行政院國家科學委員會補助專題研究計畫成果報告
※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ 周 邊 血 液 單 核 細 胞 內 C 型 肝 炎 病 毒 之 分 子 病 毒 學 研 究 ( 2 / 2 ) ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ ※ 計畫類別:■個別型計畫 □整合型計畫 計畫編號:NSC89-2315-B-002-009- 執行期間:88 年 8 月 1 日至 89 年 7 月 31 日 計畫主持人:高嘉宏 共同主持人:陳定信 執行單位:國立台灣大學醫學院 中 華 民 國 89 年 10 月 12 日中文摘要
自從 C 型肝炎抗體檢驗試劑問世以來,吾人已確知大多數之慢性非 A 非 B 型肝炎 (non-A, non-B hepatitis) 乃由 C 型肝炎病毒 (hepatitis C virus, HCV) 所引起。近年對 C 型肝炎病毒 的分子生物學與感染後的臨床病理病程研究皆有長足之進步,但對 C 型肝炎病毒的持續感 染機轉及病毒在慢性 C 型肝炎病患周邊血液單核細胞 (peripheral blood mononuclear cell, PBMC) 內之病毒學意義則尚待加強。為釐清上述極具重要性但懸而未決的問題,吾人在 過去兩年內設計一系列的實驗由各個層面來探討 C 型肝炎病毒感染人類周邊血液單核細胞 的機轉,諸如(1) 定量血清與周邊血液單核細胞之病毒濃度,藉此瞭解宿主細胞中病毒正 股 RNA 和負股 RNA (複製中間體) 之比例並評估病毒複製速率之高低;(2) 探討血清與周 邊血液單核細胞內病毒之物種類似 (quasispecies) 程度有無差異和 (3) 尋找病毒基因體外 套高度變異區 (hypervariable region, HVR-1) 有無存在特定之胺基酸序列以決定病毒之細 胞趨向性。本系列研究之結果顯示 (1) 血清中病毒之平均濃度為 1.5 104 copies/mL,而周 邊血液單核細胞中正股 RNA 和負股 RNA 之平均濃度分別為 7.8 102和 2.7 102 copies/μg RNA。其中正股 RNA 濃度約為負股 RNA 之 1~10 倍,顯示 HCV 在周邊血液單核細胞內之 複製效率並不高。(2) HCV 在血清和周邊血液單核細胞內之平均 quasispecies complexity 分 別 為 5.4+1.7 和 7.0+2.5 , 顯 示 不 同 生 存 環 境 對 HCV 有 不 同 的 選 擇 壓 力 (selection pressure)。(3) HVR-1 之胺基酸 402 (L) 似乎和 HCV 感染周邊血液單核細胞有密切關係, 但需做進一步確認。這些成果將有助於闡明 C 型肝炎病毒持續感染和其細胞趨向性的部分 機轉,相信對未來相關分子病毒學與免疫學的研究會有相當程度的影響,同時也將裨益於 有關慢性 C 型肝炎的治療與預防性疫苗的研發。 關鍵詞:C 型肝炎病毒,周邊血液單核細胞。
Abstr act
With the recent advent of cloning a novel hepatitis C virus (HCV) and subsequent development of several serological assays, at present HCV has been recognized as the major etiologic agent of parenteral non-A, non-B hepatitis worldwide [6, 14, 44, 45, 54]. In order to clarify the biological significance and clinical implications of HCV in PBMC from patients with chronic type C hepatitis, we have conducted a series of studies including (1) quantitation of HCV in serum as well as in PBMC and estimation of the frequency of HCV-infected PBMC; (2) whether diversity of virus quasispecies exists between serum and PBMC; (3) whether specific sequences for receptor recognition exist in the HVR-1 region of HCV genome to determine the PBMC tropism in the past two years. Our results showed that (1) the average serum HCV titer is 1.5x104 copies/mL and the average titer of positive-strand and negative-strand HCV RNA in PBMC is 7.8x102 and 2.7x102 copies/ug RNA. In addition, the titer of positive-strand HCV RNA in PBMC is 1 to 10 times higher than that of negative-strand HCV RNA in PBMC, suggesting a low replication efficiency of HCV in such cells. (2) The quasispecies complexity of HCV genome in serum and PBMC is 5.4+1.7 and 7.0+2.5, suggesting different selection pressure in different tissue compartments. (3) The aa 402(L) of HVR-1 may be associated with the PBMC tropism of HCV; however, further studies are warranted to confirm this preliminary finding. Based on these results, we could understand, at least in part, the pathogenesis of viral persistence and cell tropism of HCV. In the meantime, these data will shed much light on the establishment of culture system for HCV infection and will significantly influence our future strategies of hepatitis C vaccine development and antiviral therapy.
