Differentiation (2002) 70:447–456 C Blackwell Verlag 2002
O R I G I N A L A R T I C L E
Yau-Hung Chen¡ Huai-Jen Tsai
Treatment with Myf5-morpholino results in somite patterning and brain
formation defects in zebrafish
Accepted in revised form: 25 June 2002
Abstract Myf-5 is a stage-dependent transcription factor
associated with somitogenesis. To study its biological functions in zebrafish, we injected the Myf5-morphol-inos ZMF-MO (antisense nucleotides 28 to 52) and ZMF-OTHER (antisense nucleotides 3 to 27) into ze-brafish embryos to establish a myf-5 gene knockdown. No phenotypic abnormalities were observed following injection with 0.2 ng of ZMF-MO, but defects were dis-played in 2 of 118 (1.7 %) surviving embryos injected with 1 ng ZMF-MO. Morphological defects became more severe with increased dosages: 105 of 270 (38.9 %) surviving embryos injected with 4.5 ng of ZMF-MO dis-played such abnormalities as the absence of eyes or brains in addition to the following low-dosage defects in 24 hpf embryos: longitudinal yolk sacs, incomplete epib-oly coverage, abnormal and suspended tail buds, dif-fused somite boundaries, and head shrinkage. Similar results were observed in the 4.5 ng ZMF-OTHER injec-tion group. However, when fish were co-injected with 4.5 ng ZMF-MO and 4.5 ng myf-5 mRNA, abnormality rates decreased from 49.6 % to 5.5 %. Our results show that the brain krox20 gene was down-regulated at rhom-bomere 3; the pax2.1 gene was completely down-regu-lated; myoD was expressed normally; myogenin was sub-stantially down-regulated in whole somites; and desmin was partly inhibited in newly forming somites. Our con-clusion is that zebrafish Myf-5 may play important roles in brain formation and in the convergence and extension of shield epiblasts and tail buds during early embryogen-esis, in addition to its well-understood role as a muscle regulatory factor in somites.
Y.-H. Chen¡ H.-J. Tsai (
✉
)Institute of Fisheries Science, National Taiwan University, 1 Roosevelt Road, Sec. 4, Taipei, Taiwan
Tel:π886 2 2366 1540, Fax: π886 2 2363 8483 e-mail: hjtsai/ccms.ntu.edu.tw
U. S. Copyright Clearance Center Code Statement:0301–4681/2002/7008–447 $ 15.00/0
Key words brain¡ embryogenesis ¡ fish ¡ morphant ¡ morpholino¡ myf-5 ¡ somite
Introduction
In eukaryotic cells, muscle-specific gene transcription is controlled by a series of complex interactions among DNA-binding transcription factors and specific DNA recognition sequences. These factors can be categorized in terms of shared sequences and structural motifs, in-cluding the basic helix-loop-helix (bHLH) motif. Myog-enesis involves several bHLH proteins, such as MyoD, MYF-5, Myogenin, and MRF4. The moleculars of my-ogenic regulatory factors (MRFs) have been identified in mammals (Buonanno et al., 1992), birds (Saitoh et al., 1993), frogs (Hopwood et al., 1991), fish (Chen et al., 2000; 2002), insects (Michelson et al., 1990) and nematodes (Krause et al., 1994).
In mice, MyoD and Myf-5 play distinct roles in myog-enesis. Cells in the epaxial myotome usually express
myoD, while hypaxial myotomes express myf-5
(Rud-nicki and Jaenisch, 1995). Epaxial and hypaxial myo-tomes eventually differentiate into limbs and trunk muscles, respectively. Braun et al. (1992) reported that the targeted inactivation of myf-5 results in embryonic death; the mice embryos they studied were unable to breathe owing to the absence of ribs and sternum, even though normal muscle development was observed. In fish, the biological functions of Myf-5 have yet to be fully characterized. One exception is Neyt et al.’s (2000) use of zebrafish myoD as a molecular marker to demon-strate that teleost fin muscles in tetrapod species develop from migratory mesenchymal precursor cells that pos-sess molecular and morphogenetic similarities with limb muscle precursors. However, no reports have been pub-lished to date on the targeted inactivation or gene knockdown of fish myf-5.
448
Recently, Chen et al. (2001) and Coutelle et al. (2001) reported that zebrafish myf-5 transcripts were detectable at 7.5 hours post-fertilization (hpf), increased substan-tially until 16 hpf, then gradually declined to undetect-able levels by 26 hpf. They also found that during somi-togenesis, zebrafish myf-5 transcripts were primarily found in somites and segmental plates. Prominent sig-nals occurred transiently in adaxial cells in two parallel rows, but did not extend beyond positive-signal somites. Since adaxial cells and presomitic mesoderms are pre-cursors of slow and fast muscles, respectively (Devoto et al., 1996), we believe that zebrafish myf-5 may be in-volved in slow and fast muscle formation.
Morpholino, a nucleotide analogue, has been de-scribed by Nasevicus and Ekker (2000) as a powerful tool for gene function knockdown in zebrafish. To deter-mine the biological functions of zebrafish myf-5, we in-jected Myf5-morpholino (myf5-MO) into single-cell ze-brafish embryos and recorded the resulting myf-5 knock-down phenotypes. From our observations of zebrafish
myf-5 morphants, we found that myf-5 is not only
in-volved in myogenesis, but also plays important roles dur-ing early embryogenesis in the convergence and exten-sion of shield epiblasts, as well as in tail bud and brain formation.
Methods
FishAB strain Zebrafish were kept at a photoperiod of 14 h light/10 h dark at approximately 28.5æC. After fertilization, eggs were col-lected and cultured in an aquarium. Embryonic cleavage numbers were counted and somite formation observed with a light micro-scope to determine developmental stage (Kimmel et al., 1995).
