. (C) Overexpression of MKK1/2-CA rescued the levels of cytotoxicity induced by MMC and curcumin. Cells were transfected with MKK1/2-CA vectors for 24 h, then the cells were followed by adding MMC (10μM) or/and curcumin (40 μM) for 24 h. The cytotoxicity affected by MKK1/2-CA vector transfection was determined by MTS assay. The results (mean ± SEM) were obtained from four independent experiments. The ** respectively denote pb0.01 using Student's t-test for the comparisons between the cells transfected with MKK1-CA or pcDNA3 vectors. (D) A549 and H1975 cells were exposed to U0126 (5μM), a MKK1/2 inhibitor, and MMC (2.5 μM) and curcumin (10 μM) for 24 h. After treatment, cell viability was determined by MTS assay. **pb0.01 using Student's t-test for comparisons between cells treated with DMSO alone or with U0126 co-treatment.
336 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327 – 338
et al., 2007) and the association of Rad51 with chromatin and the nuclear matrix caused by MMC (Mladenov et al., 2006). Our studies have shown that the upregulation of Rad51 expression via the MKK1/2–ERK1/2 signaling pathway in MMC-treated A549 and H1975 cells was required for cell survival. Consistent with our studies, reducing Rad51 expression by siRNA resulted in enhancement of MMC sensitivity in human lung cancer H1299 cells (Luoto et al., 2010). In contrast, a recent report has shown that knockdown of MYC, a basic helix–loop–helix leucine zipper transcription factor, using siRNA led to decreased Rad51 expression. However, an MYC-dependent decrease in Rad51 expression is not sufficient to sensitize cells to MMC in H1299 lung cancer cells (Luoto et al., 2010). Together, Rad51 must be carefully regulated to maintain genomic stability in cells responding to various DNA damage.
Previous studies have indicated that HR-defective cells are sensitive to several DNA-damaging agents, for example, MMC, cisplatinum, gemcitabine, ionizing radiation (IR), poly(ADP-ribose) polymerase inhibitors, thus suggesting the DNA repair protein participates in HR may be a target for overcoming chemoresistance in cancer therapy (Ashworth, 2008; Bryant et al., 2005; Helleday et al., 2007). Our study is thefirst to demonstrate that curcumin can decrease cellular and induced Rad51 expression, which results in enhancement of MMC-induced cytotoxicity. Curcumin, extracted from the rhizomes of the plant C. longa L., is the major component in curry dishes. Curcumin consumption at up to 100 mg/day in certain countries (Ammon and Wahl, 1991) has testified to confirm its pharmacological safety (Cheng et al., 2001). Curcumin has been reported to have numerous therapeutic benefits, including chemoprevention in cancer, and anti-inflammatory, antioxidant, and antiviral activities (Cheng et al., 2001; Levi et al., 2001; Rao et al., 1995; Ruby et al., 1995). In addition, curcumin enhances the chemotherapeutic effect of vinorelbine, which mediates apoptosis in human squamous cell lung carcinoma cells through the mitochondrial pathway (Sen et al., 2005). Curcumin also suppresses the paclitaxel-induced expression of antiapoptotic (XIAP, IAP-1, IAP-2, 2, and Bcl-xL), and proliferative (cyclooxygenase 2, c-Myc, and cyclin D1) proteins in breast cancer cells (Aggarwal et al., 2005). This study showed that combined treatment with curcumin and MMC could augment the cytotoxic effects of MMC against NSCLC cells. However, curcumin alone did not significantly affect on cell viability in human fibroblasts HFW and embryonic kidney HEK293 cells (Tsai et al., 2011). Consistent with our studies, Zhou et al. reported that combined treatment with MMC and curcumin reduced breast cancer tumor weight and inhibited tumor growth. Moreover, curcumin reduce the side effects of MMC by inhibiting the glucose regulatory protein (GRP58)-mediated DNA cross-linking through the ERK/p38 MAPK pathway in human breast cancer MCF-7 cells (Zhou et al., 2009). Additionally, our results are the first to provide an additional mechanism through which curcumin may mediate its antiproliferative effects, that is the downregulation of Rad51 expression in NSCLC cells.
In conclusion, we found that curcumin significantly suppressed MMC-induced cell proliferation and Rad51 expression in cultured NSCLC cells through inhibiting the MKK1/2–ERK1/2 pathway. These findings may aid in the understanding of the mechanisms of the synergistic cytotoxic effect of curcumin and MMC and may provide a research basis for continued biomedical study of this compound in appropriate models of human NSCLC. Although our in vitro study is supporting for study of curcumin as a modulating agent for MMC, there are many factors in consideration of human safety that help determine the dose of MMC for oncology treatments including, the type of cancer being treated, the method, patient health conditions, tolerance of side effects, and many others.