Introduction
Since the application of sensitive serologic markers for the diagnosis of hepatitis A and hepatitis B infections, it becomes clear that there are other hepatitis viruses designating non-A, non-B hepatitis viruses [2]. Because of lacking reliable markers, the diagnosis of non-A, non-B hepatitis (NANBH) required the exclusion of ongoing HAV or HBV infections and hepatotoxin or drug exposure [19]. Clinically, more than 50% of acute NANBH cases develop chronic hepatitis and 20% of them will progress to liver cirrhosis [19, 80, 81]. Besides, a potentially causal association of NANB virus infection with hepatocellular carcinoma (HCC) has been reported [42, 68]. With the recent advent of cloning a novel hepatitis C virus (HCV) and subsequent development of several serological assays, at present HCV has been recognized as the major etiologic agent of parenteral non-A, non-B hepatitis worldwide [6, 14, 44, 45, 54]. The characteristics of HCV have been described [11, 15, 31, 40, 64, 67, 77]. It is an enveloped, 50-60 nm virus with a single positive-stranded RNA genome of 9,500 nucleotides. There is some homology between the HCV and the flavivirus or pestivirus group. The genome organization of HCV is similar to that of pestiviruses or flaviviruses and consists of 5' and 3' untranslated regions (5UT and 3UT) flanking the virus structural (core, envelope) and nonstructural region 1/envelope region 2 (E2/NS1) and nonstructural regions 2 to 5 (NS2 - NS5) [31]. Several HCV isolates cloned from different geographic areas show significant sequence divergence [11, 15, 40, 64, 67, 77]. Recent phylogenetic analysis of the NS5 region of HCV genome has classified the virus into 6 major groups and 11 subtypes at least 6 major genotypes of HCV have been proposed to classify all of the reported isolates [4, 76], but only genomes of 4 subtypes (1a, 1b, 2a and 2b) have been wholly sequenced [11, 15, 40, 64, 67, 77]. The 5UT region of HCV is the most conserved among different HCV isolates and has been shown useful in detecting the virus by polymerase chain reaction (PCR) assay [4, 11, 31, 38, 66]. By contrast, the envelope region (E1) and E2/NS1 are the moderately variable and hypervariable portions among different subtypes of HCV genomes, respectively [4, 11, 15, 27, 28, 31, 43, 58, 62, 63, 64, 67, 77, 82, 83]. In recent reports, the genotypes and the amino acid heterogeneity in the variable regions of E2/NS1 viral protein of HCV have been claimed to be closely associated with the response to interferon-α therapy [86]. Because of the sequence diversity, the envelope region can be used as an effective tool to elucidate unresolved issues about HCV infections such as tracing transmission routes [35, 38] as well as correlating the interrelationships between the acute exacerbations of chronic type C hepatitis and the evolution of hepatitis C viral genomes [34]. However, there had been no efficient primers suitable for amplification of the envelope region until recently [17, 23, 36, 66].
Although direct sequencing of the amplified hypervariable region has been used to identify the main HCV subtype [11, 15, 40, 64, 67, 77], it is sophisticated and time-consuming and thus cannot be applied to a large number of samples. Recently, several easier genotyping methods for
HCV have been introduced and the incidence of genotypes in different countries has been determined [76]. Moreover, the genotypes of HCV have been claimed to be associated with the severity of chronic hepatitis and the response to interferon-α therapy [86]. By using a PCR typing assay with specific primers, we have demonstrated that type 1b virus is the predominant genotype in chronic type C hepatitis in Taiwan [36, 39], and type 1b virus as well as mixed infection may trigger more severe liver disease [34, 37].
Chronic viral hepatitis, liver cirrhosis and hepatocellular carcinoma are common in Taiwan and most are HBV-related [8, 10, 85]. The possible etiology accounting for the remaining chronic liver disease in Taiwan has been less mentioned until Chen et al [6] reported that 65% of HBsAg negative patients with chronic hepatitis, 44% of HBsAg negative cirrhotics and 63% of HCC patients seronegative for HBsAg are positive for anti-HCV. These data indicate that HCV infection plays an important role in HBsAg negative patients with chronic liver disease in Taiwan.
The high frequency of acute HCV infection evolving into chronicity [19, 80, 81], like human immunodeficiency virus (HIV) infection, is rather unusual as compared to other virus-related diseases. The possible mechanisms of viral persistence are as follows [69]: (1) tolerance to viral antigens as the hepatitis B e antigen in the perinatal transmission of HBV; (2) immunosuppression and infection of lymphoid tissue as HIV and Epstein-Barr virus (EBV) [12, 32]; (3) antigenic variation of virus as HIV and HDV [29, 30, 51, 57, 70]; (4) inaccessibility of the immune system to the infected cells like the HIV-infected neuron cells. Among these possibilities, lack of adequate host immunity [13, 22, 34, 65, 71], frequent viral antigen mutation [21, 27, 62, 63] and infection of lymphocytes by organ-specific selection of viral variants [1, 3, 24, 61, 72, 79, 87] which enable HCV to escape from host immune pressures may be most likely involved in the pathogenesis of HCV persistence. Viral variants and their cell tropism have been documented to play major roles in the persistent infection of viruses. Recent studies of HIV indicated that antigenic variation in the hypervariable V3 loop sequence of envelope glycoprotein (gp120) not only can escape from host immune response, but also contributes to the determination of lymphocyte versus macrophage tropism [12, 32, 33]. Several recent studies revealed that replication intermediates of HCV can be demonstrated in the peripheral blood mononuclear cells (PBMC) of a proportion (70%-100%) of chronic HCV-infected patients, implying HCV may infect PBMC [24, 61, 72, 79, 87]. Whether these also contribute to the possible mechanisms of HCV persistence remains unknown. Moreover, a recent report indicated that determination of HCV RNA in PBMCs at the end of interferon (IFN) therapy may predict whether the disease will relapse after IFN discontinuation [25], suggesting the presence of HCV in PBMC may have its clinical implications.
Although several patterns of serum ALT activities in chronic hepatitis C have been identified [39], the viral mechanisms of fluctuating hepatitis activities are far from clear. Recent studies based on chimpanzee experiments and human observations have revealed partly pathogenesis
responsible for episodes of hepatitis flares in chronic HCV carriers [22, 34, 65, 71]. In chronic hepatitis B, superinfection with other hepatitis viruses and reactivation of the original hepatitis B virus are two major causes of clinical exacerbations [7, 10, 18, 49, 55, 73]. Whether similar mechanisms also contribute to the acute exacerbations of chronic hepatitis C remains to be explored. Several cross-challenge studies in chimpanzees have provided evidence that reinfection with either homotypic or heterotypic HCV along with chronic HCV carriage does occur [22, 23, 65, 71]. In human beings, Lai et al. [47] indicated that the multiple episodes of acute hepatitis in 2 of their 3 polytransfused thalassemic children are due to reinfection with a different HCV strain and in the third may be due to reactivation of primary infection. Our previous studies indicated that superinfection of heterotypic HCV indeed occurs in humans and mixed infections of heterotypic HCV may be important in causing hepatitis flares of chronic hepatitis C, and thus also reinforced the concept of multiple infection with HCV as well [34, 37].
