Basic Science Investigations
Respiration 093 DOI: 10.1159/000XXXXXX
Effects of Activated Protein C on
Ventilator-Induced Lung Injury in Rats
Jiunn-Song Jiang
a
Hsiu-Chu Chou
b
Leng-Fang Wang
c
Yaw-Dong Lang
d
Chung-Ming Chen
e
a Department of Internal Medicine, Shin Kong Wu Ho-Su Memorial Hospital and School of Respiratory Therapy, and
Departments of b Anatomy and c Biochemistry, College of Medicine, and d Graduate Institute of Medical Sciences,
Taipei Medical University, and e Department of Pediatrics, Taipei Medical University Hospital, Taipei , Taiwan, ROC
gas tension were comparable between the two ventilation groups throughout the study period. Rats treated with the HVZP protocol exhibited significantly higher total protein and macrophage inflammatory protein-2 concentrations in bronchoalveolar lavage fluid (BALF) and higher lung PAI-1 mRNA expression and plasma active PAI-1 levels than did the control group. Administration of APC tended to reduce the BALF protein content and systemic PAI-1 activity but did not improve the lung histology in the HVZP+APC group. Plasma levels of D-dimers were comparable among the three study groups. Conclusions: These results suggest that APC admin-istered at a higher dosage might improve lung function by reducing alveolar protein leakage and systemic coagula-tion. Copyright © 2010 S. Karger AG, Basel
Key Words
Bronchoalveolar lavage ⴢ Dimers ⴢ Macrophage inflammatory protein-2 ⴢ Plasminogen activator inhibitor-1 ⴢ Ventilator-induced lung injury
Abstract
Background: Mechanical ventilation with a high tidal vol-ume (V T ) increases lung and systemic plasminogen activator inhibitor (PAI)-1 levels and alveolar fibrin deposition. Acti-vated protein C (APC) may decrease PAI activity in endothe-lial cell-conditioned medium and thus enhance fibrinolysis. Objectives: The aims of this study were to test the hypoth-esis that APC can neutralize PAI-1 activity and improve lung function in an animal model of ventilator-induced lung in-jury. Methods: Rats were ventilated with a high-volume zero positive end-expiratory pressure (PEEP; HVZP) protocol by a volume-cycled ventilator for 2 h at a V T of 30 ml/kg, a respi-ratory rate of 25 breaths/min, and an Fi O 2 of 0.21. Fifteen
min-utes before ventilation, the rats received intravenous APC (250 g/kg, HVZP+APC group) or normal saline (vehicle; HVZP group). Another group that received no ventilation served as the control group. Results: Levels of arterial blood
Received: July 2, 2009
Accepted after revision: November 25, 2009 Published online: $ $ $
This study was supported by a grant from the Shin Kong Wu Ho-Su Memorial Hospital (SKH-TMU-97-04).
Jiang /Chou /Wang /Lang /Chen
Respiration 093
2
Introduction
Over the last several decades, mechanical ventilation has successfully been used to support patients with acute respiratory failure. However, it has several potential dis-advantages and complications regardless of the life-sav-ing potential of this assistance [1] . Mechanical ventilation with high tidal volumes (V T ) can damage the alveolar-capillary barrier and activates local and systemic inflam-matory responses. This course is referred to as ventilator-induced lung injury (VILI) [2] . The spectrum of VILI in-cludes disruption of endothelial and epithelial cells, and increases in endothelial and epithelial permeability and pulmonary and systemic inflammatory mediators [2–4] . Mechanisms implicated in VILI include alveolar epithe-lial and vascular endotheepithe-lial cellular responses to me-chanical stretching and inflammatory cell recruitment and activation [5, 6] . The physiological and histopatho-logical features of VILI are indistinguishable from those of the diffuse alveolar damage of acute lung injury. De-position of fibrin in the alveolar space and increased pro-coagulant activity of bronchoalveolar lavage fluid (BALF) are pathognomonic features of acute lung injury [7] .
