PEITC inhibits human brain glioblastoma GBM 8401 cell migration
and invasion through the inhibition of uPA, Rock1, and Ras with
inhibition of MMP-2, -7 and -9 gene expression
Yu-Cheng Chou1,2,3,4, Meng-Ya Chang2,*, Mei-Jen Wang2, Fu-Shun Yu5, Hsin-Chung Liu6, Tomor Harnod7, Chih-Huang Hung2, Hsu-Tung Lee1,8, Jing-Gung Chung6,9,*
1Division of Neurosurgical Oncology, Neurological Institute, Taichung Veterans General Hospital, Taichung 407, Taiwan
2Institute of Medical Sciences, Tzu Chi University, Hualien 970, Taiwan 3School of Medicine, National Defense Medical Center, Taipei 114, Taiwan
4Rong Hsing Research Center for Translational Medicine, National Chung Hsing University
5Department of Dentist, China Medical University, Taichung 404, Taiwan
6Departments of Biological Science and Technology, China Medical University, Taichung 404, Taiwan
7Department of Neurosurgery, Hualien Tzu Chi Hospital, Buddhist Tzu Chi Medical Foundation and College of Medicine, Tzu Chi University, Hualien 970, Taiwan 8Graduate Institute of Medical Sciences, National Defense Medical Center, Taipei 114, Taiwan
9Department of Biotechnology, Asia University, Taichung 413, Taiwan.
Running title: PEITC inhibits migration and invasion in GBM-8401
*Both have equal contribution
*Correspondence to: Jing-Gung Chung, Department of Biological Science and Technology, China Medical University, No 91, Hsueh-Shih Road, Taichung 404, Taiwan. Tel: +886 4 2205 3366 ext 8000, Fax: +886 4 2205 3764, e-mail: [email protected]
Abstract. Glioblastoma is the most aggressive primary brain malignancy, and the multimodality treatments are still not completely satisfying. Phenethyl isothiocyanate (PEITC), one member of the isothiocyanate family, can inhibit migration and invasion of many human cancer cells. PEITC induces apoptosis of human brain glioblastoma GBM 8401 cells through the extrinsic- and intrinsic-signaling pathways from our previous study. In this study, we are the first to investigate the effects of PETIC on migration and invasion of GBM 8401 cells. Our results indicated the effects of PEITC on migration of GBM 8401 cells from scratch wound healing assay and Transwell migration assay were in a dose-dependent manner. The inhibition percentage of Transwell migration assay ranged from 46.89% to 15.75%, and invasion assay ranged from 27.80% to 7.31% after 48 h-treatment of PEITC. Results from Western blotting analysis indicated that PEITC decreased the levels of proteins associated with migration and invasion, Ras, uPA, Rho A, GRB2, p-p38, p-JNK, p-ERK, p65, SOS1, Rock1, MMP-2, MM-9, MMP-13 in a dose-dependent manner. Results from real-time PCR analyses revealed that PEITC reduced the mRNA levels of MMP-2, MMP-7, MMP-9 and Rho A in a dose- and time-dependent manner. PEITC had potent anti-cancer activities through the inhibition of migration and invasion in GBM 8401 cells. Our findings elucidates the possible molecular mechanisms and signaling pathways of the anti-metastasis properties of PEITC for human brain glioblastoma cells, and PEITC may be considered to be a therapeutic agent.
Introduction
Glioblastoma is the most aggressive primary brain malignancy with a median survival rate of 14.6 months from diagnosis in unselected patients in molecules, even with maximal, feasible surgical resection, radiotherapy and standard adjuvant temozolomide (TMZ) (1). Only 0.4–0.5% of all GBM patients with extracranial metastasis has been reported, which may be attributable to the extremely shortened survival of these patients (2). Combining radiotherapy and TMZ provides better survival outcomes of glioblastoma than only radiotherapy does (3). Survival and recurrence are significantly associated with extent of resection and residual volume (4). Gross total resection associated with survival improvement is not always possible because the preservation of neurological functions is necessary. The current multimodality treatments of surgery, radiotherapy, chemotherapy for this tumor are still not completely satisfying.
