Contribution of DNA Double-strand Break Repair Gene XRCC3 Genotypes to Oral Cancer Susceptibility in Taiwan
Chia-Wen Tsai1,3, Wen-Shin Chang2,3, Juhn-Cherng Liu2,3, Ming-Hsui Tsai3, Cheng-Chieh
Lin3,4,# and Da-Tian Bau1,2,3,#
Graduate Institutes of 1Basic Medical Science and 2Clinic Medical Science, China Medical
University; 3Terry Fox Cancer Research Lab in China Medical University Hospital; 4Asia
University, Taichung, Taiwan, R.O.C.
# the last two are joint corresponding authors
Correspondence to: Cheng-Chieh Lin and Da-Tian Bau, Terry Fox Cancer Research Lab, Department of Medical Research, China Medical University Hospital, 2 Yuh-Der Road, Taichung, 404 Taiwan, R.O.C. Tel: +886422052121 Ext 7534, Fax: +886 422053366 Ext. 1511, e-mail: [email protected]; [email protected]
Abstract. The DNA repair gene XRCC3 is thought to play a major role in the double strand break repairing and genome stability maintaining. Very possibly, defective in double strand break repair capacity of the cells can lead to human carcinogenesis. Therefore, a case-control study was performed to reveal the contribution of XRCC3 genotypes to individual oral cancer susceptibility. In this hospital-based research, the association of XRCC3 rs1799794, rs45603942, rs861530, rs3212057, rs1799796, rs861539, rs28903081 genotypes with oral cancer risk in a Taiwanese population was investigated. Totally, 788 oral cancer patients and 956 age- and gender-matched healthy controls were genotyped and analyzed by PCR-RFLP method. The results showed that there were significant differential distribution among oral cancer and control subjects in the genotypic (P=0.001428) and allelic (P=0.0013) frequencies of XRCC3 rs861539. As for the other polymorphisms, there was no difference between the case and control groups. In gene-environment interaction analysis, we have firstly provided evidence showing that there is an obvious joint effect of
XRCC3 rs861539 genotype with individual areca chewing habits on oral risk. In
conclusion, the T allele of XRCC3 rs861539, which has an interaction with areca chewing habit in oral carcinogenesis, may be an early detection marker for oral cancer in Taiwan.
Oral cancer, which is the tenth most commonly diagnosed cancer all over the world (1), has the highest incidence of all head and neck cancers in Taiwan (2). Three major environmental factors, tobacco, alcohol and betel nuts, are main causes of oral cancer in Taiwan, while the genomic etiology of oral cancer is of great interest but largely unknown. Human DNA repair mechanisms protect the genome from various insults caused by endogenous and exogenous DNA damaging agents (3) and defects in the DNA repairing system are thought to be essential for tumorigenesis (4, 5). Therefore, it is logical to suspect that some genetic variants of DNA repair genes might contribute to oral cancer pathogenesis.
Environmental carcinogens such as UV light, ionizing radiation or chemical agents contained for instance in the tobacco smoke may induce double strand breaks (DSBs). DSBs are a severe type of DNA damage which should be repaired by the DNA DSB repair system (6, 7). If cells cannot remove them immediately by means of homologous recombination (HR) and nonhomologous end-joining (NHEJ), those DNA DSBs may induce precancerous lesions and cancer itself as well (8, 9). Genetic polymorphisms in DNA DSB repair genes influence DNA repair capacity and confer predisposition to several cancers, including skin (10), breast (11, 12) liver (13), gastric (14), and oral cancers (15, 16).The X-ray repair cross-complementing group 3 (XRCC3; 14q32.3) is a member of the rad51 DNA repair family, which has been shown to interact directly with rad51 and is essential with respect to the proper accumulation of rad51 at sites of DNA DSBs in the
nucleus (17).