Preparation and microinjection of morpholino
Zebrafish myf5-morpholino (ZMF-MO: 5ø-TACGTCCATGAT-TGGTTTGGTGTTG-3ø) and myf5-morpholino-other (ZMF-OTHER: 5ø-AAGGTTTCTGAGTGGTGTGGTTGCA-3ø) were established based on zebrafish myf-5 cDNA complementary se-quences from nucleotides (nt) 28 to 52 and 3 to 27, respectively (Gene Tools). Fluoresceinated chordin-MO (5ø-ATCCA-CAGCAGCCCTCCATCATCC-3ø) and standard control (non-specific) MO (STD-MO) (5ø-CCTCTTACCTCAGTTA-CAATTTATA-3ø) were used as positive and negative controls, re-spectively (Gene Tools). All of the above were prepared at stocking concentrations of 1 mM and diluted with double-distilled water to concentrations of 10, 53, 133 and 250mM (4.5 ng/embryo).
For our rescue experiment, we established the primers ZMFC1F (5ø-CCCAGTGCAACCACACCAC-3ø) and ZMFC1433R (5ø-CC-CGTTATTAAATAACACATTT-3ø) to amplify full-length ze-brafish myf-5 cDNA by reverse transcription-polymerase chain re-action, using Superscript II (Gibco) and ExTaq DNA polymerase (TAKARA) as described in Chen et al. (2001). The resulting PCR product was ligated to pGEMT-easy vector (Promega); the myf-5 cDNA in pT7-ZMF was driven by the T7 promoter. To construct pT7-EGFP, in which EGFP cDNA was also driven by the T7 pro-moter, primers GFP-F (5ø-ATGGTGAGCAAGGGCGAGGA-3ø) and GFP-R (5ø-CAGCTCGTCCATGCCATGTG-3ø) were
syn-thesized and processed for PCR. For myf-5 or GFP mRNA prep-aration, approximately 5mg of SpeI-digested ZMF or pT7-EGFP and mMESSAGE mMACHINETM (Ambion) were manu-factured according to the supplier’s directions. After synthesis, myf-5 and GFP mRNAs were diluted to working concentrations of 100, 500, 750 and 2000 ng/ml prior to each 2.3 nl injection.
Digoxigenin (DIG)-labeled riboprobe preparation
Approximately 5mg each of plasmids containing cDNA of desmin (nt 1 to 1772) (Loh et al., 2000; Xu et al., 2000), myoD (nt 216 to 951) (Weinberg et al., 1996), myogenin (nt 114 to 833) (Chen et al., 2000), pax2.1 (nt 1214 to 1854) (Pfeffer et al., 1998), and krox20 (nt 463 to 1021) (Oxtoby and Jowett, 1993) – all driven by a T7 promoter – were used as templates to prepare riboprobes (DIG RNA Labeling Kit, Rosche).
Whole-mount in situ hybridization
Normal and morphant embryos were collected at 24 hpf, fixed with 4 % paraformaldehyde for 4 h at 25æC, dechrionized, dehydrated with an ascending methanol series (25 %, 50 %, 75 % and 100 %), and stored in methanol atª 20 æC. The in situ hybridization of whole embryos was performed using DIG-labeled riboprobes of desmin, myoD, myogenin, pax2.1, and krox20 according to pro-cedures described in Chen et al. (2001), with some modifications. Embryos were permeabilized with acetone atª 20 æC, digested with 10mg/ml proteinase K for 15 min, and prehybridized for at least 4 h. Hybridization took place at 70æC in the presence of approxi-mately 10 ng DIG-labeled riboprobes for a minimum of 16 h. After washing, hybridized embryos were incubated overnight in a 1:10,000 dilution of alkaline phosphatase-conjugated anti-DIG antibody (Rosche) at 4æC. Excess antibodies were removed by pro-gressively longer washings with 0.1 % Tween-20 in phosphate-buf-fered saline (10, 10, 10, 20, 30, and 60 min). Color reactions (using nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate as substrates) were used to detect the hybridized myoD, desmin, myogenin, pax2.1, and krox20 mRNA transcripts.
Embedding and cryosectioning
Morphants and wild-type embryos were fixed with 4 % paraform-aldehyde for 4 h at 25æC, dechrionized, mounted with 5 % sucrose containing 1.5 % agarose for 1 h, cut into cubes approximately 5¿ 5¿ 5mm and stored in 5 % sucrose at 4 æC. All embryos were em-bedded using Cryomatrix (Shandon); cryosectioning (15mm thick) was performed with a Microm Cryosector (Heidelberg).
Microscopy
All wide-type, morphant, and stained embryos were examined with a DMR microscope and photographed with a MPS 60 camera (both from Leica) using FUJI X-TRA 400 ASA film.
Results
Defective morphology in Myf5-MO-injected embryos First, we injected a chordin antisense morpholino (chordin-MO) conjugated with fluorescein into zebrafish embryos as a positive control to observe specific pheno-types. Embryos injected with 4.5 ng of chordin-MO re-sulted in phenotypes marked by small heads, extremely
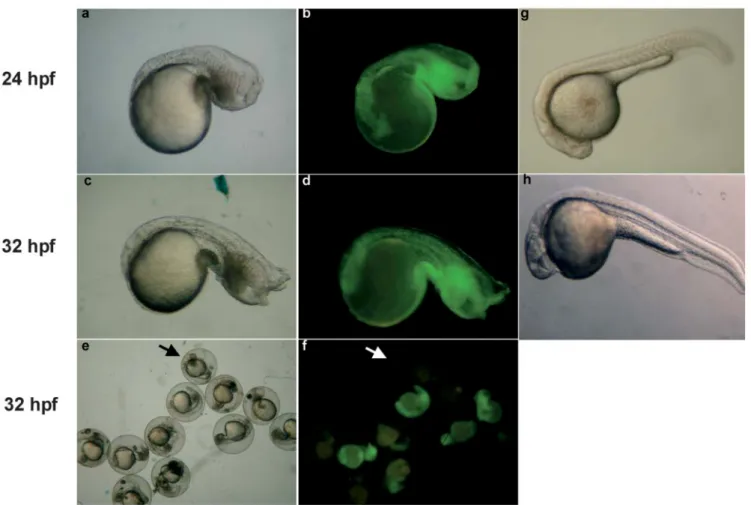
Fig. 1 Observation of chordin-morphants. Chordin-morphants de- were also illustrated. Arrows indicate that no green signals were veloped at 24 (a, b) and 32 hpf (c–f) were observed and photo- detected in chordin-MO-injection group (f), resulting in normal graphed under bright field illumination (a, c and e) and blue light phenotype (e).