Acknowledgments
We thank Dr. Show-Mei Chuang and Dr. Jia-Ling Yang for providing us with expression plasmids for transfection. This work
was supported by the National Science Council of Taiwan, Grant no. NSC 99-2320-B-415-001-MY3 (to Y-W Lin) and Hsinchu Hospital. References
Aggarwal, B.B., Kumar, A., Bharti, A.C., 2003. Anticancer potential of curcumin: preclinical and clinical studies. Anticancer Res. 23, 363–398.
Aggarwal, B.B., Shishodia, S., Takada, Y., Banerjee, S., Newman, R.A., Bueso-Ramos, C.E., Price, J.E., 2005. Curcumin suppresses the paclitaxel-induced nuclear factor-kappaB pathway in breast cancer cells and inhibits lung metastasis of human breast cancer in nude mice. Clin. Cancer Res. 11, 7490–7498.
Ammon, H.P., Wahl, M.A., 1991. Pharmacology of Curcuma longa. Planta Med. 57, 1–7. Arnaudeau, C., Helleday, T., Jenssen, D., 1999. The RAD51 protein supports homologous recombination by an exchange mechanism in mammalian cells. J. Mol. Biol. 289, 1231–1238.
Ashworth, A., 2008. A synthetic lethal therapeutic approach: poly(ADP) ribose polymerase inhibitors for the treatment of cancers deficient in DNA double-strand break repair. J. Clin. Oncol. 26, 3785–3790.
Babiak, A., Hetzel, J., Godde, F., Konig, H.H., Pietsch, M., Hetzel, M., 2007. Mitomycin C and Vinorelbine for second-line chemotherapy in NSCLC—a phase II trial. Br. J. Cancer 96, 1052–1056.
Bhattacharjee, A., Richards, W.G., Staunton, J., Li, C., Monti, S., Vasa, P., Ladd, C., Beheshti, J., Bueno, R., Gillette, M., Loda, M., Weber, G., Mark, E.J., Lander, E.S., Wong, W., Johnson, B.E., Golub, T.R., Sugarbaker, D.J., Meyerson, M., 2001. Classification of human lung carcinomas by mRNA expression profiling reveals distinct adenocar-cinoma subclasses. Proc. Natl Acad. Sci. USA 98, 13790–13795.
Booton, R., Lorigan, P., Anderson, H., Baka, S., Ashcroft, L., Nicolson, M., O'Brien, M., Dunlop, D., O'Byrne, K., Laurence, V., Snee, M., Dark, G., Thatcher, N., 2006. A phase III trial of docetaxel/carboplatin versus mitomycin C/ifosfamide/cisplatin (MIC) or mitomycin C/vinblastine/cisplatin (MVP) in patients with advanced non-small-cell lung cancer: a randomised multicentre trial of the British Thoracic Oncology Group (BTOG1). Ann. Oncol. 17, 1111–1119.
Bristow, R.G., Hill, R.P., 2008. Hypoxia and metabolism. Hypoxia, DNA repair and genetic instability. Nat. Rev. Cancer 8, 180–192.
Bryant, H.E., Schultz, N., Thomas, H.D., Parker, K.M., Flower, D., Lopez, E., Kyle, S., Meuth, M., Curtin, N.J., Helleday, T., 2005. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature 434, 913–917.
Chan, M.M., Fong, D., Soprano, K.J., Holmes, W.F., Heverling, H., 2003. Inhibition of growth and sensitization to cisplatin-mediated killing of ovarian cancer cells by polyphenolic chemopreventive agents. J. Cell. Physiol. 194, 63–70.
Chan, N., Koritzinsky, M., Zhao, H., Bindra, R., Glazer, P.M., Powell, S., Belmaaza, A., Wouters, B., Bristow, R.G., 2008. Chronic hypoxia decreases synthesis of homologous recombination proteins to offset chemoresistance and radioresistance. Cancer Res. 68, 605–614.
Chen, Y.R., Tan, T.H., 1998. Inhibition of the c-Jun N-terminal kinase (JNK) signaling pathway by curcumin. Oncogene 17, 173–178.
Chen, L., Tian, G., Shao, C., Cobos, E., Gao, W., 2010. Curcumin modulates eukaryotic initiation factors in human lung adenocarcinoma epithelial cells. Mol. Biol. Rep. 37, 3105–3110. Cheng, A.L., Hsu, C.H., Lin, J.K., Hsu, M.M., Ho, Y.F., Shen, T.S., Ko, J.Y., Lin, J.T., Lin, B.R.,
Ming-Shiang, W., Yu, H.S., Jee, S.H., Chen, G.S., Chen, T.M., Chen, C.A., Lai, M.K., Pu, Y.S., Pan, M.H., Wang, Y.J., Tsai, C.C., Hsieh, C.Y., 2001. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer. Res. 21, 2895–2900.