The most common genetic polymorphism on XRCC3 gene is the rs861539 C/T polymorphism (also named Thr241Met, T241M, C18067T and C722T). Some studies were performed to investigate the association between XRCC3 rs861539 and oral premalignant (9) and oral cancer risk (18-22), however no consistent finding was reported. The inconsistency may be caused by small sample sizes and different genetic background among ethnics. To identify the contribution of XRCC3 genotyping to oral cancer risk in Taiwan, we determined the genotypic frequencies of up to seven polymorphisms of the
XRCC3 gene at promoter A-315G (rs1799794), promoter C-280T (rs45603942), intron5
(rs861530), exon6 (rs3212057), intron7 (rs1799796), exon8 (rs861539) and exon10 (rs28903081), and evaluate the gene-environment interaction.
Materials and Methods
Study population and sample collection. Seven hundred and eighty eight patients diagnosed
with oral cancer were recruited at the China Medical University Hospital in central Taiwan during 1998 to 2010. All patients voluntarily participated, completed a self-administered questionnaire and provided 5 ml of their peripheral blood samples. The questionnaire administered to the subjects included questions on history and frequency of alcohol consumption, areca chewing and smoking habits. Self-reported alcohol consumption, areca chewing and smoking habits were evaluated and classified as categorical variables. Information on these factors was obtained as more then twice a week for years as ‘‘ever’’.
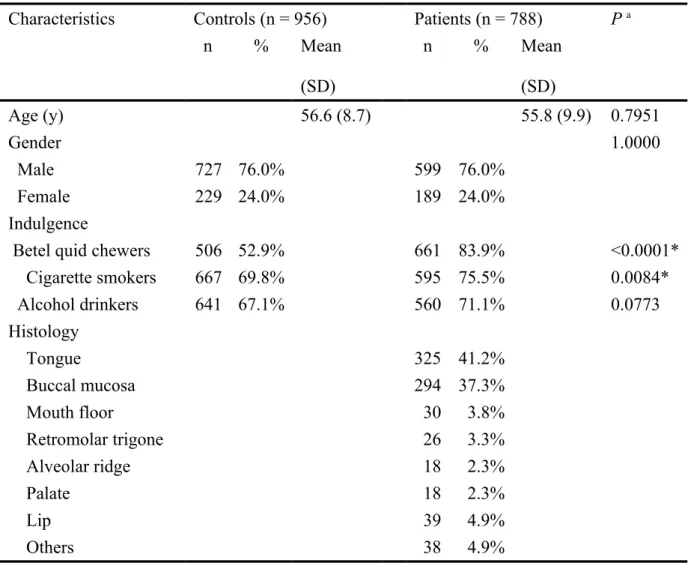
The 956 non-cancer healthy people as controls were selected by matching for age and gender after initial random sampling from the Health Examination Cohort of the hospital. The ratio of male versus female was both 76% versus 24% in each group. The mean age of the oral cancer patients and the controls were 55.8 (SD = 9.9) and 56.6 (SD = 8.7) years, respectively (see Table I for more details). Our continuous study was approved by the Institutional Review Board of the China Medical University Hospital and written-informed consents were obtained from all participants.
Genotyping conditions. Genomic DNA was prepared from peripheral blood leucocytes
using a QIAamp Blood Mini Kit (Blossom, Taipei, Taiwan) and further processed as our recent papers (23, 24). A total of seven polymorphic sites were analyzed in all the subjects in control and case groups. Briefly, all of the seven polymorphic sites were genotyped by means of a PCR-restriction fragment length polymorphism (PCR-RFLP). PCR was performed on BioRad’s Mycycler (BioRad, Hercules, CA, USA) following the normal manufacturer’s instructions. Each PCR reaction consisted of 5 min initial cycle at 94oC for
5 min; 40 cycles of 94oC for 30 s, 55oC for 30 s, and 72oC for 30 s; and a final extension at
72oC for 10 min. Then the SNP-containing DNA amplicons were subjected to individual
overnight digestion by restriction endonucleases following the manufacturer’s instructions (see Table II for more details). Following digestion, each sample was immediately analyzed by 2% agarose gel electrophoresis. Details such as the primer sequences, and enzymatic
digestion conditions for each SNP analyzed in this study were summarized in Table II.