(b, d and f). Wild-type embryos developed at 24 (g) and 32 hpf (h)
large blood islands, abnormal tail fins, and U-shaped somites (Figs. 1a and 1c). They were similar to the chordin null-mutant embryos described by Ham-merschmidt et al. (1996), Fisher et al. (1997), and Nasev-icus and Ekker (2000). Green signals were equally dis-tributed, indicating chordin-MO stability in 24-hpf (Fig. 1b) and 32-hpf embryos (Fig. 1d).
Table 1 Dose response of phenotypes caused by ZMF-MO and ZMF-OTHER injection
Injection dose Mock ZMF-MO ZMF-OTHER STD-MO Chordin-MO
(ng/embryo) 0.2 1 2.5 4.5 4.5 4.5 4.5 Injection embryos 334 178 193 142 501 466 166 193 Survival Embryos 289 (86.5%) 150 (84.3%) 118 (61.1%) 96 (67.6%) 270 (53.9%) 266 (57.1%) 145 (87.3%) 146 (75.6%) Mild defects 0 0 2 (1.7%) 3 (3.1%) 42 (15.6%) 38 (14.3%) 0 N.D. Severe defects 0 0 9 (7.6%) 11 (11.5%) 105 (38.9%) 86 (32.3%) 0 97 (66.4%). The percentages of abnormal phenotypes shown in parentheses were calculated from survival embryos.
.
Severe defects in chordin-MO injection group including reduced head, extremely expanded blood island, abnormal tail fin and U-shaped somites.
No abnormal phenotypes were observed in negative control embryos injected with standard control/non-spe-cific MO (STD-MO); survival was measured at 87.3 % (Table 1). Likewise, no abnormal phenotypes were ob-served in mock control embryos injected with the same volume of double-distilled water, with an 86.5 % survival rate (Table 1). In the experimental group, all 150
sur-450
Fig. 2 Dynamic observation of ZMF-morphants. Wild-type em- coverage and suspended tail bud, respectively. Morphants were bryos (a–c, g) and ZMF-morphants (d–f, h–i) developed at 7.5 (a, classified into severe and mild defects based on morphological
ab-d), 10 (b, e), 11.5 (c, f) and 24 hpf (g–i) were observed with a DMR normalities described in Methods. microscope. Arrow and arrowhead indicate incomplete epiboly
viving embryos injected with 0.2 ng myf5-MO displayed normal embryonic development, but those injected with 1, 2.5 or 4.5 ng displayed abnormal phenotypes.
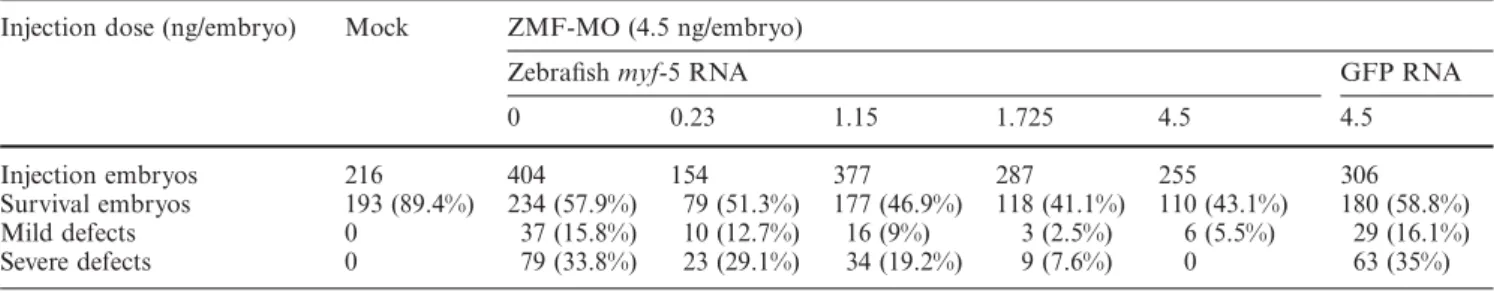
Table 2 Dose response of phenotypes rescued by coinjection of ZMF-MO and capped zebrafish myf-5 or GFP RNAs
Injection dose (ng/embryo) Mock ZMF-MO (4.5 ng/embryo)
Zebrafish myf-5 RNA GFP RNA
0 0.23 1.15 1.725 4.5 4.5
Injection embryos 216 404 154 377 287 255 306
Survival embryos 193 (89.4%) 234 (57.9%) 79 (51.3%) 177 (46.9%) 118 (41.1%) 110 (43.1%) 180 (58.8%) Mild defects 0 37 (15.8%) 10 (12.7%) 16 (9%) 3 (2.5%) 6 (5.5%) 29 (16.1%)
Severe defects 0 79 (33.8%) 23 (29.1%) 34 (19.2%) 9 (7.6%) 0 63 (35%)
The percentages of abnormal phenotypes shown in parentheses were calculated from survival embryos.