Additional evidence suggests that mechanical ventila-tion may influence pulmonary fibrin turnover in VILI [8, 9] . Alveolar fibrin deposition leads to surfactant dysfunc-tion, poor gas exchange, decreased lung compliance, and increased ventilatory dependence, and accelerates the fi-brotic process [10, 11] . The mechanisms that contribute to increased alveolar fibrin turnover are increased local tissue factor-mediated thrombin generation and de-pressed bronchoalveolar urokinase plasminogen activa-tor-mediated fibrinolysis caused by increases in plasmin-ogen activator inhibitors (PAIs) [12] . PAI-1, a fibrinolytic antiprotease, is the major plasminogen inactivator in the plasma and the primary inhibitor of tissue- and uroki-nase-type plasminogen activators resulting in decreased plasmin activity and fibrinolytic potential [13] . We found that mechanical ventilation with a high V T and no posi-tive end-expiratory pressure (PEEP) increased lung and systemic PAI-1 levels and alveolar fibrin deposition [14] . Activated protein C (APC) decreases PAI activity in en-dothelial cell-conditioned medium and enhances fibri-nolysis by inhibiting PAI-1 of the whole blood and serum [15, 16] . Administration of APC may suppress plasma PAI activity observed after thrombolysis in patients with acute myocardial infarction [17] . Therefore, we hypoth-esized that APC may inhibit PAI-1 activity and improve lung function in an animal model of VILI. The aims of this study were to investigate the effects of APC on local
and systemic fibrinolytic activities in VILI and to find a potential treatment modality against high-V T -induced
lung injury.
Materials and Methods
Animal Preparations
This study was approved by the Animal Care and Use Commit-tee of the Taipei Medical University and was performed with adult male Sprague-Dawley rats weighing 250–300 g. Rats were main-tained on a 12-hour light-dark cycle with free access to food and water. Rats were intraperitoneally anesthetized with pentobarbital (50 mg/kg, Abbott, North Chicago, Ill., USA). Fifteen minutes be-fore ventilation, rats received recombinant human APC [250 g/ kg i.v., drotrecogin ␣ (activated); Xigris, Lilly, Giessen, Germany; n = 7; HVZP+APC group] or normal saline (HVZP group; n = 7) via the tail vein. A polyethylene tube catheter (PE-50, Becton Dick-inson, Sparks, Md., USA) containing isotonic saline was placed in one femoral artery to sample the blood for gas analysis. A trache-ostomy was performed, and a 14-gauge plastic cannula was in-serted into the trachea. Animals were then ventilated with a high-V T zero PEEP (HVZP) protocol by a volume-cycled ventilator (Small Animal Ventilator, model SAR-830/AP; CWE, Ardmore, Pa., USA) for 2 h at a V T of 30 ml/kg, zero PEEP, a respiratory rate of 25 breaths/min, and Fi O 2 of 0.21. Another group that received
no ventilation served as the control group (n = 7). All animals were kept supine for the duration of the experiment, and arterial blood gases were measured with a blood gas analyzer (model 1620; In-strumentation Laboratories, Lexington, Mass., USA) at the begin-ning and every hour after randomization.
Bronchoalveolar Lavage
After 2 h of ventilation, rats were killed with an intravenous injection of pentobarbital (100 mg/kg). Blood was collected from the femoral artery in citrated (0.109 M ) vacutainer tubes, placed on ice, and spun at 4 ° C, and the resulting plasma was stored at –70 ° C until analyzed. The chest was opened and the lungs were removed intact from the animal with the tracheostomy tube in place. The right lung was ligated, and the left lung was lavaged with 2 ml 0.9% saline at 4 ° C which was washed in and out of the lungs three times and then recovered. This washing procedure was repeated two more times for each animal, with the three washes being pooled, and the total volume recorded. There were no differences in the total volume of saline infused or recovered after the lavage procedure among the three experimental groups. The right superior lobe was fixed in 10% buffered formalin and embedded in paraffin. An aliquot of the BALF from each animal was used to measure the total protein content with bovine serum albumin as the standard and macrophage inflammatory protein (MIP)-2 using an enzyme-linked immunosorbent assay kit (R&D Systems, Minneapolis, Minn., USA); values are expressed as mil-ligrams per kilogram of body weight and picograms per milliliter of lavage fluid, respectively.