Phenethyl isothiocyanate (PEITC) is one of the most extensively studied isothiocyanates (5). PEITC can induce cell cycle arrest and apoptotic cell death in various tumors (6-12). In our previous study, PEITC induces apoptosis through the extrinsic (death receptor) pathway, dysfunction of mitochondria, ROS induced ER stress, intrinsic (mitochondrial) pathway in GBM 8401 cells (13). PEITC has the anti-metastatic effects in vivo in a novel breast tumor metastasis model (14), and inhibits tumor migration and invasion via suppressing multiple signal transduction pathways in human colon cancer HT29 cells (15). But there is no available literature about how PEITC affects the migration and invasion of human brain glioblastoma cells.
In the present study, we investigated the effects of PEITC on human brain glioblastoma cells in migration and invasion through the signaling transduction pathways in GBM 8401 cells.
Materials and methods
Chemicals and Reagents. PEITC, dimethyl sulfoxide (DMSO), propidium iodide (PI),
RNase, Tris-HCl, Triton X-100 and trypan blue were obtained from Sigma Chemical Co. (St. Louis, MO, USA). RPMI-1640, fetal bovine serum (FBS), L-glutamine, penicillin-streptomycin and trypsin-EDTA were purchased from Gibco BRL/Invitrogen (Carlsbad, CA, USA). Matrigel invasion chambers were obtained from BD Biosciences (San Jose, CA, USA).
Cell culture. GBM 8401 cell line was purchased from the Food Industry Research and
Development Institute (Hsinchu, Taiwan). Cells were plated onto 75 cm2 tissue culture flasks in RPMI 1640 medium supplemented with 10% FBS, 100 Units/ml penicillin and 100 μg/ml streptomycin, 2 mM L-glutamine, and grown at 37°C under a humidified 5% CO2 and 95% air at one atmosphere. The cells were subcultured with a solution of 0.25% trypsin and 0.02% EDTA. The medium was changed every 2 days (16).
Cell morphological changes and viability. GBM 8401 cells (1.6 × 105 cells/well) onto a 12-well plate were treated with 0, 0.5, 1, 2, 4 μM PEITC, or 0, 500 μM TMZ, and incubated for 0, 24 and 48 h. Cells in each well were examined and representative photographs were taken at 200× magnification by Nikon TE2000-U inverted microscope for morphological change examinations. After cells from each well were trypanized and collected by centrifugation at 1500 rpm for 5 min, washed twice with PBS, added with 5 μg/ml PI in PBS to determine the percentage of viable cells. Nonviable cells were stained by PI dye exclusion (indicative of an intact membrane) and displayed brighter fluorescence than the unstained (viable cells). Cells were
counted by flow cytometric analysis with FACSCalibur utilizing Cell Quest software (Becton-Dickinson, San Jose, CA, USA) (17).
Scratch wound healing assay. GBM 8401 cells (1 × 105 cells/well) were placed for 24 h in 6-well plates and a wound at confluence was made with a pipette tip followed by washing with serum-free medium to remove cell debris. They were photographed under phase contrast microscopy (time=0) and then incubated in media with PEITC (0, 2 and 4 μM), or with TMZ (500 μM) at 37˚C and 5% CO2 and allowed to migrate into the wound area for up to 48 h at 37˚C. Cells were gently washed with phosphate buffered saline (PBS). Images of the scratch wounds were quantified by Image J software. The migration inhibited rate = (original scratch width – new scratch width)/original scratch width × 100% (18).
Migration assay. GBM 8401 cells were cultured in serum-free RPMI-1640 medium
containing 1% charcoal-stripped FBS for 48 h. The lower chamber of the Transwell filter was coated with 10 μg type IV collagen, and the lower chamber of the each well was filled with RPMI-1640 supplemented with 1% charcoal-stripped FBS. The filter in 6.5-mm Transwell was inserted in 24-well plates, and about 2.5 × 104 cells/filter of GBM 8401 cells were placed on the filter. These cells were treated with 0, 2, 4 μM PEITC and 500 μM TMZ for 48 h. Migrated cells were stained with 2% crystal violet then were examined and photographed under a microscope (16, 19).