At 70 % epiboly (approximately 7.5 hpf), wide-type embryos displayed spherical shapes (Fig. 2a), while mor-phants displayed longitudinal shapes and swelling yolk
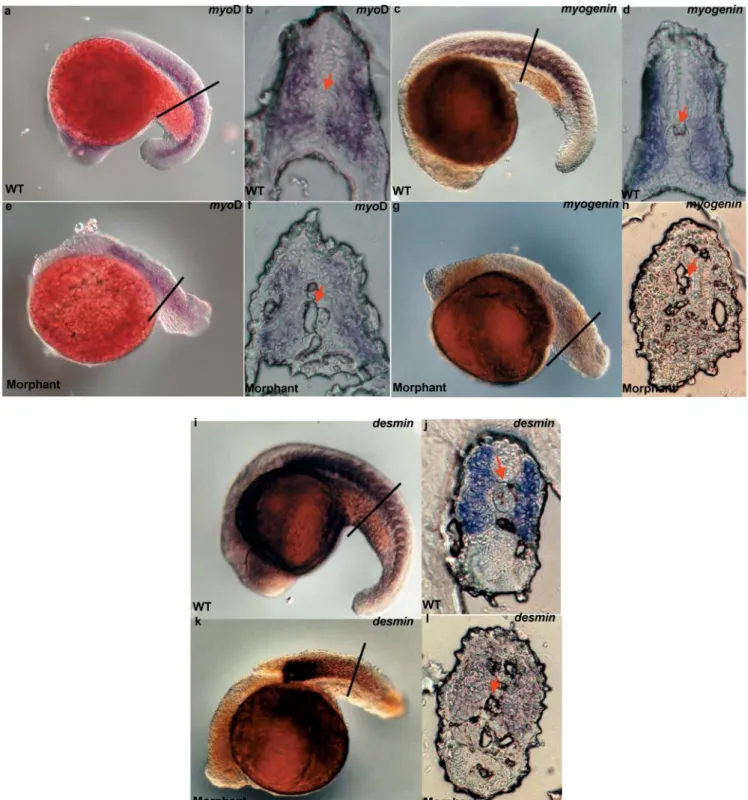
Fig. 3 Expression patterns of myoD, myogenin, and desmin tran- h, j and l) were performed using riboprobes as indicated. Lines in
scripts in wild-type embryos and ZMF-morphants. Whole-mount embryos indicate where cryosections were cut. Arrows indicate the in situ hybridization (a, c, e, g, i and k) and cryosectioning (b, d, f, position of notochord.
sacs (Fig. 2d). At the end of the epibolic stage (∂ 9 hpf), myf5-MO-injected embryos developed bubble-like tail bud vesicles (Fig. 2e), which eventually formed abnor-mal tail buds that failed to attach to the yolk at 11.5 hpf (Fig. 2f). Some embryos injected with higher dosages of
myf5-MO displayed abnormal somites at the middle seg-mental stage (18 – 24 hpf). Those abnormalities included morphologically indistinct somite boundaries, lack of uniformly sized cell blocks, unclear tail buds, and un-clear horizontal myosepta (Figs. 2h and 2i).
452
Fig. 4 Detection of brain markers, krox20 and pax2.1, in wild-type were detected at r5 (b, arrow) but not r3. In the mild-defect ZMF-and MO-treated embryos. Krox20 (a, b) ZMF-and pax2.1 (c, d) gene morphants, pax2.1 transcripts were detected at the optic vesicle (d, transcripts were detected in wild-type embryos (a, c) and ZMF- dotted-arrows) and MHB (arrowhead). MHB: midbrain-hindbrain morphants (b, d, e). In the ZMF-morphants, krox20 transcripts boundary; OTV: optic vesicle.
Abnormalities were categorized as mild or severe fol-lowing morphological observations of myf5-MO-injected embryos at the end of the segmental stage (∂ 24 hpf). Mild defects mostly consisted of abnormal tail bud somites (Fig. 2i), with a few scorpion-like tails also noted. Severe defects included the absence of eyes and abnormal heads (diminished telencephalons and mesencephalons) in ad-dition to the mild defects just described (Fig. 2h).
Dose-dependent Myf5-morphant abnormalities
Our results show that the frequency of phenotype abnor-malities was myf5-MO dose-dependent (Table 1). At 1 ng, 9.3 % (1.7 % mild, 7.6 % severe) of the surviving em-bryos displayed defects; at 2.5 ng, the percentage in-creased to 14.6 % (3.1 % mild, 11.5 % severe), and at 4.5 ng, the rate jumped to 54.5 % (15.6 % mild, 38.9 %
se-vere). In addition, a sharp decrease in survival rate (from approximately 60 % to 25 %) was noted when embryos were treated with 9 ng myf5-MO: 56.4 % (31/55) of the surviving embryos showed arrested growth during the epibolic stage (data not shown).
Similar efficacies for two Myf5-MO types
We injected the morpholino ZMF-OTHER into em-bryos to gather further proof that the defective morph-ant phenotype was the result of the endogenous myf-5 knockdown. Abnormality rates resulting from 4.myf-5 ng injections of ZMF-OTHER (14.3 % mild and 32.3 % se-vere) were similar to those caused by 4.5 ng injections of ZMF-MO (15.6 % mild and 38.9 % severe). The data suggest that the mild and severe defects were both the result of myf-5 knockdown phenotypes.
Co-injection of capped zebrafish myf-5 mRNA and ZMF-MO
We found that incidences of severe and mild defects were reduced when embryos were co-injected with various dosages of capped myf-5 RNA and 4.5 ng of ZMF-MO. In the low-dosage-injection group (0.23 ng/embryo), the abnormality rate decreased from 49.6 % (15.8 % mild and 33.8 % severe) to 41.8 % (12.7 % mild and 29.1 % se-vere); in the high-dosage-injection group (4.5 ng/em-bryo), the abnormality rate decreased significantly to 5.5 % (5.5 % mild and 0 % severe) (Table 2). However, severe and mild defects increased slightly in control group em-bryos co-injected with 4.5 ng of capped GFP RNA and ZMF-MO (52.1 % total, 16.1 % mild and 35 % severe) (Table 2). According to the data, the effect of ZMF-MO on knockdown myf-5 was both distinct and consistent.
Zebrafish Myf-5 and somitogenesis
To identify which muscle-specific genes were down-regu-lated in the 24-hpf ZMF morphants (resulting in somite-patterning defects), we performed whole-mount in situ hybridization with zebrafish myoD, myogenin, and
des-min riboprobes. Our results show that myoD transcripts
were strongly detectable in the somites and adaxial cells of wild-type embryos (Fig. 3a). Myogenin expression patterns were similar to those for myoD (Fig. 3c). In ZMF-morphants, myogenin transcripts were substan-tially down-regulated in the entire somite block (Fig. 3g), but myoD transcripts were not (Fig. 3e).