The influence of virus amount of HCV on clinicopathological course of chronic type C hepatitis has been demonstrated [26, 46, 50]. In addition, viral genetic variation during chronic infection has been documented in hepatitis viruses [5, 16, 48, 52]. For chronic type B hepatitis, severe liver damage is related to the clustering missense mutation in the core gene [16]. Similarly, the evolution rates of HDV genomes appear to be correlated with the changes of clinical pictures of hepatitis [52]. Thus, the more drastic the change of symptoms of hepatitis, either related to HBV or HDV, the more nucleotide changes are detected. HCV, like many other RNA viruses, conforms extensive variability in genomic sequences [20, 29, 56], especially in the 5' end of E2/NS1 region (nt. 1156-1233, 78 nucleotides, 26 amino acids) so-called hypervariable region (HVR-1), appears to be one of the major targets of host immune response and the hypervariation is a result of the selection pressure like the V3 loop of HIV-1 [4, 11, 15, 21, 27, 28, 31, 43, 58, 62-64, 67, 77, 82-84]. Our recent data showed that quasispecies nature of HCV genomes exists in humans and patients with different clinicopathologic course of chronic type C hepatitis may have sequential variations in virus amounts as well as neutralization epitopes of the HVR-1 [35a].
The fact that no suitable animal model or in vitro cell culture system for experimental HCV infection except chimpanzee inoculation limits the biologic assay of HCV. Thus it is imperative to develop either a less expensive small animal model or an in vitro system for propagating HCV. Two recent reports have demonstrated that some human T cell leukemia lines can support the replication if HCV [74, 75], and this prompted us to establish our own in vitro cell culture system and small animal model for HCV propagation.
In order to clarify the biological significance and clinical implications of HCV in PBMC from patients with chronic type C hepatitis, in the following two years a series of studies including (1) quantitation of HCV in serum as well as in PBMC and estimation of the frequency of HCV-infected PBMC; (2) whether diversity of virus quasispecies exists between serum and PBMC; (3) whether specific sequences for receptor
recognition exist in the HVR-1 region of HCV genome to determine the PBMC tropism will be performed in our laboratory (Hepatitis Research Center, National Taiwan University Hospital).
Patients and Methods
I. Patients:A total of 30 patients with chronic type C hepatitis have been enrolled, and paired plasma and PBMC samples from these patients were stored at -70oC until used.
II. Methods:
1. PBMC and plasma are separated using Ficoll-Hypaque gradient centrifugation. 2. Extraction of ribonucleic acids:
HCV RNA in serum and PBMC are extracted by single-step method with acid guanidinium thiocyanate-phenol-chloroform.
3. Reverse transcription and nested polymerase chain reaction (RT-PCR) genotyping with type-specific primers [38, 39, 78]. By distinct sizes of PCR products, 4 genotypes (1a, 1b, 2a and 2b) of HCV can be identified.
4. Amplification of the HVR-1 of HCV genome [34-36, 38, 53]. The primers that produce a PCR product encompassing E1 and HVR-1 regions are designed after comparing different genotypes of HCV (type 1a to 2b) and have been reported [11, 15, 64, 67].
5. To detect negative or antigenomic strand (replication intermediate) of HCV in PBMC, outer sense PCR primers instead of random primers are used in cDNA synthesis. After cDNA synthesis and heat inactivation of reverse transcriptase at 95oC for 2 hours, RNase A (0.1 µg) is added subsequently (37oC for 30 minutes) to ensure removal of residual positive strand or genomic HCV RNA [59].
6. The sequences of amplified HVR-1 PCR products are determined by using fluorescence labelled primers with 373A Sequencer (Applied Biosystems, Foster City, CA)
7. Comparative sequence analysis of the HVR-1 from serum and PBMC to detect conserved domain that can be specific for receptor recognition
8. Selected amplified HVR-1 PCR products from serum and PBMC are ligated to pCR-ScriptTM SK(+) vectors and then transformed to E. coli XL1-Blue MRF' competent cells (Stratagene, La Jolla, CA). Plasmid DNAs are extracted from white colonies by WinardTM minipress DNA purification system (Promega, Madison, WI).The sequences of inserted DNA are also determined by an automatic sequencer.
9. Quantitation of HCV RNA (positive and negative strands) in serum (copies/mL serum), PBMC (copies/µg RNA) and by a competitive PCR assay [53]. The estimated frequencies of HCV-infected PBMC are calculated accordingly.
Results
1.HCV RNA level in PBMC of patients with chronic type C hepatitis
Table 1. Clinical and laboratory data of 10 patients with chronic type C hepatitis. ________________________________________________________________________
HCV genotype
____________________________________
Serum Plus-strand Minus-strand
Sex, age ALT ________________ ________________
Patient (years) (U/L) Serum PBMC Serum PBMC
________________________________________________________________________ 1 F, 52 51 1b 1b –* 1b 2 F, 59 157 1b 1b – 1b 3 M, 54 16 2a 2a – 2a 4 F, 47 57 1b 1b – 1b 5 M, 58 48 1b 1b – 1b 6 F, 73 191 1b 1b – 1b 7 F, 63 24 1b 1b – 1b 8 M, 54 56 2a 2a – 2a 9 M, 55 52 1b 1b – 1b 10 M, 73 123 1b 1b – 1b ________________________________________________________________________
Note. ALT, alanine aminotransferase; M, male; F, female; PBMC, peripheral blood mononuclear cell.