Statistical analyses. The subjects matched with all SNPs data and clinical (case/control
=788/956) were taken into final analyzing. To ensure that the controls used were representative of the general population and to exclude the possibility of genotyping error, the deviation of the genotype frequencies of XRCC3 SNPs in the control subjects from those expected under the Hardy-Weinberg equilibrium was assessed using the goodness-of-fit test. Pearson’s chi-square test was used to compare the distribution of the XRCC3 genotypes between cases and controls. Cancer risk associated with the genotypes was estimated as odds ratio (ORs) and 95% confidence intervals (CIs) using unconditional logistic regression. Data was recognized as significant when the statistical P-value was less than 0.05.
Results
The clinical characteristics of recruited 788 oral cancer patients and 956 of age- and gender-matched controls are shown in Table I. Since the controls are age-, gender-matched with the cases, there was no significant difference between the two groups as in their age and gender (Table I). However as for the personal habits, there was significant difference that the case group seemed to have more smokers and betel quid chewers (Table I). The frequency distributions of the genotypes for the XRCC3 rs1799794, rs45603942, rs861530, rs3212057, rs1799796, rs861539 and rs28903081 polymorphic sites between controls and
oral cancer patients are shown in Table III. Genotypic distribution pattern of XRCC3 rs861539 was significantly different between oral cancer and control groups (P<0.05), while those for rs1799794, rs45603942, rs861530, rs3212057, rs1799796 and rs28903081 were not significant (P>0.05) (Table III). In detail, distributions of XRCC3 rs861539 CC homozygote/heterozygote/TT homozygote in controls and oral cancer patients were 91.9/7.6/0.5% and 86.8/11.7/1.5%, respectively (Table III). There was no heterozygote or homozygote variant for XRCC3 rs3212057 and rs28903081 among Taiwanese subjects (Table III). To sum up, the genotype of XRCC3 rs861539, not rs1799794, rs45603942, rs861530, rs3212057, rs1799796 or rs28903081, is associated with oral cancer risk and may be a biomarker for oral cancer early detection and prediction.
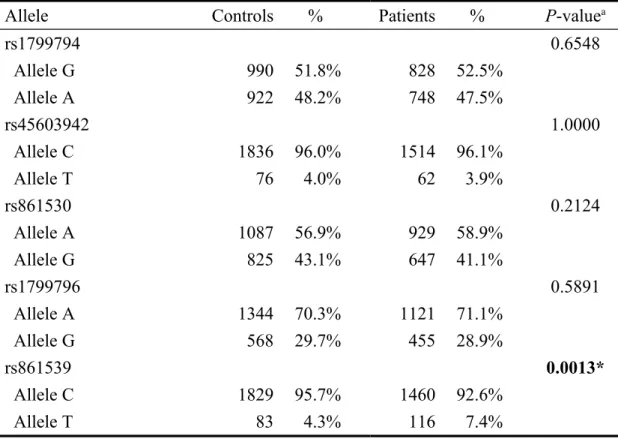
The frequencies of the alleles for the XRCC3 rs1799794, rs45603942, rs861530, rs3212057, rs1799796, rs861539 and rs28903081 of all the recruited subjects are shown in Table IV. Among them, the carriers of XRCC3 rs861539 allele T were of higher risk for oral cancer (P=0.0013), while genotypes of XRCC3 rs1799794, rs45603942, rs861530, rs3212057, rs1799796 and rs28903081 were not associated with oral cancer susceptibility (Table IV).