Lung PAI-1 mRNA Expression by Real-Time Polymerase Chain Reaction
Lung tissue was ground into a powder in liquid nitrogen, and PAI-1 mRNA expression was measured using real-time PCR. Total
RES093.indd 2
RNA was extracted using the TRIzol Reagent (Invitrogen Life Technologies, Paisley, UK). Reverse transcription was performed on 1 g of RNA with oligo-dT primers and avian myeloblastosis virus reverse transcriptase (Roche, Indianapolis, Ind., USA). Primer sequences for the SYBR green real-time PCR includ-ed: PAI-1 sense (5 ⴕ -ATGGCTCAGAACAACAAGTTCAAC-3 ⴕ ) and antisense (5 ⴕ -CAGTTCCAGGATGTCGTACTCG-3 ⴕ ), and GAPDH rRNA sense (5 ⴕ -ATGATTCTACCCACGGCAAG-3 ⴕ ) and antisense primers (5 ⴕ -CTGGAAGATGGTGATGGGTT-3 ⴕ ). Gene expression was quantitatively analyzed using the comparative CT ( ⌬ CT) method, in which CT is the threshold cycle number (the minimum number of cycles needed before the product can be de-tected). The arithmetic formula for the ⌬ CT method is the differ-ence in the number of threshold cycles for a target (PAI-1) and an endogenous reference (the GAPDH rRNA housekeeping gene).
PAI-1 and the Dimer Assay
PAI-1 was assayed by a commercially available assay kit that measures active PAI-1 (Innovative Research, Southfield, Mich., USA) in the plasma and BALF. Fibrinolytic activity was assessed by measuring plasma concentrations of the D-dimer with an ELI-SA kit purchased from American Diagnostica (Stamford, Conn., USA).
Immunohistochemistry of PAI-1
Immunohistochemical staining for PAI-1 was performed on paraffin sections with immunoperoxidase visualization. After deparaffinization in xylene and rehydration in an alcohol series, sections were first preincubated for 1 h at room temperature in 0.1 M phosphate-buffered saline containing 10% normal goat se-rum and 0.3% H 2 O 2 to block endogenous peroxidase activity and
nonspecific binding of the antibody before being incubated for 20 h at 4 ° C with a rabbit polyclonal antibody against rat PAI-1. Sections were then treated for 1 h at room temperature with bio-tinylated goat anti-rabbit IgG (1: 200; Vector Laboratories, Burlin-game, Calif., USA). This was followed by reaction with the re-agents from an ABC kit (Avidin-Biotin Complex, Vector) accord-ing to the manufacturer’s recommendations, and the reaction products were visualized by 3,3 diaminobenzidine and 0.003%
H 2 O 2 in 0.5 M Tris buffer (pH 7.6) before the sections were
mount-ed on gelatin-coatmount-ed slides using Permount (Fisher Scientific, Pittsburgh, Pa., USA). Sections for PAI-1 were mounted in glyc-erin gelatin and counterstained with hematoxylin. A minimum of four random lung fields of immunohistochemically stained sections per animal were captured with a digital camera and im-ported into the computerized image analysis system (Image-Pro Plus 5.1 for Windows). The automatic object counting and mea-suring process was used to quantify the immunoreactivity of the sections [14] . These generated a percentage of positively stained cells, and the value was expressed as a labeling index.
Histology
Specimens were embedded in paraffin, stained with hematox-ylin and eosin, and examined by a pathologist who was blinded to the protocol and experimental groups. Lung injury was scored according to the following items: (1) alveolar congestion, (2) hem-orrhage, (3) infiltration of neutrophils into the airspaces or the vessel wall, and (4) thickness of the alveolar wall [14] . Each item was graded according to a five-point scale: 0, minimal (little) damage; 1, mild damage; 2, moderate damage; 3, severe damage, and 4, maximal damage.
Statistical Analysis
The lung injury score data are given as medians and ranges, whereas other data are presented as means 8 SD. Statistically significant differences were analyzed by ANOVA followed by Tukey’s post hoc analysis. The Spearman test was used for corre-lation analyses of recorre-lationships between BALF protein contents and plasma active PAI-1 levels. Differences were considered sig-nificant at p ! 0.05.
Results Gas Exchange
The arterial blood gas tensions were comparable among the three study groups before mechanical ventila-tion ( table 1 ). Rats treated with HVZP ventilaventila-tion exhib-ited a higher mean pH and lower mean carbon dioxide tension throughout the study period.