Table 2. HCV titers of serum , plus- and minus-strand in PBMC of 10 patients with chronic type C hepatitis.
________________________________________________________________________ Serum titer Titer in PBMC (copies/µg RNA)
Patient Genotype (copies/mL) Plus-strand Minus-strand
________________________________________________________________________ 1 1b 5x102 102 102 2 1b 5x103 103 102 3 2a 5x102 102 102 4 1b 2.5x104 102 10 5 1b 2.5x103 5x102 102 6 1b 5x102 103 102 7 1b 5x103 103 103 8 2a 5x10 103 102 9 1b 104 2x103 103 10 1b 105 103 102 ________________________________________________________________________ Average 1.5x104 7.8x102 2.7x102 ________________________________________________________________________ Note. PBMC, peripheral blood mononuclear cell.
2. Degr ee of quasispecies of HCV in PBMC of patients with chronic type C hepatitis Table 1. Clinical and laboratory characteristics of 5 patients with chronic hepatitis C included in the study
________________________________________________________________________
Patient Sex/ Peak serum HCV HCV RNA*
No. Age (yr) ALT (units/L) Genotype Plasma PBMC
________________________________________________________________________ 1 M/65 52 1b +/– +/+ 2 M/73 254 1b +/– +/+ 3 M/58 84 1b +/– +/+ 4 F/47 69 1b +/– +/+ 5 F/61 152 1b +/– +/+ ________________________________________________________________________ NOTE. M. male; F, female; ALT, alanine aminotransferase; PBMC, peripheral blood mononuclear cells.
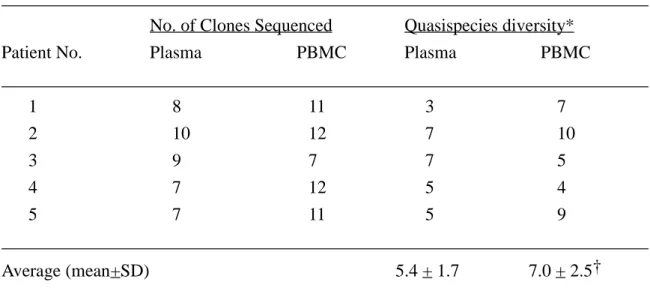
Table 2. Comparison of the quasispecies diversity of HCV genome between plasma and peripheral blood mononuclear cells
________________________________________________________________________ No. of Clones Sequenced Quasispecies diversity*
Patient No. Plasma PBMC Plasma PBMC
________________________________________________________________________ 1 8 11 3 7 2 10 12 7 10 3 9 7 7 5 4 7 12 5 4 5 7 11 5 9 ________________________________________________________________________ Average (mean+SD) 5.4 + 1.7 7.0 + 2.5† ________________________________________________________________________ NOTE. PBMC, peripheral blood mononuclear cells.
* No. of clones with different amino acid sequences. † Statistically non-significant by Student's t test.
3. Specific sequences in the HVR-1 region of HCV genome and PBMC tr opism
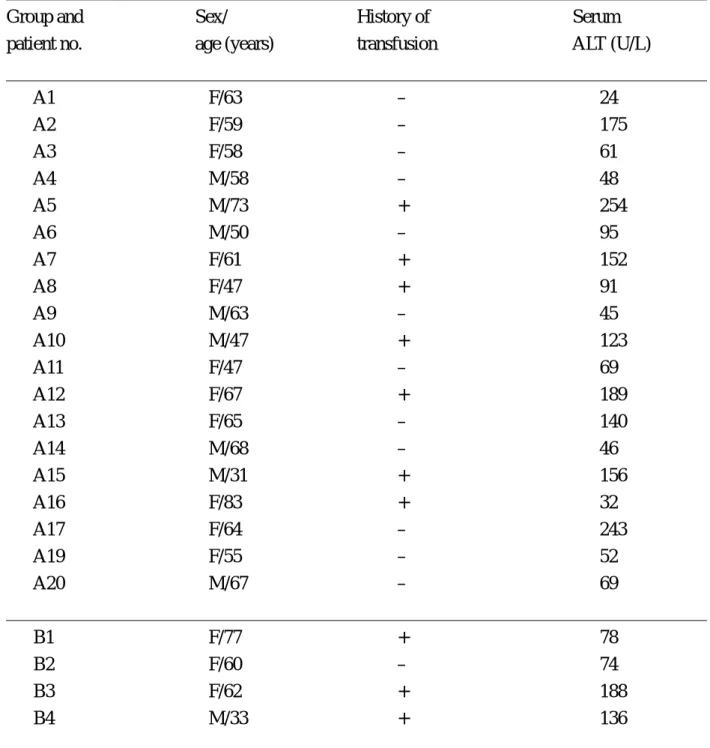
Table 1 Demographic and clinical data of group A and group B chronic hepatitis C patients
______________________________________________________________________________
Group and Sex/ History of Serum
patient no. age (years) transfusion ALT (U/L)
______________________________________________________________________________ A1 F/63 – 24 A2 F/59 – 175 A3 F/58 – 61 A4 M/58 – 48 A5 M/73 + 254 A6 M/50 – 95 A7 F/61 + 152 A8 F/47 + 91 A9 M/63 – 45 A10 M/47 + 123 A11 F/47 – 69 A12 F/67 + 189 A13 F/65 – 140 A14 M/68 – 46 A15 M/31 + 156 A16 F/83 + 32 A17 F/64 – 243 A19 F/55 – 52 A20 M/67 – 69 ______________________________________________________________________________ B1 F/77 + 78 B2 F/60 – 74 B3 F/62 + 188 B4 M/33 + 136 ______________________________________________________________________________ Group A: chronic hepatitis C patients with HCV infection of peripheral blood mononuclear cells; Group B: chronic hepatitis C patients without HCV infection of peripheral blood mononuclear cells;
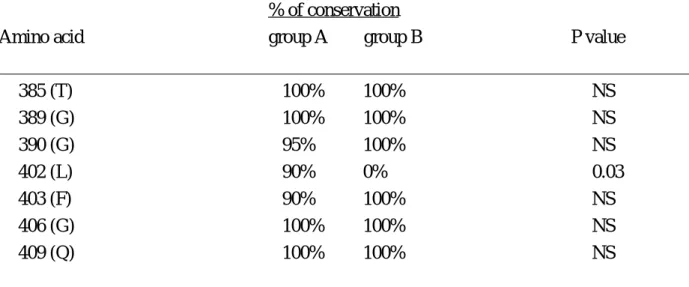
Table 2 Conservation of amino acids in the hypervariable region of hepatitis C virus genome between group A and group B chronic hepatitis C patients
______________________________________________________________________________ % of conservation
Amino acid group A group B P value
______________________________________________________________________________ 385 (T) 100% 100% NS 389 (G) 100% 100% NS 390 (G) 95% 100% NS 402 (L) 90% 0% 0.03 403 (F) 90% 100% NS 406 (G) 100% 100% NS 409 (Q) 100% 100% NS ______________________________________________________________________________ Group A: chronic hepatitis C patients with HCV infection of peripheral blood mononuclear cells; Group B: chronic hepatitis C patients without HCV infection of peripheral blood mononuclear cells.