We were interested to investigate the potential gene-environment interactions between
XRCC3 gene and oral cancer-related individual habits. In Taiwan, the habits of smoking,
alcoholism and betel quid chewing are believed to significantly increase oral cancer risk. Therefore, the risk of oral cancer related to XRCC3 genotypes was further examined with
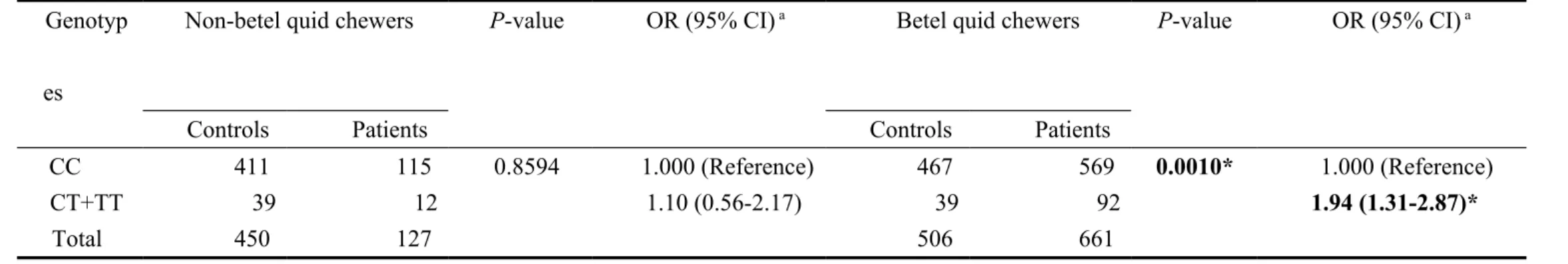
stratification by areca chewing, smoking and alcohol drinking status. Table V showed the interaction of XRCC3 genotype and betel quid chewing status on personal oral cancer susceptibility (Table V). The results showed that compared with C/C genotype, the C/T or T/T significantly enhanced the risk only in the areca chewers (P=0.0010, OR=1.94, 95%CI=1.31-2.87), not in the non-areca chewers (P>0.05, OR=1.10, 95%CI=0.56-2.17) (Table V). With the same strategy, the interactions among XRCC3 genotype and smoking or alcohol drinking statuses were also analyzed, but there were no significant interaction found (data not shown).
Discussion
In recent years, there were a few papers investigated the contribution of genetic variations on DSB repair genes to oral cancer risk (9, 15, 16, 18-22, 25-29). Among them, several studies had analyzed the interaction of genetic variations and behavioral factors on oral cancer (19, 20, 26, 28). The present study is to investigate the role of XRCC3 gene polymorphisms in oral cancer risk in Taiwan, where the oral cancer prevalence density in highest all over the world due to an over-exposure to environmental factors such as betel quid, smoke and alcohol. Among the seven polymorphisms of XRCC3, the one rs861539 located in the exon region and the T allele on it was associated with oral cancer in Taiwan (Table III and IV), but the other six polymorphisms were not. The rs861539 genetic variation direct result in an amino acid coding alteration from Thr to Met. Possibly, this
predisposing to oral carcinogenesis. Up to now, the studies investigating the association of
XRCC3 polymorphism with oral cancer risk did not reach a consistent conclusion. This may
be due to different recruited populations with different genetic background, various behavioral and environmental factors were taken into consideration, and inconsistent polymorphic sites were chosen. In 2012, XRCC3 rs3212057 was found to be associated with head and neck cancer in Poland (30). There were also some negative findings reporting no association between XRCC3 genotype and oral cancer in Brazil (18), Belgium (20), and India (19). As for XRCC3 rs861539, positive finding was reported in Thailand, but the sample size of the study was rather small with only 112 oral cancer cases and 119 controls (21).
Thus, the large enough sample size (control : case=956 : 788) and concise data analysis without adjustment strengthen the accuracy and reliability of our finding, and the frequencies of XRCC3 polymorphisms variant alleles were similar to those reported in the NCBI website in the Asian population studies, for example minor T allele frequencies of XRCC3 rs861539 is 4.3% (Table IV) in our control group and 4.7 to 11.0% for Asian population in NCBI. In 2005, in Jin and his colleagues’ work, the minor T allele frequencies of XRCC3 rs861539 is 0.36% in 280 controls and 0.71% in 140 colorectal cancer patients in Taiwan (31). The data suggested that no selection bias for the subject’s enrolments in terms of genotypes. Therefore the verifying of our findings in further larger studies is not so urgently needed.