Total Protein, MIP-2, and PAI-1 in the BALF
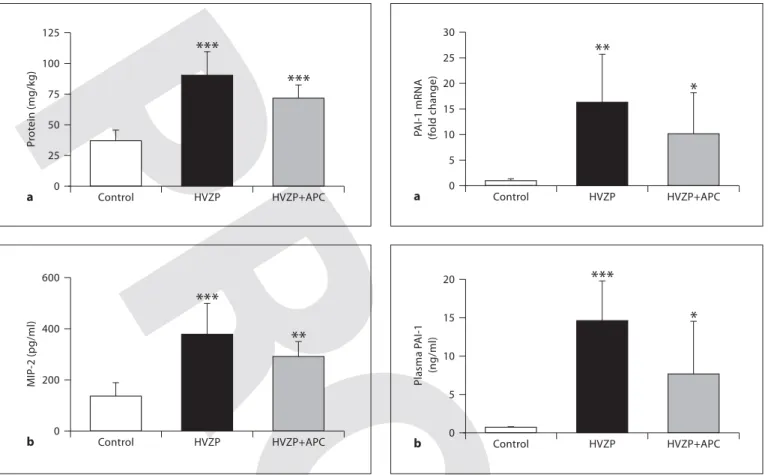
Total protein contents recovered from the BALF were significantly higher in rats ventilated with the HVZP and HVZP+APC protocols than in the control group ( fig. 1 a). In the HVZP+APC group, we found a trend towards de-creased BALF protein compared to the HVZP group (p = 0.059). MIP-2 concentrations in the BALF increased after HVZP ventilation, and the values were approximately 2-fold higher in the HVZP group compared to the control group ( fig. 1 b). The addition of APC did not significantly decrease MIP-2 levels. PAI-1 activity was undetectable in all BALF samples from control animals and was barely detectable in HVZP-treated animals.
Table 1. Arterial blood PaO2,PaCO2, andpH at baseline (0 h) and
after 1 and 2 h of ventilation
Group n pH PaO2 mm Hg PaCO2 mm Hg Control 7 0 h 7.4180.03 9288 4383 HVZP 7 0 h 7.4080.06 93815 43810 1 h 7.6280.06 11186 2184 2 h 7.6180.03 11088 2282 HVZP+APC 7 0 h 7.4280.04 95816 4283 1 h 7.6180.03 109811 2283 2 h 7.6380.02 101812 2383
Jiang /Chou /Wang /Lang /Chen
Respiration 093
4
Lung PAI-1 mRNA Expression and Plasma PAI-1 Levels
PAI-1 mRNA expression was significantly increased ( ⬃ 15-fold) in rats ventilated with the HVZP and HVZP+APC protocols in comparison to the control ani-mals ( fig. 2 a). Compared to the control group, rats treated with HVZP ventilation demonstrated significantly high-er plasma PAI-1 levels ( fig. 2 b). The HVZP+APC group tended to have lower PAI-1 levels than the HVZP group (p = 0.056).
Immunohistochemistry of PAI-1
PAI-1 immunoreactivity was mainly detected in air-way epithelial and mesenchymal cells ( fig. 3 a–c), and the
immunoreactivity markedly increased in rats treated with HVZP compared to the control and HVZP+APC groups by quantitative analysis ( fig. 3 d).
Plasma D-Dimers
Rats treated with the HVZP protocol exhibited non-significantly higher levels of plasma D-dimers compared to the control and HVZP+APC groups ( fig. 4 ).
Histology
After 2 h of ventilation, the HVZP and HVZP+APC groups had significantly higher lung injury scores than the control group ( table 2 ). Lung injury was characterized by alveolar congestion, hemorrhaging, and inflammato-0 P rot ein (mg/kg) 125 Control HVZP HVZP+APC *** *** a 25 50 75 100 0 MIP -2 (pg/ml) 200 400 600 Control HVZP HVZP+APC *** ** b
Fig. 1. Total protein and MIP-2 in BALF in the control
(non-ven-tilated), high-V T zero PEEP (HVZP), and HVZP+APC groups. n = 7/group. a The BALF protein content was significantly higher in the HVZP and HVZP+APC groups compared to control rats ( * * * p ! 0.001 vs. the control group). b MIP-2 recovered from BALF was significantly higher in rats in the HVZP and HVZP+APC groups than in the control group ( * * p ! 0.01 and * * * p ! 0.001, vs. the control group).