Discussion
Our results showed that (1) the average serum HCV titer is 1.5x104 copies/mL and the average titer of positive-strand and negative-strand HCV RNA in PBMC is 7.8x102 and 2.7x102 copies/ug RNA. In addition, the titer of positive-strand HCV RNA in PBMC is 1 to 10 times higher than that of negative-strand HCV RNA in PBMC, suggesting a low replication efficiency of HCV in such cells. (2) The quasispecies complexity of HCV genome in serum and PBMC is 5.4+1.7 and 7.0+2.5, suggesting different selection pressure in different tissue compartments. (3) The aa 402(L) of HVR-1 may be associated with the PBMC tropism of HCV; however, further studies are warranted to confirm this preliminary finding. Based on these results, we could understand, at least in part, the pathogenesis of viral persistence and cell tropism of HCV. In the meantime, these data will shed much light on the establishment of culture system for HCV infection and will significantly influence our future strategies of hepatitis C vaccine development and antiviral therapy.
Refer ences
1. Ahmed R, Oldstone MBA. Organ-specific selection of viral variants during chronic infection. J Exp Med 1988; 167: 1719-24.
2. Alter HJ, Purcell RH, Holland PV, et al. Clinical and serological analysis of transfusion-associated hepatitis. Lancet 1975; 2: 838-41.
3. Bouffard P, Hayashi PH, Acevedo R, Levy N, Zeldis JB. Hepatitis C virus is detected in a monocyte/macrophages subpopulation of peripheral blood mononuclear cells of infected patients. J Infect Dis 1992; 166: 1276-80.
4. Bukh J, Purcell RH, Miller RH. At least 12 genotypes of hepatitis C virus predicted by sequence analysis of the putative E1 gene of isolates collected worldwide. Proc Natl Acad Sci USA 1993; 90: 8234-8.
5. Carmen W, Thomas H, Domingo E. Viral genetic variation: hepatitis B virus as a clinical example. Lancet 1993; 341: 349-53.
6. Chen DS, Kuo G, Sung JL, Lai MY, Sheu JC, Chen PJ, Hahn LC, Choo QL, Wang TH, Houghton M. Hepatitis C virus infection in an area hyperendemic for hepatitis B and chronic liver diseases: the Taiwan experience. J Infect Dis 1990; 162: 817-22.
7. Chen DS, Lai MY, Lee SC, Yang PM, Sheu JC, Sung JL. Serum HBsAg, HBeAg, anti-HBe, and hepatitis B viral DNA in asymptomatic carriers in Taiwan. J Med Virol 1986; 19: 87-94. 8. Chen DS, Sung JL and Lai MY. A seroepidemiological study of hepatitis B virus infection in Taiwan. J Formosan Med Assoc 1978; 77: 908-18.
9. Chen DS. From hepatitis to hepatoma: lessons from type B viral hepatitis. Science 1993; 262: 369-370.
10. Chen PJ, Chen DS, Chen CR, Chen YY, Chen Hsu HM, Lai MY, Sung JL. δ infection in asymptomatic carriers of hepatitis B surface antigen: low prevalence of δ activity and effective suppression of hepatitis B virus replication. Hepatology 1988; 8: 1121-4.
11. Chen PJ, Lin MH, Tai KF, Liu PC, Lin CJ, Chen DS. The Taiwanese hepatitis C virus genome: sequence determination and mapping the 5' termini of viral genomic and antigenomic RNA. Virology 1992; 188: 102-13.
12. Chesebro B, Wehrly K, Nishio J, Perryman S. Macrophage-tropic human immunodeficiency virus isolates from different patients exhibit unusual V3 envelope sequence homogeneity in comparison with T-cell-tropic isolates: definition of critical amino acids involved in cell tropism. J Virol 1992; 66: 6547-54.
13. Chien DY, Choo QL, Ralston R, Spaete R, Tong M, Houghton M, Kuo G. Persistence of HCV despite antibodies to both putative envelope glycoproteins. Lancet 1993; 342: 933 [letter]. 14. Choo QL, Kuo G, Weiner AJ, et al. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 1989; 244: 359-62.
Medina-Selby A, Barr PJ, Weiner AJ, Bradley DW, Kuo G, Houghton M. Genetic organization and diversity of the hepatitis C virus. Proc Natl Acad Sci USA 1991; 88: 2451-5.