There were three behavioral factors reported to be closely related to oral carcinogenesis in Taiwan, cigarette smoking, alcohol consumption and betel quid chewing. Previously, our group has provided evidence for the interactions between DNA double strand break genes and betel quid chewing habit, such as XRCC4, XRCC5 (15, 29). Again, the results in this study have shown that there was positive interaction of variant DNA double strand break gene XRCC3 rs861539 genotypes with betel quid chewing habits in oral cancer risk (Table V). People with betel quid chewing habit and carrying the T allele of
XRCC3 rs861539 have the higher risk of oral cancer among our stratified subgroups. These
findings strengthened the theory of oral carcinogenesis that genetic variants in DNA double strand break system may enhance the genomic vulnerability to areca chewing-caused DNA insults, leading to oral carcinogenesis.
In conclusion, we found the genotype of XRCC3 rs861539, but not those on rs1799794, rs45603942, rs861530, rs3212057, rs1799796 or rs28903081, was associated with higher oral cancer risk. In addition, the increasing of oral cancer risk by variant genotypes of XRCC3 rs861539 was more obviously enhanced among betel quid chewers, but not among none-chewers. Individual smoking and alcohol drinking habits could not enhance the risky genotype in increasing oral cancer susceptibility. The XRCC3 rs861539 polymorphism might become a potential biomarker for the oral oncology early detection and prediction and further investigation of the phenotypic effects determined by this genotypic variation on oral carcinogenesis are needed.
Acknowledgements
We thank Tsai-Ping Ho, Chieh-Lun Hsiao, Lin-Lin Hou, Chia-En Miao, Tzu-Chia Wang, Yun-Ru Syu and Tissue-bank of China Medical University Hospital for technical assistance. This study was supported by research grants from Terry Fox Cancer Research Foundation of China Medical University and the National Science Council (NSC101-2320-B-039-045 and NSC102-2320-(NSC101-2320-B-039-045).
References
1 Jemal A, Bray F, Center MM, Ferlay J, Ward E and Forman D: Global cancer statistics. CA Cancer J Clin 61: 69-90, 2011.
2 Department of Health, Taiwan: Cancer registration system annual report. Taiwan,
Department of Health; 2012.
3 Sugimura T, Kumimoto H, Tohnai I, Fukui T, Matsuo K, Tsurusako S, Mitsudo K, Ueda M, Tajima K and Ishizaki K: Gene-environment interaction involved in oral carcinogenesis: molecular epidemiological study for metabolic and DNA repair gene polymorphisms. J Oral Pathol Med 35: 11-18, 2006.
4 Vogelstein B, Alberts B and Shine K: Genetics. Please don't call it cloning! Science
295: 1237, 2002.
5 Miller KL, Karagas MR, Kraft P, Hunter DJ, Catalano PJ, Byler SH and Nelson HH: XPA, haplotypes, and risk of basal and squamous cell carcinoma. Carcinogenesis 27: 1670-1675, 2006.
6 Wood RD, Mitchell M, Sgouros J and Lindahl T: Human DNA repair genes. Science 291: 1284-1289, 2001.
7 Yu Z, Chen J, Ford BN, Brackley ME and Glickman BW: Human DNA repair systems: an overview. Environ Mol Mutagen 33: 3-20, 1999.
8 Khanna KK and Jackson SP: DNA double-strand breaks: signaling, repair and the cancer connection. Nat Genet 27: 247-254, 2001.
9 Yang H, Lippman SM, Huang M, Jack Lee J, Wang W, Spitz MR and Wu X: Genetic polymorphisms in double-strand break DNA repair genes associated with
risk of oral premalignant lesions. Eur J Cancer 44: 1603-1611, 2008.
10 Han J, Colditz GA, Samson LD and Hunter DJ: Polymorphisms in DNA double-strand break repair genes and skin cancer risk. Cancer Res 64: 3009-3013, 2004. 11 Bau DT, Fu YP, Chen ST, Cheng TC, Yu JC, Wu PE and Shen CY: Breast cancer
risk and the DNA double-strand break joining capacity of nonhomologous end-joining genes are affected by BRCA1. Cancer Res 64: 5013-5019, 2004.