0 P AI-1 mRNA (f old change) 30 Control HVZP HVZP+APC ** * a 5 10 15 20 25 0 Plasma P AI-1 (ng/ml) 20 Control HVZP HVZP+APC *** * b 5 10 15
Fig. 2. Lung PAI-1 mRNA expression and plasma active PAI-1
levels in the control, HVZP, and HVZP+APC groups. Treatment details are given in the legend to figure 1. a Rats treated with the HVZP and HVZP+APC protocols had significantly higher levels of lung PAI-1 mRNA expression compared to the control group ( * p ! 0.05 and * * p ! 0.01, vs. the control group). b Compared to the control group, rats treated with the HVZP and HVZP+APC protocols demonstrated significantly higher plasma PAI-1 levels ( * p ! 0.05 and * * * p ! 0.001, vs. the control group).
RES093.indd 4
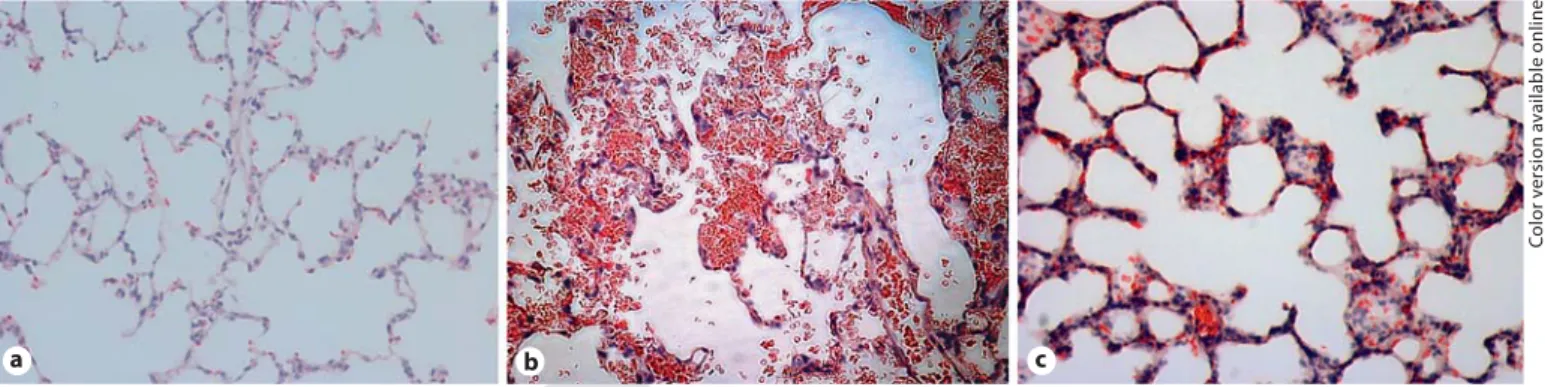
ry cell infiltration ( fig. 5 ). These findings are consistent with changes in alveolar damage found with acute lung injury. The HVZP group showed patchy areas of hem-orrhaging and thickened alveolar walls, and the HVZP+ APC group showed less hemorrhaging and alveolar con-gestion. No major histological abnormalities were pres-ent in the control animals.
Discussion
The main findings of this study are that high-V T ven-tilation is associated with increased lung capillary-alveo-lar leakage, BALF MIP-2 levels, lung PAI-1 mRNA ex-pression, and plasma active PAI-1 levels. The findings of our in vivo lung injury model are consistent with altera-tions known to occur in VILI. These data indicate that high-V T ventilation may enhance local and systemic
co-agulation and suppress systemic fibrinolysis. The BALF protein content is a measure of capillary-alveolar protein leakage which is due to physical disruption of the plasma membrane integrity. In this study, we found that admin-istration of APC did not significantly decrease the BALF protein content. These results are in contrast to the find-ings of Finigan et al. [18] , who reported that APC
treat-ment significantly decreased pulmonary leakage induced by mechanical ventilation when given either before or af-ter initiation of ventilation. This discrepancy can possi-bly be explained by different APC dosages and different magnitudes of lung injury.