Our inspections of somite block cross-sections showed that myogenin transcripts appeared in the entire somite block, including slow- and fast-muscle pioneers in nor-mal embryos (Fig. 3d), but they were just barely detect-able in morphants (Fig. 3h). On the other hand, desmin transcripts were strongly detectable in whole somite blocks (anterior to posterior) as well as in slow- and fast-muscle pioneers, including adaxial cells (Figs. 3j and 3l). In the oldest 24-hpf ZMF-morphant somites, desmin transcripts were clearly detectable, although with vague contours (Fig. 3k). The only signs of desmin inhibition were noted in newly forming somites (Fig. 3k). Note that all sections of myf5-morphants displayed a shrink-age notochord (Figs. 3f, 3h and 3i), indicating that Myf-5 may be involved in notochord formation.
Zebrafish krox20 and pax2.1 transcripts
Our results show that ZMF-MO treatment resulted in severe brain defects in ZMF-MO morphants. We used the same procedure just described with forebrain (pax2.1) and hindbrain (krox20) riboprobes to identify which portions of the brain in zebrafish were affected by ZMF-MO. In normal embryos, pax2.1 transcripts were
detected in the forebrain, midbrain-hindbrain boundary (MHB), and optic stalk (Fig. 4c); krox20 transcripts were detected in rhombomeres 3 and 5 (r3 and r5) (Fig. 4a). The krox20 transcripts were expressed normally at the r5 primordium, but down-regulated at r3 (Fig. 4b). In contrast, pax2.1 transcripts were strongly detected in the optic primordium and MHB (Fig. 4c), but were sub-stantially down-regulated in the forebrains of mildly de-fective ZMF-morphants (Fig. 4d).
Discussion
RNA-interference (RNAi) has been described as being very strong in Caenorhabditis elegans (Fire et al., 1998) and Drosophila (Misquitta and Paterson, 1999); Zhao et al. (2001) reported an unacceptably high level of RNAi in zebrafish. The use of a morpholino is currently considered the best antisense approach for in vivo studies of knock-down gene function in zebrafish (Nasevicius and Ekker, 2000; Ekker and Larson, 2001) for known (Dutton et al., 2001) as well as novel genes (Scholpp and Brand, 2001).
Epiblast convergence and extension
Data from our whole-mount in situ hybridization show that endogenous zebrafish myf-5 transcripts were first de-tected at 70 % epiboly, reached maximum expression at 16 hpf, and down-regulated at the end of the segmentation stage. ZMF-morphant defects were also first noticed at 70 % epiboly (Fig. 2d); a RT-PCR analysis revealed that en-dogenous zebrafish myf-5 transcripts were weakly detect-able at 50 % epiboly (data not shown). Chen et al. (unpub-lished results) found that injection with a DNA construct containing zebrafish myf-5 upstreamª9977 to ª1 fused with a GFP reporter gene resulted in green signals becom-ing detectable in epiblasts at 50 % epiboly; by this point, epiblasts are undergoing convergence and extension, and cell lineages are becoming increasingly specific. Kiefer and Hauschka (2001) found that chicken myf-5 was ex-pressed in epiblasts at 50 % epiboly and were exex-pressed in some non-muscle mesodermic cells following whole-mount in situ hybridization. In the present study, half of the surviving embryos in the 9 ng-injected myf5-MO group showed arrested growth during the epibolic stage. In the case of total myf-5 knockdown, embryos were un-able to complete the epibolic stage and commence with segmentation. We therefore suggest that zebrafish Myf-5 has multiple functions, including epiblast migration and segmentation.
Myf-5 knockdown phenotypes
At 24 hpf, ZMF morphants displayed mild and severe defects. Both ZMF-MO (nt 28 to 52 of the antisense
454
strand) and ZMF-OTHER (nt 3 to 27 of the antisense strand) were capable of causing such defects in 24-hpf morphants. No significant differences were noted in sur-vival rates between embryos treated with either ZMF-MO (53.9 %) or ZMF-OTHER (57.1 %), but a slightly larger difference was noted in defective rates resulting from MO treatment (54.5 %) compared to ZMF-OTHER treatment (46.6 %); this may be due to different blocking translation efficiencies between the two. Defect reduction attributed to the co-injection of ZMF-MO and capped myf-5 RNA was dose-dependent (Table 2). Following injections of capped myf-5 RNA at dosages as high as 4.5 ng/embryo, morphant survival was meas-ured at 43.1 % with a defect rate of only 5.5 %. However, injections of capped GFP RNA were incapable of reduc-ing defective embryo rates (Table 2). These observations indicate that the mild and severe defects noted in this study were both distinct and consistent.
Comparing zebrafish ZMF-morphant and mice myf-5 knockout phenotypes
Zebrafish embryos treated with myf-5 morpholino showed developmental defects, including epiblast mi-gration retardment, abnormal somite patterning, brain deformation, and lethal effect of embryos. Severe rib de-fects and embryonic death were also noted in myf-5 knockout mice embryos, but defects in epiblast mi-gration, somite patterning, and brain formation have not been reported. The greater number of abnormalities found in zebrafish treated with myf-5 morpholino may be due to a) a more concentrated effort to identify ze-brafish morphants, and/or b) Myf-5 performs different biological functions in fish and mammalian systems.
Biological functions of Myf-5 during somitogenesis In this study, zebrafish morphants showed abnormal muscle and defective somite patterning, suggesting that Myf-5 is essential to muscle development. To identify which muscle-specific factors are regulated by Myf-5 during somitogenesis, we searched for myoD, myogenin, and desmin expression patterns in wild-type embryos and ZMF-morphants. Myf-5, MyoD, and Myogenin are all classified as basic helix-loop-helix transcription fac-tors, with MyoD being involved in myocyte determi-nation and Myogenin being involved in myotube forma-tion (Rudnicki and Jaenisch, 1995). Desmin is a muscle-specific intermediate filament protein that is regulated by Myf-5, MyoD, and Myogenin in mice (Li and Capet-anaki, 1993; Rudnicki et al., 1993; Li et al., 1994). We detected desmin transcripts in zebrafish in both the superficial muscle cells and the deeper parts of myo-tomes (Figs. 3i and 3j). These were co-localized with
myf-5, myoD, and myogenin, suggesting that desmin is
also regulated by MRFs. According to Braun et al. (1992) and Rudnicki et al. (1992), Myf-5 and MyoD compensate for each other in mice. Both teams of re-searchers also found normal myogenin and desmin tran-scription levels in myoD or myf-5 knockout mice. Hol-lenberg et al. (1993) demonstrated that myogenin is a direct transcriptional target of MyoD. Surprisingly, our results show that myoD transcripts were not affected in
myf-5 knockdown zebrafish, but that myogenin and des-min transcripts were down-regulated in all somites and
newly forming somites, respectively. Recently, Lindon et al. (2001) proposed that Myf-5 is a potent inducer of
myogenin at high cell densities; there is also evidence that
Myf-5 is fully capable of activating myogenin. We there-fore suggest that myogenin is a transcriptional target found downstream of both myf-5 and myoD.
