行政院國家科學委員會專題研究計畫 成果報告
非光合作用被子植物質體基因研究
計畫類別: 個別型計畫
計畫編號: NSC92-2311-B-002-026-
執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日
執行單位: 國立臺灣大學生態學與演化生物學研究所
計畫主持人: 胡哲明
計畫參與人員: 徐馨怡,呂佳穎
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 93 年 11 月 3 日
行政院國家科學委員會專題研究計畫成果報告
非光合作用被子植物質體基因研究
Studies of plastid genes in non-photosynthetic plants
計畫編號:NSC 92-2311-B-002-026
執行期間:
92年08月01日至93年07月31日
計畫主持人:胡哲明 國立台灣大學生態學與演化生物學研究所
計畫參與人員:徐馨怡,呂佳穎 國立台灣大學生態學與演化生物學
研究所
一、中文摘要
葉綠體存在於所有綠色植物體內,為 進行光合作用的場所。然而有些被子植物 喪失光合作用的能力,成為全寄生或腐生 的植物。這些非光合作用植物卻仍然保有 一個縮減的質體基因組,其中大部分與光 合作用相關的基因,如rbcL等,都已丟失 或成為偽基因(pseudogene) 。本計畫即以 九種台灣的非綠色被子植物為材料,進行 其質體基因的分析,探討質體基因的演 化。結果共獲得九條18S rDNA、七條16S rDNA序 列 , 並 依 據 其 序 列 進 行 譜 系 分 析。另外穗花蛇菰、無根藤和列當均利用 PCR得到接近全長的rbcL與matK序列。但 是在菟絲子中只得到rbcL,而野菰只能得 到matK的序列。而RT-PCR的結果顯示菟 絲子的rbcL有微量表現,應具有些微光合 作用能力。關鍵詞:
非光合作用植物,質體基因組, 18S rDNA,16S rDNA,rbcL,matKAbstract
Chloroplasts are the eukaryotic organelles mainly responsible for photosynthesis. There are some flowering plants, however, lost the photosynthetic
ability and became holoparasitic or saprophytic plants. The non-photosynthetic plants usually retain a reduced plastid genome, although many of plastid genes are lost or become pseudogenes, especially the photosynthesis-related genes such as rbcL. In order to elucidate the plastid genome evolution in non-photosynthetic plants, we examined nine non-photosynthetic plants native to Taiwan. We obtained all 18S rDNA sequences and seven 16S rDNA sequences of the examined taxa, and further using the sequences to perform phylogenetic analyses. In Balanophora,
Cassytha and Orobanche, we obtained
almost full length rbcL and matK sequences. However, we only obtained rbcL from
Cuscuta and matK sequence from Aeginetia.
The RT-PCR result shows that the rbcL transcripts were detected in Cuscuta, and suggests Cuscuta may have retain photosynthetic ability.
Keywords:
non-photosynthetic plants, plastid genome, 18S rDNA, 16S rDNA,二、緣由與目的