Collett, G.P., Campbell, F.C., 2004. Curcumin induces c-jun N-terminal kinase-dependent apoptosis in HCT116 human colon cancer cells. Carcinogenesis 25, 2183–2189.
Daboussi, F., Dumay, A., Delacote, F., Lopez, B.S., 2002. DNA double-strand break repair signalling: the case of RAD51 post-translational regulation. Cell. Signal. 14, 969–975.
Dronkert, M.L., Kanaar, R., 2001. Repair of DNA interstrand cross-links. Mutat. Res. 486, 217–247.
Du, B., Jiang, L., Xia, Q., Zhong, L., 2006. Synergistic inhibitory effects of curcumin and 5-fluorouracil on the growth of the human colon cancer cell line HT-29. Chemotherapy 52, 23–28.
Godthelp, B.C., Wiegant, W.W., van Duijn-Goedhart, A., Scharer, O.D., van Buul, P.P., Kanaar, R., Zdzienicka, M.Z., 2002. Mammalian Rad51C contributes to DNA cross-link resistance, sister chromatid cohesion and genomic stability. Nucleic Acids Res. 30, 2172–2182.
Han, S.S., Keum, Y.S., Seo, H.J., Surh, Y.J., 2002. Curcumin suppresses activation of NF-kappaB and AP-1 induced by phorbol ester in cultured human promyelocytic leukemia cells. J. Biochem. Mol. Biol. 35, 337–342.
Helleday, T., Lo, J., van Gent, D.C., Engelward, B.P., 2007. DNA double-strand break repair: from mechanistic understanding to cancer treatment. DNA Repair (Amst) 6, 923–935. Helleday, T., Petermann, E., Lundin, C., Hodgson, B., Sharma, R.A., 2008. DNA repair
pathways as targets for cancer therapy. Nat. Rev. Cancer 8, 193–204.
Klein, H.L., 2008. The consequences of Rad51 overexpression for normal and tumor cells. DNA Repair (Amst) 7, 686–693.
Ko, J.C., Ciou, S.C., Cheng, C.M., Wang, L.H., Hong, J.H., Jheng, M.Y., Ling, S.T., Lin, Y.W., 2008. Involvement of Rad51 in cytotoxicity induced by epidermal growth factor receptor inhibitor (gefitinib, IressaR) and chemotherapeutic agents in human lung cancer cells. Carcinogenesis 29, 1448–1458.
Ko, J.C., Ciou, S.C., Jhan, J.Y., Cheng, C.M., Su, Y.J., Chuang, S.M., Lin, S.T., Chang, C.C., Lin, Y.W., 2009. Roles of MKK1/2–ERK1/2 and phosphoinositide 3-kinase-AKT signaling pathways in erlotinib-induced Rad51 suppression and cytotoxicity in human non-small cell lung cancer cells. Mol. Cancer Res. 7, 1378–1389.
337 J.-C. Ko et al. / Toxicology and Applied Pharmacology 255 (2011) 327–338
Landis, S.H., Murray, T., Bolden, S., Wingo, P.A., 1999. Cancer statistics, 1999. CA Cancer J. Clin. 49 (8–31), 31.
Levi, M.S., Borne, R.F., Williamson, J.S., 2001. A review of cancer chemopreventive agents. Curr. Med. Chem. 8, 1349–1362.
Luoto, K.R., Meng, A.X., Wasylishen, A.R., Zhao, H., Coackley, C.L., Penn, L.Z., Bristow, R.G., 2010. Tumor cell kill by c-MYC depletion: role of MYC-regulated genes that control DNA double-strand break repair. Cancer Res. 70, 8748–8759. McHugh, P.J., Spanswick, V.J., Hartley, J.A., 2001. Repair of DNA interstrand crosslinks:
molecular mechanisms and clinical relevance. Lancet Oncol. 2, 483–490. Mladenov, E., Anachkova, B., Tsaneva, I., 2006. Sub-nuclear localization of Rad51 in
response to DNA damage. Genes Cells 11, 513–524.
Mladenov, E., Tsaneva, I., Anachkova, B., 2007. Activation of the S phase DNA damage checkpoint by mitomycin C. J. Cell. Physiol. 211, 468–476.