0 P AI-1 labeling index (%) 50 Control HVZP HVZP+APC *** d 10 20 30 40
Fig. 3. Immunohistochemical staining for PAI-1 in the control
(non-ventilated; a ), HVZP ( b ), and HVZP+APC groups ( c ). ! 200 (online version: brown = positive staining). d Quantitative analy-sis of PAI-1 immunoreactivity. PAI-1 immunoreactivities were mainly detected in airway epithelial and some mesenchymal cells, and the immunoreactivity markedly increased in rats treated with HVZP compared to the control (non-ventilated) and HVZP+APC groups ( * * * p ! 0.001). a b c 0 Dimer (ng/ml) 30 Control HVZP HVZP+APC 5 10 15 20 25
Fig. 4. Plasma D-dimer in the control (non-ventilated), HVZP,
and HVZP+APC groups. Rats treated with HVZP ventilation had nonsignificantly higher plasma D-dimer levels compared to the control and HVZP+ACP groups.
Co lo r v e rs io n av a il a b le o n li n e
Jiang /Chou /Wang /Lang /Chen
Respiration 093
6
In this study, although rats treated with HVZP venti-lation exhibited lung tissue injury, arterial blood tension levels were comparable between the two study groups. These data suggest that monitoring of gas exchange is not a dependable indicator for the early recognition of VILI in previously healthy lungs. These results are supported by studies by Moriondo et al. [19] and Villar et al. [20] , who found satisfactory results of arterial blood gas analy-sis in rats with high-V T -induced lung tissue injury.
Increased procoagulant activity of the BALF is a pathognomonic feature of acute lung injury [7, 21] . In this study, PAI-1 activity was undetectable in all BALF sam-ples from control animals and barely detectable in HVZP-treated animals. We speculated that the difficulties in quantifying PAI-1 are because lung injury evolves rap-idly and makes the timing of the analysis critical or the BAL procedure dilutes the alveolar contents. We also found that the total protein in the BALF was positively correlated with plasma levels of PAI-1 in all study animals (r = 0.695, p ! 0.001). These results suggest that abnor-malities in coagulation and fibrinolysis may play
impor-tant roles in the pathogenesis of VILI. Animal models of VILI showed that high-V T ventilation increases alveolar fibrin deposition and systemic PAI-1 activity, and lung-protective mechanical ventilation decreases BALF levels of PAI-1, attenuates coagulation, and enhances fibrinoly-sis [14, 22] . APC exerts its anticoagulant activity by inac-tivating procoagulant factors Va and VIIIa and promotes fibrinolysis by inactivating PAI-1 [23] . Recombinant APC was shown to significantly reduce mortality in patients with severe sepsis [24] . Theoretical considerations sug-gest that anticoagulant therapy with APC might benefit animals with VILI. APC was shown not only to be an an-ticoagulant but also to have anti-inflammatory proper-ties [25, 26] . The distinctive combination of anticoagu-lant and anti-inflammatory properties of APC has made it is an important natural anticoagulant. However, APC did not improve lung function, although APC treatment tended to reduce systemic PAI-1 activity in this animal model of VILI.
D-dimers are derived from the degradation of cross-linked fibrin polymers, which come from clots, not
free-Table 2. Lung injury scores (medians and ranges)
Treatment n Alveolar Hemorrhage Neutrophil Alveolar wall Lung injury score
congestion infiltration thickness
Control 7 0 (0–1) 1 (0–2) 1 (0–1) 1 (1–2) 2 (1–3) HVZP 7 3 (2–4) 3 (1–4) 2 (2–4) 2 (1–2) 11 (6–14)*** HVZP+APC 7 2 (2–4) 2 (2–4) 2 (2–4) 1 (1–2) 7 (7–14)*** *** p < 0.001 vs. control. a b c
Fig. 5. Representative lung tissue photomicrographs ( ! 200). a Control group showing no major histological
abnormalities. b HVZP group showing patchy areas of hemorrhaging and thickened alveolar walls. c HVZP+APC group showing less hemorrhaging and alveolar congestion.