12 Bau DT, Mau YC, Ding SL, Wu PE and Shen CY: DNA double-strand break repair capacity and risk of breast cancer. Carcinogenesis 28: 1726-1730, 2007.
13 Hsu CM, Yang MD, Chang WS, Jeng LB, Lee MH, Lu MC, Chang SC, Tsai CW, Tsai Y, Tsai FJ and Bau DT: The contribution of XRCC6/Ku70 to hepatocellular carcinoma in Taiwan. Anticancer Res 33: 529-535, 2013.
14 Chiu CF, Wang CH, Wang CL, Lin CC, Hsu NY, Weng JR and Bau DT: A novel single nucleotide polymorphism in XRCC4 gene is associated with gastric cancer susceptibility in Taiwan. Ann Surg Oncol 15: 514-518, 2008.
15 Chiu CF, Tsai MH, Tseng HC, Wang CL, Wang CH, Wu CN, Lin CC and Bau DT: A novel single nucleotide polymorphism in XRCC4 gene is associated with oral cancer susceptibility in Taiwanese patients. Oral Oncol 44: 898-902, 2008.
16 Bau DT, Tseng HC, Wang CH, Chiu CF, Hua CH, Wu CN, Liang SY, Wang CL, Tsai CW and Tsai MH: Oral cancer and genetic polymorphism of DNA double strand break gene Ku70 in Taiwan. Oral Oncol 44: 1047-1051, 2008.
125-135, 2005.
18 Dos Reis MB, Losi-Guembarovski R, de Souza Fonseca Ribeiro EM, Cavalli IJ, Morita MC, Ramos GH, de Oliveira BV, Mizuno LT, Rogatto SR and de Syllos Colus IM: Allelic variants of XRCC1 and XRCC3 repair genes and susceptibility of oral cancer in Brazilian patients. J Oral Pathol Med 42: 180-185, 2013.
19 Majumder M, Sikdar N, Paul RR and Roy B: Increased risk of oral leukoplakia and cancer among mixed tobacco users carrying XRCC1 variant haplotypes and cancer among smokers carrying two risk genotypes: one on each of two loci, GSTM3 and XRCC1 (Codon 280). Cancer Epidemiol Biomarkers Prev 14: 2106-2112, 2005. 20 Werbrouck J, De Ruyck K, Duprez F, Van Eijkeren M, Rietzschel E, Bekaert S,
Vral A, De Neve W and Thierens H: Single-nucleotide polymorphisms in DNA double-strand break repair genes: association with head and neck cancer and interaction with tobacco use and alcohol consumption. Mutat Res 656: 74-81, 2008. 21 Kietthubthew S, Sriplung H, Au WW and Ishida T: Polymorphism in DNA repair
genes and oral squamous cell carcinoma in Thailand. Int J Hyg Environ Health 209: 21-29, 2006.
22 Yen CY, Liu SY, Chen CH, Tseng HF, Chuang LY, Yang CH, Lin YC, Wen CH, Chiang WF, Ho CH, Chen HC, Wang ST, Lin CW and Chang HW: Combinational polymorphisms of four DNA repair genes XRCC1, XRCC2, XRCC3, and XRCC4 and their association with oral cancer in Taiwan. J Oral Pathol Med 37: 271-277, 2008.
23 Chang WS, Tsai CW, Ji HX, Wu HC, Chang YT, Lien CS, Liao WL, Shen WC, Tsai CH and Bau DT: Associations of cyclooxygenase 2 polymorphic genotypes with bladder cancer risk in Taiwan. Anticancer Res 33: 5401-5405, 2013.
24 Hsia TC, Tsai CW, Liang SJ, Chang WS, Lin LY, Chen WC, Tu CY, Tsai CH and Bau DT: Effects of ataxia telangiectasia mutated (ATM) genotypes and smoking habits on lung cancer risk in Taiwan. Anticancer Res 33: 4067-4071, 2013.