葉綠體為植物體內負責行光合作用 的胞器,衍生自藍綠菌的內共生。不過演 化至今,葉綠體的基因組大小只剩藍綠菌 的5~10%,約為150~200kb,縮減了20~30 倍之多(Martin et al. 2002)。葉綠體基因組 成環狀構造,主要可分為三類:(1)與轉 錄、轉譯有關的基因,(2)與光合作用有關 的基因,以及(3)其他一些與脂肪酸、色素 等生合成相關的基因(Lohan et al. 1998)。 然而在全寄生或腐生植物,這些喪失 光合作用能力的被子植物中,只存在退化 的葉綠體(dePamphilis et al. 1990, Nickrentet al. 1997),也保有一個更為縮減的質體 基因組(20~88kb)。這些非光合作用植物質 體中與光合作用相關的基因,大部分已丟 失或成為偽基因(pseudogene),如rbcL等 (Wolfe et al. 1992)。因此非光合作用植物 實為研究質體演化的最佳天然材料,透過 研究其質體,可進一步了解質體在植物細 胞中所扮演的角色,以及基因在無演化選 汰壓力下的行為。 目前已有一些相關研究針對非光合 作用植物的rbcL進行分析(van der Kooij et al. 2000),本計畫即以九種台灣的非綠色 被子植物為材料:Mitrastemon kanehirai ( 菱 形 奴 草 , 大 花 草 科 ) , Balanophora laxiflora (穗花蛇菰 ,蛇菰科),Cuscuta australes ( 菟 絲 子 , 旋 花 科 ) , Aeginetia indica ( 野 菰 , 列 當 科 ) , Orobanche coerulescens ( 列當 ,列 當科) ,Cassytha filiformi ( 無 根 藤 , 樟 科 ) , Cheilotheca humilis (水晶蘭,鹿蹄草科),Cheilotheca macrocarpa (阿里山水晶蘭,鹿蹄草科)以 及Galeola lindleyana (山珊瑚,蘭科) 。利 用PCR及點狀雜合法來分析這些非光合 作用植物的細胞核基因18S rDNA以及質 體基因16S DNA、rbcL和matK,也進一步 將序列進行譜系分析以及藉由RT-PCR檢 測 三 種 非 光 合 作 用 植 物rbcL 基 因 的 表 現。進而探討光合作用基因在非光合作用 植物中的演化。
三、結果與討論
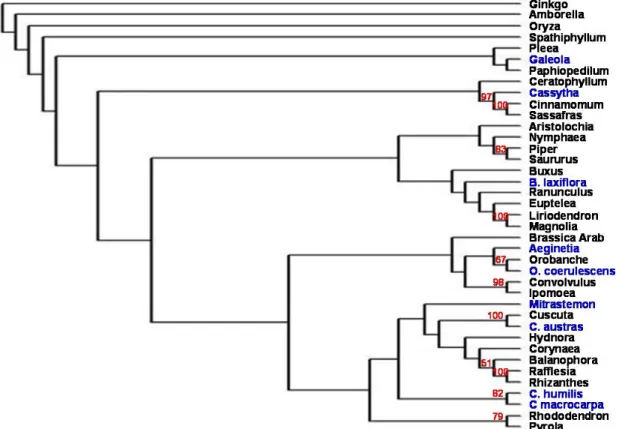
細胞核基因 18S rDNA 所有選取的非光合作用植物皆能利 用通用的引子增幅其18S rDNA片段,經 過定序後,其序列加上其他由GenBank下 載的被子植物18S rDNA序列以ClustalX 進行排序分析,再利用PAUP*4.0進行譜 系分析,得到譜系樹如Fig. 1。除菱形奴 草、穗花蛇菰外,其餘七種植物皆與相近 物種聚集在一起;而整個18S rDNA譜系 樹中的親緣關係也大致與其他研究符合 (Nickrent et al. 2000) 。 不 過 譜 系 樹 的 bootstrap值不高,而菱形奴草及穗花蛇菰 未與相近物種群聚一起,這些則尚待進一 步的譜系分析來加以釐清。 質體基因 16S rDNA 九種植物中,菱形奴草及穗花蛇菰無 法利用通用的引子獲得16S rDNA片段, 顯示其16S rDNA具有較大的變異。其他 七種非光合作用植物以及由GenBank下載 的被子植物16S rDNA序列經過譜系分析 後,所得譜系樹如Fig. 2。所有非光合作 用被子植物在譜系樹中的位置皆靠近其 親緣關係相近物種,而16S rDNA譜系樹 也與Nickrent所建構的相仿(Nickrent et al. 2000)。 rbcL與matK 在選取的植物中,穗花蛇菰、無根藤 及列當均能用通用引子獲得接近全長的 rbcL與matK序列。菟絲子和野菰則分別Fig. 1 利用Neighbor-joining分析方法所建構的18S rDNA譜系樹。藍色字體為本研究所選取的九
種非光合作用被子植物,紅色數字則為bootstrap值。
Fig. 2 利用Neighbor-joining分析方法所建構的16S rDNA譜系樹。藍色字體為本研究所選取的非
只能得到rbcL及matK序列;而菱形奴草、 兩種水晶蘭以及山珊瑚皆無法成功的增 幅rbcL與matK序列。進一步利用點狀雜合 法分析(Fig. 3),結果顯示所有的非光合作 用植物均具有rbcL與matK,表示這九種植 物皆存有質體基因組,而無法利用PCR增 幅者,其基因可能已產生相當的變異,但 尚未丟失。 rbcL基因的表現 為了進一步了解光合作用相關基因 的表現,我們選取了菱形奴草、穗花蛇菰 和菟絲子,利用RT-PCR檢測其rbcL的表 現(Fig. 4)。雖然經由PCR以及點狀雜合分 析結果得知菱形奴草與穗花蛇菰皆具有 rbcL基因,但由RT-PCR結果可看出,兩 者的RT-PCR產物均小於正常大小,其基 因序列應已經產生相當的變異。因此菱形 奴草及穗花蛇菰的rbcL雖可轉錄成 mRNA,但是否真正具有功能,仍需進一 步的實驗來加以確定。反之,菟絲子 Fig. 3 九種非光合作用植物之matK與rbcL基 因點狀雜合分析結果。 RT-PCR 的 結 果 看 出 具 有 正 常 大 小 的 產 物,顯示其rbcL基因有微量的表現;加上 菟絲子的植物體為黃綠色,由此推測菟絲 子可能具有些微光合作用能力,此結果與 van der Kooij 等 人 的 研 究 相 符 (van der Kooij et al. 2000)。 