Invasion assay. The same protocols were done as described in the migration assay
except that cells were put on a matrigel-coated Transwell filter (Matrigel invasion chamber, BD Biosciences) then were examined and photographed under a microscope (16, 19).
Gelatin zymography assay. GBM 8401 cells (1 × 106 cells/well) were plated in 12-well tissue culture plates and incubated with 0, 2, 4 μM PEITC or 500 μM TMZ for 24 and 48 h.The conditioned medium was collected and separated by electrophoresis on 10% SDS-PAGE with 0.2% gelatin (Sigma-Aldrich Corp.). The gels were soaked in 2.5% Triton X-100 in dH2O twice for a total of 60 min at 25 ˚C at the end of electrophoresis, and they were incubated in substrate buffer (50 mM Tris HCl, 5 mM CaCl2, 0.02% NaN3 and 1% triton X-100, pH 8.0) at 37 ˚C for 18 h. Bands related to enzyme activities of MMP-2 were visualized by negative staining using 0.2% Coomassie blue in 50% methanol and 10% acetic acid (20). The bands were measured by ImageJ software.
Western blotting assay. GBM 8401 cells (1× 106 cells/dish) were placed in 10 cm dish, and 0, 2, 4 μM PEITC or 500 μM TMZ were added to cells. They were incubated for 0, 24 and 48 h. Cells were collected and lysed in lysate buffer composed of 50 μM Tris (pH 8.0), 150 μM NaCl, 5 μM ethylenediaminetetraacetic acid and 0.5% NP-40 with protease inhibitor solution (Roche, Mannheim, Germany). The protein concentration from each treatment was determined by using Bio-Rad protein assay kit. About 30 μg of protein from each sample was separated on a 10% sodium dodecyl sulfate-polyacrylamide electrophoretic gel (SDS-PAGE) and transferred to nitrocellulose membranes (GE Healthcare, NJ, USA). The blot was soaked with blocking buffer, 5% non-fat dry milk in Tris-buffered saline containing Tween-20 (TBS-T) for 1 h at 25˚C. They were incubated with the specific primary antibodies for matrix metalloproteinases (MMP)-2, MMP-9, Ras, urokinase-type plasminogen
receptor-bound protein 2 (GRB2), p-p38, phospho-Jun NH2-terminal kinase (p-JNK), p-extracellular-signal-regulated kinases (p-ERK), p65, Son of sevenless homolog 1 (SOS1), rho-associated coiled-coil-containing protein kinase 1(Rock1), and MMP-13
(Santa Cruz Biotechnology, Santa Cruz, CA) in blocking buffer at 4˚C for overnight. Immunoreactive proteins were detected with horseradish peroxidase conjugated secondary antibodies and detected by chemiluminescence (GE Healthcare, NJ, USA) and autoradiography by using BioMax LightFilm (Eastman Kodak, New Heaven, CT, USA) (21). The relative proteins amounts from each treatment were assessed by densitometry scanning of X-ray films, and analyzed by Eagle Eye Image System (Stratagene, La Jolla, CA).
Real-time polymerase chain reaction (RT-PCR). GBM 8401 cells (1 × 106 cells/well) onto 6-well tissue culture plates treated with 0, 2, 4 μM PEITC, or 500 μM TMZ, and incubated for 0, 24, 48 h. Cells from each sample was collected, the total RNA was extracted using the Qiagen RNeasy Mini Kit as described previously (16, 22). According to the standard protocol of the supplier (Applied Biosystems), all RNA samples were reverse-transcribed for 30 min at 42˚C with High Capacity cDNA Reverse Transcription Kit. Quantitative PCR conditions were: 2 min at 50˚C, 10 min at 95˚C, and 40 cycles of 15 sec at 95˚C, 1 min at 60˚C using 1 μl of the cDNA reverse-transcribed as described above, 2X SYBR-Green PCR Master Mix (Applied Biosystems) and 200 nM of forward and reverse primers as shown in Table 1. Each assay was processed by using the Applied Biosystems 7300 Real-Time PCR system in triplicate, and fold-changes expression were measured by using the comparative CT method. The ratios of gene expressions related to that for GAPDH were presented.