ZMF-morphants display brain abnormalities
Another possible explanation for brain abnormalities re-sulting from myf-5 knockdown zebrafish is reduced skel-etal muscle. During nervous system development, motor neurons are believed to compete for specific skeletal muscle sites, with their long-term survival dependent upon receiving trophic support from the targeted muscle (Oppenheim, 1996). In addition, Kablar and Rudnicki (1999) report that motor neurons in mice normally ap-pear in the spinal cord, and that all somatic motor neu-rons in mutant mice are eliminated by apoptosis in
myf5ª/ª:myoDª/ª double knockout mice. Sporle et al. (1996) described brain defects and strongly altered myf-5 expression patterns in open brain (opb) mutant mice. Combined, this evidence suggests that nervous develop-ment is intimately coupled with skeletal myogenesis. Clearly, myf-5 mRNAs exist in the central nervous sys-tem (CNS), but their biological functions during neuron or brain development require further study.
Finally, we found that reduced krox20 and pax2.1 ex-pression domains in the anterior hindbrain region re-sulted in MHB and r3 hypoplasia. We therefore suggest that zebrafish Myf-5 may not be essential for hindbrain segmentation and specification, but may be essential for
krox20 activation in the anterior hindbrain region.
Myf-5 may be involved in neurogenesis
Most studies on Myf-5 focus on myogenesis. However, Tajbakhsh and Buckingham (1995) detected myf-5 tran-scripts in mice brains, and therefore suggested that myf-5 might play an important role in brain development. Furthermore, Daubas et al. (2000), Hadchouel et al. (2000), and Carvajal et al. (2001) detected myf-5 mRNA in the CNS of mice, and therefore suggested that CNS-specific cis-elements may occur in the upstream -8.8 kb. These reports clearly show the presence of myf-5
tran-scripts in neurons, but their biological roles during neurogenesis remain unclear.
In this study, we observed that brain development was impeded in myf5-MO-treated embryos. According to Chen et al. (unpublished results), a) the zebrafishª2937 to ª290 (ª2937/ª290) myf-5 upstream regulatory se-quence contains notochord-specific elements and b) in germ-line transmitted zebrafish, an myf-5 ª6212/ª1 segment fused with EGFP is capable of directing GFP expression in the spinal cord. Based on these obser-vations, we propose the possible presence of a neuron-specific regulatory element within the zebrafish myf-5 gene. Zeller and Granato (1999) demonstrated that a distinct population of myotomal cells plays a pivotal role in the early migration of zebrafish motor axons. They also identified the diwanka gene as a somite-derived cue that is required for establishing an axonal path from the spinal cord to the somites. Their evidence provides in-sight into how neuronal and myotomal precursor cells are linked, and raise the possibility that the zebrafish
myf-5 gene may follow the same pathway.
Comparing ZMF-morphants with known mutants Two types of somite-defective zebrafish mutants have been described and used for comparisons with myf5-morphants. The first, known as fss-type mutants, in-clude defects associated with the segmentation and epi-thelialization of the somatic mesoderm: fused somites (fss), beamters (bea), deadly sevens (des) and after eights (aei) (van Eeden et al., 1996). In each of these, the seg-mental expression of MyoD in unsegmented regions is lost, but it is expressed throughout the somatic meso-derm (van Eeden et al., 1996). In the present study, the
myoD transcripts were normally expressed in
myf5-mor-phants (Fig. 3), therefore we can rule out the possibility that myf5-morphants are fss-type mutants. Because they all result in U-shaped somites, the second group is known as type mutants; they include you (you),
you-too (yot), slow-muscle-omitted (smu), and sonic-you (syu)
(van Eeden et al., 1996; Schauerte et al., 1998; Barresi et al., 2000; Varga et al., 2001). While we noticed that the somite boundaries of myf5-morphants were missing, they could not be considered U-shaped (Fig. 2).
In other words, the myf5-morphants in our study dis-played somite patterning defects that are not pheno-typically similar to either fss- or you-type mutants. This makes them similar to myf5-morphants with such brain mutations as no isthmus (noi) and white tail (wit); the former lacks MHB, while the latter has some character-istics of fss-type mutants in addition to brain and spinal cord deficiencies (Brand et al., 1996; Jiang et al., 1996). A search of current databases failed to identify mutants that are phenotypically similar to the myf5-morphants observed in this study. We found that myf-5 in knock-down zebrafish is lethal to embryos, in the same way
that it is lethal to the embryos of knockout mice (Braun et al., 1992). These results indicate the importance of
myf-5 to early embryonic development. They may also
explain why such an upstream control gene as myf-5 does not present phenotypes that overlap with other genes that are involved in these development pathways.
Our conclusion is that even though Myf-5 is best known as a muscle regulatory factor in somites, ze-brafish Myf-5 is involved in the convergence and exten-sion of shield epiblasts, tail buds, and brain formation during early embryogenesis.
Acknowledgements We are grateful to Dr. Wen-Chang Chang,
Insti-tute of Biochemical Science, for providing plasmids containing cDNAs of pax2.1 and krox20. We are also grateful to Prof. Ruey-Ping Lin, Department of Zoology, for helping us with the cryosec-tion experiments. This research was supported by the Nacryosec-tional Science Council, Republic of China (grant no. NSC 90 – 2313-B002 – 260).