16. Chuang WL, Omata M, Ehata T, et al. Precore mutations and core clustering mutations in chronic hepatitis B virus infection. Gastroenterology 1993; 104: 263-71.
17. Cristlano K, Di Bisceglie AM, Hoofnagle JH, Feinstone SM. Hepatitis C viral RNA in serum of patients with chronic non-A, non-B hepatitis: Detection by the polymerase chain reaction using multiple primer sets. Hepatology 1991; 14: 51-5.
18. Davis GL, Hoofnagle JH, Waggoner JG. Acute type A hepatitis during chronic hepatitis B virus infection: association of depressed hepatitis B virus replication with appearance of endogenous alpha interferon. J Med Virol 1984;14:141-7.
19. Dienstag JL and Alter HJ. Non-A, non-B hepatitis. Sem Liver Dis 1986; 6: 67-81.
20. Domingo E, Martinez-Salas E, Sobrino F, et al. The quasispecies (extremely heterogenous) nature of viral RNA genome population: biological relevance - a review. Gene 1985; 40: 1-8. 21. Enomoto N, Sakamoto N, Kurosaki M, Marumo F, Sato C. The hypervariable region of the HCV genome changes sequentially during the progression of acute HCV infection to chronic hepatitis. J Hepatol 1993; 17: 415-22.
22. Farci P, Alter HJ, Govindarajan S, et al. Lack of protective immunity against reinfection with hepatitis C virus. Science 1992; 258: 135-40.
23. Feray C, Samuel D, Thiers V, et al. Reinfection of liver graft by hepatitis C virus after liver transplantation. J Clin Invest 1992; 89: 1361-5.
24. Fong TL, Shindo M, Feinstone SM, et al. Detection of replicative intermediates of hepatitis C viral RNA in liver and serum of patients with chronic hepatitis C. J Clin Invest 1991; 88: 1058-60.
25. Gil B, Qian C, Riezu-Boj JI, Civeira MP, Prieto J. Hepatic and extrahepatic HCV RNA strands in chronic hepatitis C: different patterns of response to interferon treatment. Hepatology 1993; 18: 1050-4.
26. Hagiwara H, Hayashi N, Mita E, Naito M, Kasahara A, Fusamoto H, Kamada T. Quantitation of hepatitis C virus RNA in serum of asymptomatic blood donors and patients with type C chronic liver disease. Hepatology 1993; 17: 545-50.
27. Higashi Y, Kakumu S, Yoshioka K, Wakita T, Mizokami M, Ohba K, Ishikawa YT, Takayanagi M, Nagai Y. Dynamics of genome change in the E2/NS1 region of hepatitis C virus in vivo. Virology 1993; 197: 659-68.
28. Hijikata M, Kato N, Ootsuyama Y, Nakagawa M, Ohkoshi S, Shimotohno K. Hypervariable regions in the putative glycoprotein of hepatitis C virus. Biochem Biophys Res Commun 1991; 175: 220-8.
29. Holland J, Spindler K, Horodyski F, et al. Rapid evolution of RNA genomes. Science 1982; 215: 1577-85.
sequence evolution in the surface envelope glycoprotein of human immunodeficiency virus type 1 within a single infected patient. Proc Natl Acad Sci USA 1992; 89: 4835-9.
31. Houghton M, Weiner A, Han J, Kuo G, Choo QL. Molecular biology of the hepatitis C viruses: implications for diagnosis, development and control of viral disease. Hepatology 1991; 14: 381-8.
32. Hwang SS, Boyle TJ, Lyerly HK, Cullen BR. Identification of envelope V3 loop as the major determinant of CD4 neutralization sensitivity of HIV-1. Science 1992; 257: 535-7.
33. Hwang SS, Boyle TJ, Lyerly HK, Cullen BR. Identification of the envelope V3 loop as the primary determinant of cell tropism in HIV-1. Science 1991; 253: 71-4.
34. Kao JH, Chen PJ, Lai MY, Chen DS. Superinfection of heterologous hepatitis C virus in a patient with chronic type C hepatitis. Gastroenterology 1993; 105: 583-7.
35. Kao JH, Chen PJ, Lai MY, Wang TH, Chen DS. Sexual transmission of HCV. Lancet 1993; 342: 626.
35a. Kao JH, Chen PJ, Lai MY, Wang TH, Chen DS. Quasispecies of Hepatitis C Virus and Genetic Drift of the Hypervariable Region in Chronic Type C Hepatitis. J Infect Dis (in press) 36. Kao JH, Chen PJ, Lai MY, Yang PM, Sheu JC, Wang TH, Chen DS. Detection of divergent hepatitis C virus envelope sequences. J Biomed Sci 1994; 1: 158-62.
37. Kao JH, Chen PJ, Lai MY, Yang PM, Sheu JC, Wang TH, Chen DS. Mixed infections of hepatitis C virus as a factor in acute exacerbation of chronic type C hepatitis. J Infect Dis 1994; 170: 1128-33.
38. Kao JH, Chen PJ, Yang PM, Lai MY, Sheu JC, Wang TH, Chen DS. Intrafamilial transmission of hepatitis C virus: the important role of infections between spouses. J Infect Dis 1992; 166: 900-3.
39. Kao JH, Tsai SL, Yang PM, Sheu JC, Lai MY, Hsu HC, Sung JL, Wang TH, Chen DS. A clinicopathologic study of chronic non-A, non-B (type C) hepatitis in Taiwan: comparison between posttransfusion and sporadic patients. J Hepatol 1994; 21: 244-9.
40. Kato N, Hijikata M, Ootsuyama Y, Nakagawa M, Ohkoshi S, Sugiura T, Shimotohno K. Molecular cloning of the human hepatitis C virus genome from Japanese patients with non-A, non-B hepatitis. Proc Natl Acad Sci USA 1990; 87: 9524-8.