25 Werbrouck J, De Ruyck K, Duprez F, Veldeman L, Claes K, Van Eijkeren M, Boterberg T, Willems P, Vral A, De Neve W and Thierens H: Acute normal tissue reactions in head-and-neck cancer patients treated with IMRT: influence of dose and association with genetic polymorphisms in DNA DSB repair genes. Int J Radiat Oncol Biol Phys 73: 1187-1195, 2009.
26 Matullo G, Dunning AM, Guarrera S, Baynes C, Polidoro S, Garte S, Autrup H, Malaveille C, Peluso M, Airoldi L, Veglia F, Gormally E, Hoek G, Krzyzanowski M, Overvad K, Raaschou-Nielsen O, Clavel-Chapelon F, Linseisen J, Boeing H, Trichopoulou A, Palli D, Krogh V, Tumino R, Panico S, Bueno-De-Mesquita HB, Peeters PH, Lund E, Pera G, Martinez C, Dorronsoro M, Barricarte A, Tormo MJ, Quiros JR, Day NE, Key TJ, Saracci R, Kaaks R, Riboli E and Vineis P: DNA repair polymorphisms and cancer risk in non-smokers in a cohort study. Carcinogenesis 27: 997-1007, 2006.
patients. Laryngoscope 115: 2221-2231, 2005.
28 Mondal P, Datta S, Maiti GP, Baral A, Jha GN, Panda CK, Chowdhury S, Ghosh S, Roy B and Roychoudhury S: Comprehensive SNP scan of DNA repair and DNA damage response genes reveal multiple susceptibility loci conferring risk to tobacco associated leukoplakia and oral cancer. PLoS One 8: e56952, 2013.
29 Hsu CF, Tseng HC, Chiu CF, Liang SY, Tsai CW, Tsai MH and Bau DT: Association between DNA double strand break gene Ku80 polymorphisms and oral cancer susceptibility. Oral Oncol 45: 789-793, 2009.
30 Gresner P, Gromadzinska J, Polanska K, Twardowska E, Jurewicz J and Wasowicz W: Genetic variability of Xrcc3 and Rad51 modulates the risk of head and neck cancer. Gene 504: 166-174, 2012.
31 Jin MJ, Chen K, Song L, Fan CH, Chen Q, Zhu YM, Ma XY and Yao KY: The association of the DNA repair gene XRCC3 Thr241Met polymorphism with susceptibility to colorectal cancer in a Chinese population. Cancer Genet Cytogenet
Table I. Summary of the characteristics for all the oral cancer patients and healthy controls. Characteristics Controls (n = 956) Patients (n = 788) P a
n % Mean (SD) n % Mean (SD) Age (y) 56.6 (8.7) 55.8 (9.9) 0.7951 Gender 1.0000 Male 727 76.0% 599 76.0% Female 229 24.0% 189 24.0% Indulgence
Betel quid chewers 506 52.9% 661 83.9% <0.0001*
Cigarette smokers 667 69.8% 595 75.5% 0.0084* Alcohol drinkers 641 67.1% 560 71.1% 0.0773 Histology Tongue 325 41.2% Buccal mucosa 294 37.3% Mouth floor 30 3.8% Retromolar trigone 26 3.3% Alveolar ridge 18 2.3% Palate 18 2.3% Lip 39 4.9% Others 38 4.9%
Table II. Summary of the rs numbers, primers, amplicon length before and after enzyme digestion, restriction enzymes for all the XRCC3 SNPs investigated in this study.