Co lo r v e rs io n av a il a b le o n li n e RES093.indd 6 RES093.indd 6 13.01.2010 09:03:5313.01.2010 09:03:53
ly circulating fibrin or fibrinogen. Elevated levels of D-dimers indicate increases in blood coagulation and turn-over of cross-linked intravascular fibrin activation [27] . In this murine model of VILI, we found that high-V T ven-tilation increased plasma levels of D-dimers, and APC treatment did not significantly decrease plasma D-dimer levels. These results suggest that APC treatment at this dosage cannot inhibit fibrin formation or activate fibri-nolysis.
APC is a serine protease with potent anti-inflamma-tory actions in addition to its anticoagulation properties [28] . The anti-inflammatory activity of APC depends on its ability to suppress the secretion of several cytokines [29] . MIP-2 is associated with leukocyte migration and activation, and was found to be closely associated with them in animal models of pneumonia and VILI [30, 31] . Intravenous APC significantly decreased BALF levels of MIP-2 in leukocyte elastase-induced lung injury in mice [32] . In this study, plasma levels of active PAI-1 tended to
significantly decrease, and BALF levels of MIP-2 were comparable to those with APC treatment. These changes imply that the anticoagulant effect and anti-inflamma-tory activity of APC are independent of each other. The major limitation of this study is a lack of a dose-response analysis. There are potential beneficial effects at the APC dose used, and the possibility exists that higher doses may be more efficacious in reducing VILI.
In conclusion, systemic coagulation-fibrinolysis ab-normalities induced by high-V T ventilation were
indicat-ed by increasindicat-ed pulmonary PAI-1 mRNA expression and plasma levels of PAI-1. Intravenous administration of APC tended to be associated with decreased alveolar pro-tein leakage and plasma levels of PAI-1 and with compa-rable BALF levels of MIP-2 but could not improve oxy-genation or the lung histology. We speculate that APC administered at a higher dosage might improve lung function by reducing alveolar protein leakage and sys-temic coagulation.
References
1 Sandur S, Stoller JK: Pulmonary complica-tions of mechanical ventilation. Clin Chest Med 1999; 20: 223–247.
2 Dos Santos CC, Slutsky AS: Invited review: mechanisms of ventilator-induced lung in-jury: a perspective. J Appl Physiol 2000; 89: 1645–1655.
3 Slutsky AS: Lung injury caused by mechani-cal ventilation. Chest 1999; 116: 9S–15S. 4 Tremblay LN, Slutsky AS:
Ventilator-in-duced injury: from barotrauma to biotrau-ma. Proc Assoc Am Physicians 1998; 110: 482–488.
5 Lionetti V, Recchia FA, Ranieri VM: Over-view of ventilator-induced lung injury mech-anisms. Curr Opin Crit Care 2005; 11: 82– 86.
6 Ren Y, Zhan Q, Hu Q, Sun B, Yang C, Wang C: Static stretch induces active morphologi-cal remodeling and functional impairment of alveolar epithelial cells. Respiration 2009; 78: 301–311.
7 Abraham E: Coagulation abnormalities in acute lung injury and sepsis. Am J Respir Cell Mol Biol 2000; 22: 401–404.
8 Dahlem P, Bos AP, Haitsma JJ, Schultz MJ, Meijers JC, Lachmann B: Alveolar fibrino-lytic capacity suppressed by injurious me-chanical ventilation. Intensive Care Med 2005; 31: 724–732.
9 Dahlem P, Bos AP, Haitsma JJ, et al: Mechan-ical ventilation affects alveolar fibrinolysis in LPS-induced lung injury. Eur Respir J 2006; 28: 992–998.
10 Idell S: Coagulation, fibrinolysis, and fibrin deposition in acute lung injury. Crit Care Med 2003; 31(4 suppl):S213–S220.
11 Marshall R, Bellingan G, Laurent G: The acute respiratory distress syndrome: fibrosis in the fast lane. Thorax 1998; 53: 815–817. 12 Choi G, Schultz MJ, Levi M, van der Poll T,
Millo JL, Garrard CS: Protein C in pneumo-nia. Thorax 2005; 60: 705–706.
13 Vassalli JD, Sappino AP, Belin D: The plas-minogen activator/plasmin system. J Clin Invest 1991; 88: 1067–1072.
14 Chen CM, Chou HC, Wang LF, Lang YD: Captopril decreases plasminogen activator inhibitor-1 in rats with ventilator-induced lung injury. Crit Care Med 2008; 36: 1880– 1885.