41. Keys B, Karis J, Fadeel B, Valentin A, Norkrans G, Hagberg L, Chiodi F. V3 sequences of paired HIV-1 isolates from blood and cerebrospinal fluid cluster according to host and show variation related to the clinical stage of disease. Virology 1993; 196: 475-483.
42. Kiyosawa K, Sodeyama T, Tanaka E, et al. Interrelationship of blood transfusion, non-A, non-B hepatitis and hepatocellular carcinoma: Analysis by detection of antibody to hepatitis C virus. Hepatology 1990; 12: 671-5.
43. Kumar U, Brown J, Monjardino J, Thomas HC. Sequence variation in the large envelope glycoprotein (E2/NS1) of hepatitis C virus during chronic infection. J Infect Dis 1993; 167: 726-30.
44. Kuo G, Choo QL, Alter HJ, Gitnick GI, Redeker AG, Purcell RH, Miyamura T, Dienstag JL, Alter MJ, Stevens CE, Tegtmeier GE, Overby LR, Bradley DW, Houghton M. An assay for circulating antibodies to a major etiologic virus of human non-A, non-B hepatitis. Science 1989; 244: 362-4.
45. Kuo MYP, Hahn LJ, Hong CY, Kao JH and Chen DS. Low prevalence of hepatitis C virus infection among dentists in Taiwan. J Med Virol 1993; 40: 10-3.
46. Kurosaki M, Enomoto N, Sato C, et al. Correlation of plasma hepatitis C cirus RNA levels with serum alanine aminotransferase in non-A, non-B chronic liver disease. J Med Virol 1993; 39: 246-50.
47. Lai ME, Mazzoleni AP, Argiolu F, De Virgilis S, Balestrieri A, Purcell RH, Cao A, Farci P. Hepatitis C virus in multiple episodes of acute hepatitis in polytransfused thalassaemic children. Lancet 1994; 343: 388-90.
48. Lai MMC, Lee CM, Bih FY, Govindarajan S. The molecular basis of heterogeneity of hepatitis delta virus. J Hepatol 1991; 13: S121-4.
49. Lai MY, Chen DS, Lee SC, Su IJ, Yang PM, Hsu HC, Sung JL. Reactivation of hepatitis B virus in anti-HBe-positive chronic active type B hepatitis: molecular and immunohistochemical studies. Hepatogastroenterology 1988; 35: 17-21.
50. Lau JY, Davis GL, Kniffen J, et al. Significance of serum hepatitis C virus RNA levels in chronic hepatitis C. Lancet 1993; 341: 1501-4.
51. Lee CM, Bih FY, Chao YC, et al. Evolution of hepatitis delta virus RNA during chronic infection. Virology 1992; 188: 265-73.
52. Lee CM, Bih FY, Chao YC, Govindarajan S, Lai MMC. Evolution of hepatitis delta virus RNA during chronic infection. Virology 1992; 188: 265-73.
53. Lin HH, Kao JH, Hsu HY, Ni YH, Yeh SH, Hwang LH, Chang MH, Hwang SC, Chen PJ, Chen DS. Possible role of high-titer maternal viremia in perinatal transmission of hepatitis C vorus. J Infect Dis 1994; 169: 638-41.
54. Lin HH, Kao JH, Leu JH, Young YC, Lee TY, Chen PJ and Chen DS. Comparison of three different immunoassays and PCR for the detection of hepatitis C virus infection in pregnant women in Taiwan. Vox Sang 1993; 65: 117-21.
55. Lok ASF, Lai CL. Acute exacerbations in Chinese patients with chronic hepatitis B virus (HBV) infection. Incidence, predisposing factors and etiology. J Hepatol 1990; 10: 29-34.
56. Martell M, Esteban JI, Quer J, Genesca J, Weiner A, Esteban R, Guardia J, Gómez J. Hepatitis C virus (HCV) circulates as a population of different but closely related genomes: quasispecies nature of HCV genome distribution. J Virol 1992; 66: 3225-9.
57. Matloubian M, Somasundaram S, Kolhekar SR, Selvakumar R, Ahmed R. Genetic basis of viral persistence: single amino acid change in the viral glycoprotein affects ability of lymphocytic choriomeningitis virus to persist in adult mice. J Exp Med 1990; 172: 1043-8. 58. Matsuura Y, Harada S, Suzuke R, Watanabe Y, Inoue Y, Saito I, Miyamura T. Expression of
processed envelope protein of hepatitis C virus in mammalian and insect cells. J Virol 1992; 66: 1425-31.
59. McGuiness PH, Bishop GA, McCaughan GW, Trowbrige R, Gowans EJ. False detection of negative-strand hepatitis C virus RNA. Lancet 1994; 343: 551-2.
60. Milich L, Margolin B, Swanstrom R. V3 loop of the human immunodeficiency virus type 1 Env protein: interpreting sequence variability. J Virol 1993; 67: 5623-34.
61. Muller HM, Pfaff E, Goeser T, Kallinowski B, Solbach C, Theilmann L. Peripheral blood leukocytes serve as a possible extrahepatic site for hepatitis C virus replication. J Gen Virol 1993; 74: 669-76.
62. Ogata N, Alter HJ, Miller RH, Purcell RH. Nucleotide sequence and mutation rate of the H strain of hepatitis C virus. Proc Natl Acad Sci USA 1991; 88: 3392-6.
63. Okamoto H, Kojima M, Okada SI, Yoshizawa H, Iizuka H, Tanaka T, Muchmore EE, Peterson DA, Ito Y, Mishiro S. Genetic drift of hepatitis C virus during an 8.2-year infection in a chimpanzee: variability and stability. Virology 1992; 190: 894-9.