References
Barresi, M.J.F., Stickney, H.L. and Devoto, S.H. (2000) The ze-brafish slow-muscle-omitted gene product is required for Hedge-hog signal transduction and the development of the slow muscle identity. Development 127:2189–2199.
Brand, M., Heisenberg, C.P., Jiang, Y.J., Beuchle, D., Lun, K., Fu-rutani-Seiki, M., Granato, M., Haffter, P., Hammerschmidt, M., Kane, D., Kelsh, R., Mullins, M., Odenthal, J., van Eeden, F.J.M. and Nusslein-Volhard, C. (1996) Mutations in zebrafish genes affecting the formation of the boundary between midbrain and hindbrain. Development 123:179–190.
Braun, T., Rudnicki, M.A., Arnold, H.H. and Jaenisch, R. (1992) Targeted inactivation of the muscle regulatory gene Myf-5 results in abnormal rib development and perinatal death. Cell 71:369– 382.
Buonanno, A., Apone, L., Morasso, M.I., Beers, R., Brenner, H.R. and Eftimie, R. (1992) The MyoD family of myogenic factors is regulated by electrical activity: isolation and characterization of a mouse Myf-5 cDNA. Nucleic Acids Res 20:539–544.
Carvajal, J.J., Cox, D., Summerbell, D. and Rigby, P.W.J. (2001) A BAC transgenic analysis of the Mrf4/ Myf5locus reveals inter-digitated elements that control activation and maintenance of gene expression during muscle development. Development 128:1857–1868.
Chen, Y.H., Lee, W.C., Cheng, C.H. and Tsai, H.J. (2000) Muscle regulatory factor gene: zebrafish (Danio rerio) myogenin cDNA. Comp Biochem Physiol 127B:97–103.
Chen, Y.H., Lee, W.C., Liu, C.F. and Tsai, H.J. (2001) Molecular structure, dynamic expression and promoter analysis of zebrafish (Danio rerio) myf-5 gene. Genesis 29:22–35.
Chen, Y.H., Liang, C.T. and Tsai, H.J. (2002) Expression, purifi-cation and DNA-binding activity of tilapia muscle-specific tran-scription factor MyoD, produced in Escherichia coli. Comp Bio-chem Physiol 131B:795–805.
Coutelle, O., Blagden, C.S., Hampson, R., Halai, C., Rigby, P.W.J. and Hughes, S.M. (2001) Hedgehog signaling is required for maintenance of myf5 and myoD expression and timely terminal differentiation in zebrafish adaxial myogenesis. Dev Biol 236:136–150.
Daubas, P., Tajbakhsh, S., Hadchouel, J., Primig, M. and Buck-ingham, M. (2000) Myf5 is a novel early axonal marker in the mouse brain and is subjected to post-transcriptional regulation in neurons. Development 127:319–331.
456
Devoto, S.H., Melancon, E., Eisen, J.S. and Westerfield, M. (1996) Identification of separate slow and fast muscle precursor cells in vivo, prior to somite formation. Development 122:3371–3380. Dutton, K., Dutton, J.R., Pauliny, A. and Kelsh, R.N. (2001) A
morpholino phenocopy of the colourless mutant. Genesis 30:188–189.
Ekker, S.C. and Larson, J.D. (2001) Morphant technology in model developmental system. Genesis 30:89–93.
Fire, A., Xu, S., Montgomery, M.K., Kostas, S.A., Driver, S.E. and Mello, C.C. (1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391:806–811.
Fisher, S., Amacher, S.L. and Halpern, M.E. (1997) Loss of cer-ebrum function ventralizes the zebrafish embryo. Development 124:1301–1311.
Hadchouel, J., Tajbakhsh, S., Primig, M., Chang, T.H.T., Daubas, P., Rocancourt, D. and Buckingham, M. (2000) Modular long-range regulation of Myf5 reveals unexpected heterogeneity be-tween skeletal muscles in the mouse embryo. Development 127:4455–4467.
Hammerschmidt, M., Pelegri, F., Mullins, M.C., Kane, D.A., van Eeden, F.J., Granato, M., Brand, M., Furutani-Seiki, M., Hafft-er, P., Heisenberg, C.P., Jiang, Y.J., Kelsh, R.N., Odenthal, J., Warga, R.M. and Nusslein-Volhard, C. (1996) dino and merced-es, two genes regulating dorsal development in the zebrafish em-bryo. Development 123:95–102.
Hollenberg, S.M., Cheng, P.F. and Weintraub, H. (1993) Use of a conditional MyoD transcription factor in studies of MyoD transactivation and muscle determination. Proc Natl Acad Sci USA 90:8028–8032.
Hopwood, N.D., Pluck, A. and Gurdon, J.B. (1991) Xenopus Myf-5 marks early muscle cells and can activate muscle genes ec-topically in early embryos. Development 111:551–560.
Jiang, Y.J., Brand, M., Heisenberg, C.P., Beuchle, D., Furutani-Seiki, M., Kelsh, R.N., Warga, R.M., Granato, M., Haffter, P., Hammerschmidt, M., Kane, D.A., Mullins, M.C., Odenthal, J., van Eeden, F.J.M. and Nusslein-Volhard, C. (1996) Mutations affecting neurogenesis and brain morphology in the zebrafish, Danio rerio. Development 123:205–216.
Kablar, B. and Rudnicki, M.A. (1999) Development in the absence of skeletal muscle results in the sequential ablation of motor neu-rons from the spinal cord to the brain. Dev Biol 208:93–109. Kiefer, J.C. and Hauschka, S.D. (2001) Myf-5 is transiently
ex-pressed in nonmuscle mesoderm and exhibits dynamic regional changes within the presegmented mesoderm and somites I–IV. Dev Biol 232:77–90.
Kimmel, C.B., Ballard, W.W., Kimmel, S.R., Ullmann, B. and Schilling, T.F. (1995) Stages of embryonic development of the zebrafish. Dev Dyn 203:253–310.