18S與16S rDNA建構出的譜系樹兩 者間並無太大的衝突之處,也與其他研究 相吻合;不過也未能釐清分類地位尚不清 楚的非光合作用植物物種,有待更進一步 的譜系分析。本研究結果大致上可對應到 植物體的形態,如四個基因均能增幅的無 根藤其幼株為明顯的綠色,列當植株則為 褐色。只有三個基因能增幅的菟絲子為黃 綠色,而穗花蛇菰與野菰植物體也具有明 顯的顏色。其餘菱形奴草,兩種水晶蘭以 及山珊瑚的植物體顏色由白至淡黃,均不 顯著;其質體基因的增幅結果也不甚理 想。其中,菱形奴草只成功增幅18S rDNA,推測其質體基因組很可能有極端 Fig. 4 利用RT-PCR進行菱形奴草、穗花蛇菰 及菟絲子rbcL基因之表現結果。
變異的質體基因組。不過上述推論,仍需 進一步進行其他質體基因的分析,以及檢 測光合作用相關基因的表現來加以證實。
四、計畫成果自評
透過本研究計畫的結果,我們能夠進 一步的了解質體基因組的演化,進而探討 質體在植物細胞中的角色,以及光合作用 相關基因在無選汰壓力下的演化行為;而 相關的譜系分析也有助於釐清這些非光 合作用植物的分類地位。除此之外,這些 全寄生或腐生被子植物的光合作用相關 基因從具有些微功能到已產生相當的變 異,可看出這些植物雖為全寄生或腐生, 但其質體具高度的變異,顯示這些植物的 多樣性。期能藉由更進一步的研究,讓我 們對非光合作用植物以及質體的演化有 更深入的了解。五、參考文獻
dePamphilis, C. W., and J. D. Palmer (1990) Loss of photosynthetic and
chlororespiratory genes from the plastid genomes of a parasitic flowering plant. Nature 348: 337-339.
Lohan, A. J., and K. H. Wolfe (1998) A subset of conserved tRNA genes in plastid DNA of nongreen plants. Genetics 150: 425-433.
Martin, W., T. Rujan, E. Richly, A. Hansen, S. Cornelsen, T. Lins, D. Leister, B. Stoebe, M. Hasegawa, and D. Penny (2002) Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of
cyanobacterial genes in the nucleus. Proc Natl Acad Sci USA 99: 12246-12251.
Nickrent, D. L., O. Y. Yan, R. J. Duff, and C. W. dePamphilis (1998) Do nonasterid holoparasitic flowering plants have plastid genomes? Plant Mol Biol 34: 717-729.
Nickrent, D. L., R. O. Duff, A. E. Colwell, A. D. Wolfe, N. D. Young, K. E. Steiner, and C. W. dePamphilis (2000) Molecular phylogenetic and evolutionary studies of parasitic plants. In Molecular systematics of plants II: DNA sequencing, eds. D. E. Soltis, P. S. Soltis, and J. J. Doyle, pp. 211-241. Kluwer Academic Publishers, Norwell.
van der Kooij, T. A. W., K. Krause, I. Dorr, and K. Krupinska (2000) Molecular, functional and ultrastructural characterisation of plastids from six species of the parasitic flowering plant genus Cuscuta. Planta 210: 701-707. Wolfe, K. H., C. W. Morden, and J. D.
Palmer (1992) Function and evolution of a minimal plastid genome from a
nonphotosynthetic parasitic plant. Proc Natl Acad Sci USA 89: 10648-10652.