Pfister, D.G., Johnson, D.H., Azzoli, C.G., Sause, W., Smith, T.J., Baker Jr., S., Olak, J., Stover, D., Strawn, J.R., Turrisi, A.T., Somerfield, M.R., 2004. American Society of Clinical Oncology treatment of unresectable non-small-cell lung cancer guideline: update 2003. J. Clin. Oncol. 22, 330–353.
Qiao, G.B., Wu, Y.L., Yang, X.N., Zhong, W.Z., Xie, D., Guan, X.Y., Fischer, D., Kolberg, H.C., Kruger, S., Stuerzbecher, H.W., 2005. High-level expression of Rad51 is an independent prognostic marker of survival in non-small-cell lung cancer patients. Br. J. Cancer 93, 137–143.
Radhakrishna Pillai, G., Srivastava, A.S., Hassanein, T.I., Chauhan, D.P., Carrier, E., 2004. Induction of apoptosis in human lung cancer cells by curcumin. Cancer Lett. 208, 163–170.
Rao, C.V., Rivenson, A., Simi, B., Reddy, B.S., 1995. Chemoprevention of colon carcinogenesis by dietary curcumin, a naturally occurring plant phenolic compound. Cancer Res. 55, 259–266.
Richardson, C., 2005. RAD51, genomic stability, and tumorigenesis. Cancer Lett. 218, 127–139.
Richardson, C., Stark, J.M., Ommundsen, M., Jasin, M., 2004. Rad51 overexpression promotes alternative double-strand break repair pathways and genome instability. Oncogene 23, 546–553.
Ruby, A.J., Kuttan, G., Babu, K.D., Rajasekharan, K.N., Kuttan, R., 1995. Anti-tumour and antioxidant activity of natural curcuminoids. Cancer Lett. 94, 79–83.
San Filippo, J., Sung, P., Klein, H., 2008. Mechanism of eukaryotic homologous recombination. Annu. Rev. Biochem. 77, 229–257.
Schiller, J.H., Harrington, D., Belani, C.P., Langer, C., Sandler, A., Krook, J., Zhu, J., Johnson, D.H., 2002. Comparison of four chemotherapy regimens for advanced non-small-cell lung cancer. N Engl J. Med. 346, 92–98.
Sen, S., Sharma, H., Singh, N., 2005. Curcumin enhances Vinorelbine mediated apoptosis in NSCLC cells by the mitochondrial pathway. Biochem. Biophys. Res. Commun. 331, 1245–1252.
Singh, S., Aggarwal, B.B., 1995. Activation of transcription factor NF-kappa B is suppressed by curcumin (diferuloylmethane) [corrected]. J. Biol. Chem. 270, 24995–25000.
Takenaka, T., Yoshino, I., Kouso, H., Ohba, T., Yohena, T., Osoegawa, A., Shoji, F., Maehara, Y., 2007. Combined evaluation of Rad51 and ERCC1 expressions for sensitivity to platinum agents in non-small cell lung cancer. Int. J. Cancer 121, 895–900.
Tsai, M.S., Kuo, Y.H., Chiu, Y.F., Su, Y.C., Lin, Y.W., 2010. Down-regulation of Rad51 expression overcomes drug resistance to gemcitabine in human non-small-cell lung cancer cells. J. Pharmacol. Exp. Ther. 335, 830–840.
Tsai, M.S., Weng, S.H., Kuo, Y.H., Chiu, Y.F., Lin, Y.W., 2011. Synergistic effect of curcumin and cisplatin via down-regulation of thymidine phosphorylase and excision repair cross-complementary 1 (ERCC1). Mol. Pharmacol. 80, 136–146.
Vispe, S., Cazaux, C., Lesca, C., Defais, M., 1998. Overexpression of Rad51 protein stimulates homologous recombination and increases resistance of mammalian cells to ionizing radiation. Nucleic Acids Res. 26, 2859–2864.
Warren, A.J., Hamilton, J.W., 1996. Synthesis and structural characterization of the N2G-mitomycin C-N2G interstrand cross-link in a model synthetic 23 base pair oligonucleotide DNA duplex. Chem. Res. Toxicol. 9, 1063–1071.
Weir, N.M., Selvendiran, K., Kutala, V.K., Tong, L., Vishwanath, S., Rajaram, M., Tridandapani, S., Anant, S., Kuppusamy, P., 2007. Curcumin induces G2/M arrest and apoptosis in cisplatin-resistant human ovarian cancer cells by modulating Akt and p38 MAPK. Cancer Biol. Ther. 6, 178–184.
Zhou, Q.M., Zhang, H., Lu, Y.Y., Wang, X.F., Su, S.B., 2009. Curcumin reduced the side effects of mitomycin C by inhibiting GRP58-mediated DNA cross-linking in MCF-7 breast cancer xenografts. Cancer Sci. 100, 2040–2045.