Krause, M., Harrison, S.W., Xu, S.Q., Chen, L. and Fire, A. (1994) Elements regulating cell- and stage-specific expression of the C. elegans MyoD family homolog hlh-1. Dev Biol 166:133–148. Li, H. and Capetanaki, Y. (1993) Regulation of the mouse desmin
gene: transactivated by MyoD, myogenin, MRF4 and Myf5. Nucleic Acids Res 21:335–343.
Li, H, Choudhary, S.K., Milner, D.J., Munir, M.I., Kuisk, I.R. and Capetanaki, Y. (1994) Inhibition of desmin expression blocks myoblast fusion and interferes with the myogenic regulators MyoD and myogenin. J Cell Biol 124:827–841.
Lindon, C., Albagli, O., Pinset, C. and Montarras, D. (2001) Cell density-dependent induction of endogenous myogenin (myf-4) gene expression by Myf5. Dev Biol 240:574–584.
Loh, S.H., Chan, W.T., Gong, Z., Lim, T.M. and Chua, K.L. (2000) Characterization of a zebrafish (Danio rerio) desmin cDNA: an early molecular marker of myogenesis. Differentiation 65:247– 254.
Michelson, A.M., Abmayr, S.M., Bate, M., Arias, A.M. and Maniatis, T. (1990) Expression of a MyoD family member pre-figures muscle pattern in Drosophila embryos. Genes Dev 4:2086–2097.
Misquitta, L. and Paterson, B.M. (1999) Targeted disruption of gene function in Drosophila by interference (RNA-i): a role for nautilus in embryonic somatic muscle formation. Proc Natl Acad Sci USA 96:1451–1456.
Nasevicius, A. and Ekker, S.C. (2000) Effective targeted gene ‘‘knockdown’’ in zebrafish. Nature Genet 26:216–220.
Neyt, C., Jagla, K., Thisse, C., Thisse, B., Haines, L. and Currie, P.D. (2000) Evolutionary origins of vertebrate appendicular muscle. Nature 408:82–84.
Oppenheim, R.W. (1996) Neurotrophic survival molecules for mo-toneurons: an embarrassment of riches. Neuron 17:195–197. Oxtoby, E. and Jowett, T. (1993) Cloning of the zebrafish krox-20
gene (krx-20) and its expression during hindbrain development. Nucleic Acids Res 12:1087–1095.
Pfeffer, P.L., Gerster, T., Lun, K., Brand, M. and Busslinger, M. (1998) Characterization of three novel members of the zebrafish Pax2/5/8 family: dependency of Pax5 and Pax8 expression on the Pax2.1 (noi) function. Development 125:3063–3074.
Rudnicki, M.A., Braun, T., Hinuma, S. and Jaenisch, R. (1992) Inactivation of MyoD in mice leads to up-regulation of the my-ogenic HLH gene Myf-5 and results in apparently normal muscle development. Cell 71:383–390.
Rudnicki, M.A. and Jaenisch, R. (1995) The MyoD family of tran-scription factors and skeletal myogenesis. Bioessays 17:203–209. Rudnicki, M.A., Schnegelsberg, P.N., Stead, R.H., Braun, T., Ar-nold, H.H. and Jaenisch, R. (1993) MyoD or Myf-5 is required for the formation of skeletal muscle. Cell 75:1351–1359. Saitoh, O., Fujisawa-Sehara, A., Nabeshima, Y. and Periasamy, M.
(1993) Expression of myogenic factors in denervated chicken breast muscle: isolation of the chicken Myf5 gene. Nucleic Acids Res 21:2503–2509.
Schauerte, H.E., van Eeden, F.J.M., Fricke, C., Odenthal, J., Strah-le, U. and Haffter, P. (1998) Sonic Hedgehog is not required for floor plate induction in the zebrafish. Development 125:2983– 2993.
Scholpp, S. and Brand, M. (2001) Morpholino-induced knock-down of zebrafish engrailed genes eng2 and eng3 reveals redun-dant and unique functions in midbrain-hindbrain boundary de-velopment. Genesis 30:129–133.
Sporle, R., Gunther, T., Struwe, M. and Schughart, K. (1996) Se-vere defects in the formation of epaxial musculature in open brain (opb) mutant mouse embryos. Development 122:79–86. Tajbakhsh, S. and Buckingham, M.E. (1995) Lineage restriction of
the myogenic conversion factor myf-5 in the brain. Development 121:4077–4083.
van Eeden, F.J., Granato, M., Schach, U., Brand, M., Furutani-Sei-ki, M., Haffter, P., Hammerschmidt, M., Heisenberg, C.P., Jiang, Y.J., Kane, D.A., Kelsh, R.N., Mullins, M.C., Odenthal, J., War-ga, R.M., Allende, M.L., Weinberg, E.S. and Nüsslein-Volhard, C. (1996) Mutations affecting somite formation and patterning in the zebrafish, Danio rerio. Development 123:153–64.
Varga, Z.M., Amores, A., Lewis, K.E., Yan, Y.L., Postlethwait, J.H., Eisen, J.S. and Westerfield, M. (2001) Zebrafish smoothen-ed functions in ventral neural tube specification and axon tract formation. Development 128:3497–3509.
Weinberg, E.S., Allende, M.L., Kelly, C.S., Abdelhamid, A., Mu-rakami, T., Andermann, P., Doerre, O.G., Gruwald, D.J. and Riggleman, B. (1996) Developmental regulation of zebrafish MyoD in wild-type, no tail and spadetail embryos. Development 122:271–280.
Xu, Y., He, J., Wang, X., Lim, T.M. and Gong, Z. (2000) Asyn-chronous activation of 10 muscle-specific protein (MSP) genes during zebrafish somitogenesis. Dev Dyn 219:201–215.
Zeller, J. and Granato, M. (1999) The zebrafish diwanka gene con-trols an early step of motor growth cone migration. Develop-ment 126:3461–3472.
Zhao, Z., Cao, Y., Li, M. and Meng, A. (2001) Double-stranded RNA injection produces nonspecific defects in zebrafish. Dev Biol 229:215–223.