Statistical analysis. Results were expressed as mean ± SD of 3 experiments. The
differences between PEITC-treated (experimental group) or TMZ-treated (positive control group), and vehicle control group were evaluated by using the Student’s t-test. The p value of less than 0.05 was considered a significant difference. *p<0.05; #p<0.05.
Results
PEITC induces cell morphological changes and didn’t decrease the viability of GBM 8401 cells. GBM 8401 cells were treated with 0, 0.5, 1, 2 and 4 μM PEITC or 500 μM
TMZ for 24, 48 h to realize the cytotoxic effects of PEITC. No marked morphological change of GBM 8401 cells was induced by PEITC (Fig. 1A). Total percentages of viable cells were measured by flow cytometric assay. PEITC or TMZ did not decrease the percentage of viable GBM 8401 cells in a dose- and time-dependent manner (Fig. 1B). The total viable cells were not significantly decreased in GBM 8401 cells exposed as high as 4 μM PEITC or 500 μM TMZ after 24, 48 h-treatment. Consequently, the concentrations of equal or lesser than 4 μM PEITC or 500 μM TMZ were selected in further experiments.
PEITC inhibits the migration of GBM 8401 cells. GBM 8401 cells were incubated
with different concentrations of PEITC and 500 μM TMZ for 48 h to determine the effects of PEITC on cell migration. The scratch wound healing assay was performed, and the results were shown in Figure 2. An apparent and gradual increase of cells in the denuded zone at different concentrations of PEITC was observed with light microscopy. The migration inhibited rates were 12.7%, 42.4%, 44.3%, 44.2% after cells were treated with 0, 2, 4 μM PEITC, 500 μM TMZ for 48 h respectively (Fig. 2B). These effects of PEITC on migration of GBM 8401 cells from scratch wound
healing assay were dose-dependent.
Results from the Transwell migration assay indicated that PEITC significantly inhibited the migration of GBM 8401 cells at concentrations between 2-4 μM (Fig. 3), and the inhibition percentage ranged from 46.89% to 15.75% when cells were incubated with PEITC for 48 h (Fig. 3B). These effects of PEITC on migration of GBM 8401 cells Transwell migration assay were also dose-dependent. TMZ also had an inhibition effect on GBM 8401 cells migration at the concentration of 500 μM.
PEITC inhibits the invasion of GBM 8401 cells. GBM 8401 were able to invade
through a filter coated with Matrigel from the upper to the lower chamber in the control (Fig. 4), but the penetration of the filter by GBM 8401 cells was inhibited by PEITC at the concentrations between 2-4 μM. The inhibition percentage ranged from 27.80% to 7.31% after 48 h-treatment (Fig. 4B). These effects of PEITC on invasion were in a dose-dependent manner. The invasion of GBM 8401 cells was also inhibited by 500 μM TMZ.
PEITC decreased the enzyme activities of MMP-2 in GBM 8401 cells. Gelatin
zymography assay indicated the enzyme activities of MMP-2 were reduced in dose-dependent manner after GBM 8401 cells were treated with 2, 4 μM PEITC for 24, 48 h (Fig. 5). The enzyme activities of MMP-2 were also decreased after cells were treated with 500 μM TMZ for 24, 48 h (Fig. 5).
PEITC inhibited the levels of proteins associated with migration and invasion in GBM 8401 cells. Western blotting assay was applied to determine the effects of
PEITC and TMZ on the levels of proteins associated with migration and invasion of GBM 8401 cells. PEITC decreased the protein levels of Ras, uPA, Rho A, GRB2
(Fig. 6A), p-p38, p-JNK, p-ERK, p65 (Fig. 6B), SOS1, Rock1, MMP-2, MM-9, MMP-13 (Fig. 6C) in a dose-dependent manner after cells were treated with 2, 4 μM PEITC for 48 h. TMZ reduced the protein levels of Ras, uPA, Rho A, GRB2, p-p38, p-JNK, p-ERK, p65, SOS1, Rock1, MMP-2, MMP-9, MMP-13 (Fig. 6A-C).