64. Okamoto H, Kurai K, Okada S, et al. Full-length sequence of a hepatitis C virus genome having poor homology to reported isolates: comparative study of four distinct genotypes. Virology 1992; 188: 331-41.
65. Okamoto H, Mishiro S, Tokita H, Tsuda F, Miyakawa Y, Mayumi M. Superinfection of chimpanzees carrying hepatitis C virus of genotype II/1b with that of genotype III/2a or I/1a. Hepatology 1994; 20: 1131-6.
66. Okamoto H, Okada S, Sugiyama Y, et al. Detection of hepatitis C virus RNA by a two-stage polymerase chain reaction with two pairs of primers deduced from the 5'-noncoding region. Jpn J Exp Med 1990; 60: 215-22.
67. Okamoto H, Okada S, Sugiyama Y, et al. Nucleotide sequence of the genomic RNA of hepatitis C virus isolated from a human carrier: comparison with reported isolates for conserved and divergent regions. J Gen Virol 1991; 72: 2697-704.
68. Okuda H, Obata H, Motoike Y and Hisamitsu T. Clinicopathological features of hepatocellular carcinoma- Comparison of hepatitis B seropositive and seronegative patients. Hepatogastroenterology 1984; 31: 64-8.
69. Oldstone MBA. Molecular anatomy of viral persistence. J Virol 1991; 65: 6381-6.
70. Phillips RE, Rowland-Jones S, Nixon DF, et al. Human immunodeficiency virus genetic variation that can escape cytotoxic T cell recognition. Nature 1991; 354: 453-9.
71. Prince AM, Brotman B, Huima T, Pascual D, Jaffery M, Inchauspe G. Immunity in hepatitis C infection. J Infect Dis 1992; 165: 438-43.
72. Qian C, Camps J, Maluenda MD, Civeria MP, Prieto J. Replication of hepatitis C virus in peripheral blood mononuclear cells. J Hepatol 1992; 16: 380-3.
73. Sheen IS, Liaw YF, Chu CM, Pao CC. Role of hepatitis C virus infection in spontaneous hepatitis B surface antigen clearance during chronic hepatitis B virus infection. J Infect Dis 1992;
165: 831-4.
74. Shimizu Y, Purcell RH, Yoshikura H. Correlation between the infectivity of hepatitis C virus in vivo and its infectivity in vitro. Proc Natl Acad Sci USA 1993; 90: 6037-41.
75. Shimizu Y, Iwamoto A, Hijikata M, Purcell RH, Yoshikura H. Evidence for in vitro replication of hepatitis C virus genome in a human T cell line. Proc Natl Acad Sci USA 1992; 89: 5477-81.
76. Simmonds P, Holmes EC, Cha TA, Chan SW, McOmish F, Irvine B, Beall E, Yap PL, Kolberg J, Urdea MS. Classification of hepatitis C virus into six major genotypes and a series of subtypes by phylogenetic analysis of the NS-5 region. J Gen Virol 1993; 74: 2391-9.
77. Takamizawa A, Mori C, Fuke I, Manbe S, Murakami S, Fujita J, Onishi E, Andoh T, Yoshida I, Okayama H. Structure and organization of the hepatitis C virus genome isolated from human carriers. J Virol 1991; 65: 1105-13.
78. Tsai SL, Chen PJ, Hwang LH, Kao JH, Huang JH, Chang TH, Chen DS. Cellular immune response to hepatitis C virus (HCV) nonstructural protein in chronic HCV infection. J Hepatol 1994; 21: 403-11.
79. Wang JT, Sheu JC, Lin JT, Wang TH, Chen DS. Detection of replicative form of hepatitis C virus RNA in peripheral blood mononuclear cells. J Infect Dis 1992; 166: 1167-9.
80. Wang JT, Wang TH, Lin JT, Sheu JC, Sung JL, Chen DS. Hepatitis C virus in a prospective study of posttransfusion non-A, non-B hepatitis in Taiwan. J Med Virol 1990; 32: 83-6.
81. Wang JT, Wang TH, Sheu JC, Lin JT, Wang CY, Chen DS. Posttransfusion hepatitis revisited by hepatitis C antibody assays and polymerase chain reaction. Gastroenterology 1992; 103: 609-16.
82. Weiner AJ, Braner M, Rosenblatt J, Richman K, Tung J, Crawford K, Bonino F, Saracco G, Choo QL, Houghton M, Han JH. Variable and hypervariable domains are found in the regions of HCV corresponding to the flavivirus envelope and NS 1 proteins and the pestivirus envelope glycoproteins. Virology 1991; 180: 842-8.
83. Weiner AJ, Geysen HM, Christopherson C, Hall JE, Mason TJ, Saracco G, Bonino F, Crawford K, Marion CD, Crawford KA, Brunetto M, Barr PJ, Miyamura T, McHutchinson J, Houghton M. Evidence for immune selection of hepatitis C virus (HCV) putative envelope glycoprotein variants: potential role in chronic HCV infections. Proc Natl Acad Sci USA. 1992; 89: 3468-72.
84. Weiss RA. How does HIV cause AIDS? Science 1993; 260: 1273-8.
85. Yeh S. Some geographic aspects of common disease in Taiwan. Internatl Path 1966; 7: 24-8.
86. Yoshioka K, Kakumu S, Wakaji T, Ishikawa T, Itoh Y, Takayanagi M, Higashi Y, Shibata M, Morishima T. Detection of hepatitis C virus by polymerase chain reaction and response to interferon-α therapy: relationship to genotypes of hepatitis C virus. Hepatology 1992; 16: 293-9. 87. Zignego AL, Macchia D, Monti M, Thiers V, Mazzetti M, Foschi M, Maggi E, Romagnani
S, Gentilini P, Brechot C. Infection of peripheral mononuclear blood cells by hepatitis C virus. J Hepatol 1992; 15: 382-6.