rs number of XRCC3 SNP Primer sequence Restriction enzyme Amplicon length Genotypes and
enzymatic fragment sizes rs1799794 F: 5’-CACACTGCGGTCTTGCAGTG-3’ R: 5’-CAGGCTGGGTCTGGATACAA-3’ BtsCI 505 bp G: 505 bp A: 289 + 216 bp rs45603942 F: 5’-GGGATGCAGGTTCAACTGAC-3’ R: 5’-AACTTGGACTGTGTCAAGCA-3’ AluI 352 bp C: 352 bp T: 187 + 165 bp rs861530 F: 5’-CCGAGGAACGTGCTGAACTT-3’ R: 5’-CTCCCTAACAGCCTCCATGT-3’ FatI 497 bp G: 497 bp A: 293 + 204 bp rs3212057 F: 5’-CCATGACCGCAGGCACTTGT-3’ R: 5’-AGAACGCGACAAGGATGGTA-3’ HpyCH4III 455 bp G: 455 bp A: 235 + 220 bp rs1799796 F: 5’-GG AACCAGTTGT GTGAGCCT-3’ R: 5’-CCTGGTTGATGCACAGCACA-3’ AluI 430 bp G: 430 bp A: 226 + 204 bp
R: 5’-GTCTTCTCGATGGTTAGGCA-3’ T: 200 + 158 bp
rs28903081 F: 5’-CTGCTTCCTGTTTCTCAGGT-3’
R: 5’-GCACTGATCGTGTAGGAACA-3’
BstUI 198 bp A: 198 bp
Table III. Distribution of XRCC3 genotypes among oral cancer patients and controls Genotype Controls (n) % Patients (n) % P-valuea Odds ratio (95% CI)b rs1799794 0.8932 GG 229 24.0% 195 24.7% 1.00 (Reference) AG 532 55.6% 438 55.6% 0.97 (0.77-1.22) AA 195 20.4% 155 19.7% 0.93 (0.70-1.24) rs45603942 0.9967 CC 886 92.7% 731 92.8% 1.00 (Reference) CT 64 6.7% 52 6.6% 0.98 (0.67-1.44) TT 6 0.6% 5 0.6% 1.01 (0.31-3.32) rs861530 0.3101 AA 284 29.7% 247 31.3% 1.00 (Reference) AG 519 54.3% 435 55.2% 0.96 (0.78-1.19) GG 153 16.0% 106 13.5% 0.80 (0.59-1.08) rs3212057 1.0000 GG 956 100.0% 788 100.0% 1.00 (Reference) AG 0 0.0% 0 0.0% 1.00 AA 0 0.0% 0 0.0% 1.00 rs1799796 0.8184 AA 438 45.8% 370 47.0% 1.00 (Reference) AG 468 49.0% 381 48.3% 0.96 (0.79-1.17) GG 50 5.2% 37 4.7% 0.88 (0.56-1.37) rs861539 0.001428* CC 878 91.9% 684 86.8% 1.00 (Reference) CT 73 7.6% 92 11.7% 1.62 (1.17-2.23)* TT 5 0.5% 12 1.5% 3.08 (1.08-8.79)* rs28903081 1.0000 GG 956 100.0% 788 100.0% 1.00 (Reference) AG 0 0.0% 0 0.0% 1.00 AA 0 0.0% 0 0.0% 1.00
aP-value based on chi-square test.
b CI: confidence interval;
Table IV. Distribution of XRCC3 alleles among oral cancer patients and controls
Allele Controls % Patients % P-valuea
rs1799794 0.6548 Allele G 990 51.8% 828 52.5% Allele A 922 48.2% 748 47.5% rs45603942 1.0000 Allele C 1836 96.0% 1514 96.1% Allele T 76 4.0% 62 3.9% rs861530 0.2124 Allele A 1087 56.9% 929 58.9% Allele G 825 43.1% 647 41.1% rs1799796 0.5891 Allele A 1344 70.3% 1121 71.1% Allele G 568 29.7% 455 28.9% rs861539 0.0013* Allele C 1829 95.7% 1460 92.6% Allele T 83 4.3% 116 7.4%
aP based on chi-square test.
Table V. Odds ratios for XRCC3 rs861539 genotype and oral cancer after stratified by areca chewing status Genotyp
es
Non-betel quid chewers P-value OR (95% CI) a Betel quid chewers P-value OR (95% CI) a
Controls Patients Controls Patients
CC 411 115 0.8594 1.000 (Reference) 467 569 0.0010* 1.000 (Reference)
CT+TT 39 12 1.10 (0.56-2.17) 39 92 1.94 (1.31-2.87)*
Total 450 127 506 661
a OR: Odds ratio, CI: confidence interval; ORs were estimated with multivariate logistic regression analysis.