15 van Hinsbergh VW, Bertina RM, van Wijn-gaarden A, van Tilburg NH, Emeis JJ, Haver-kate F: Activated protein C decreases plas-minogen activator-inhibitor activity in en-dothelial cell-conditioned medium. Blood 1985; 65: 444–451.
16 Sakata Y, Loskutoff DJ, Gladson CL, Hek-man CM, Griffin JH: Mechanism of protein C-dependent clot lysis: role of plasminogen activator inhibitor. Blood 1986; 68: 1218– 1223.
17 Sakamoto T, Ogawa H, Takazoe K, et al: Ef-fect of activated protein C on plasma plas-minogen activator inhibitor activity in pa-tients with acute myocardial infarction treated with alteplase: comparison with un-fractionated heparin. J Am Coll Cardiol 2003; 42: 1389–1394.
18 Finigan JH, Boueiz A, Wilkinson E, et al: Ac-tivated protein C protects against ventilator-induced pulmonary capillary leak. Am J Physiol 2009; 296:L1002–L1011.
19 Moriondo A, Pelosi P, Passi A, et al: Proteo-glycan fragmentation and respiratory me-chanics in mechanically ventilated healthy rats. J Appl Physiol 2007; 103: 747–756. 20 Villar J, Herrera-Abreu MT, Valladares F,
Muros M, Pérez-Méndez L, Flores C, Kac-marek RM: Experimental ventilator-induced lung injury: exacerbation by positive end-ex-piratory pressure. Anesthesiology 2009; 110: 1341–1347.
21 Ware LB, Matthay MA, Parsons PE, Thomp-son BT, Januzzi JL, Eisner MD, National Heart, Lung, and Blood Institute Acute Re-spiratory Distress Syndrome Clinical Trials Network:. Pathogenetic and prognostic sig-nificance of altered coagulation and fibrino-lysis in acute lung injury/acute respiratory distress syndrome. Crit Care Med 2007; 35: 1821–1828.
22 Ware LB, Camerer E, Welty-Wolf K, Schultz MJ, Matthay MA: Bench to bedside: target-ing coagulation and fibrinolysis in acute lung injury. Am J Physiol Lung Lung Cell Mol Physiol 2006; 291:L307–L311.
23 Dahlbäck B, Villoutreix BO: Regulation of blood coagulation by the protein C antico-agulant pathway: novel insights into struc-ture-function relationships and molecular recognition. Arterioscler Thromb Vasc Biol 2005; 25: 1311–1320.
Jiang /Chou /Wang /Lang /Chen
Respiration 093
8
24 Bernard GR, Vincent JL, Laterre PF, et al: Ef-ficacy and safety of recombinant human ac-tivated protein C for severe sepsis. N Engl J Med 2001; 344: 699–709.
25 Esmon CT: Interactions between the innate immune and blood coagulation systems. Trends Immunol 2004; 25: 536–542.
26 Riewald M, Petrovan RJ, Donner A, Mueller BM, Ruf W: Activation of endothelial cell protease activated receptor 1 by the protein C pathway. Science 2002; 296: 1880–1882.
27 Huether S, McCance K (ed): Understanding Pathophysiology, ed 2. St. Louis, Mosby, 2000.
28 Esmon CT: Cell mediated events that control blood coagulation and vascular injury (re-view). Annu Rev Cell Biol 1993; 9: 1–26. 29 Esmon CT, Gu JM, Xu J, Qu D,
Stearns-Ku-rosawa DJ, KuStearns-Ku-rosawa S: Regulation and functions of the protein C anticoagulant pathway. Haematologica 1999; 84: 363–368.
30 Bonville C, Bennett N, Koehnlein M, et al: Respiratory dysfunction and proinflamma-tory chemokines in the pneumonia virus of mice (PVM) model of viral bronchiolitis. Vi-rology 2006; 349: 87–95.
31 Jiang JS, Wang LF, Chou HC, Chen CM: An-giotensin-converting enzyme inhibitor cap-topril attenuates ventilation-induced lung injury in rats. J Appl Physiol 2007; 102: 2098– 2103.
32 Tanaka A, Minoguchi K, Chen X, et al: Acti-vated protein C attenuates leukocyte elas-tase-induced lung injury in mice. Shock 2008; 30: 153–158.
RES093.indd 8