PEITC inhibited the levels of mRNA expressions in GBM 8401 cells. To investigate
the effects of PEITC on migration- and invasion-associated gene expressions in GBM 8401 cells, cells were treated with 2, 4 μM PEITC for 24 and 48 h. Real-time PCR analyses were applied to measure the mRNA levels of these gene expressions. PEITC inhibited the mRNA levels of MMP-2 (Fig. 7A), MMP-7 (Fig. 7B), MMP-9 (Fig. 7C) and Rho A (Fig. 7D) in a dose- and time-dependent manner.
Discussion
There are some studies about the effects of PEITC on human glioma cells (23, 24). PEITC also induces apoptosis of human brain glioblastoma cells from our previous study (13). The cell morphology was not significantly changed (Fig. 1A), and the cell viability were not significantly decreased in GBM 8401 cells exposed as high as 4 μM PEITC, or 500 μM TMZ after 24, 48 h-treatment (Fig. 1B). So the concentrations of PEITC and TMZ for cell migration and invasion studies were determined. From the Transwell migration assay, PEITC significantly inhibited the migration of GBM 8401 cells at concentrations between 2-4 μM in a dose-dependent manner (Fig. 3), and the inhibition percentage ranged from 46.89% to 15.75% when cells were incubated with PEITC for 48 h (Fig. 3B). From invasion assay, PEITC also significantly inhibited the invasion of GBM 8401 cells at concentrations between 2-4 μM in a dose-dependent manner (Fig. 4), and the inhibition percentage ranged from 27.80% to 7.31% after 48 h-treatment (Fig. 4B). The current standard chemotherapy, TMZ also had inhibitory
effects on migration and invasion of GBM 8401 cells at the concentration of 500 μM. MMPs, a family of zinc-dependent endopeptidases, play the roles in brain development, synaptic plasticity and repair after injury to the pathogenesis of various brain disorders (25). MMP-mediated extracellular matrix (ECM) degradation promotes tumor invasion, progression and involves the angiogenesis and metastasis. MMPs are able to degrade almost all known ECM components and play important roles in mediating glioblastoma tumor cell invasion (26). The levels of MMP-2, MMP-9, and membrane type 1 (MT1)-MMP expression in gliomas are higher than those in normal brain tissue. MMP-13 enzymatic activity is critical to highly invasive potential of cancer stem cells of human glioblastoma cell line U251 (27). The levels of MMP-7 expression are correlated with tumor aggressiveness and poor prognosis in solid tumors, but they are highly variable in patients with glioblastoma (27). The cross-talk between the tumor and the surrounding stroma to regulate MMP-7 exists: the expression of MMP-7 in human U87 glioma cells is low in culture, higher when the cells are implanted within the brain. In this study, PEITC reduced the enzyme activities of MMP-2 in a dose- and time-dependent manner after GBM 8401 cells were treated with 2, 4 μM PEITC for 24 h and 48 h (Fig. 5). It also decreased the protein levels of MMP-2, MM-9, MMP-13 (Fig. 6C) in a dose-dependent manner after cells were treated with 2, 4 μM PEITC for 48 h. PEITC inhibited the mRNA levels of MMP-2 (Fig. 7A), MMP-7 (Fig. 7B), MMP-9 (Fig. 7C) in a dose- and time-dependent manner. Taken together, PEITC may inhibit the migration and invasion of GBM 8401 cells through reduction in the enzyme activities of MMP-2, the protein levels of MMP-2, MMP-9, MMP-13, and the mRNA levels of MMP-2, MMP-7 and MMP-9.
may be enhanced by uPA-mediated direct activation of MMP-9 (28). The enhanced invasive capacity of peritumoral cells in GBM needs simultaneous Rac and Rho A activation (29). Knockdown of GRB2, mediating receptor tyrosine kinase-induced activation of RAS and downstream signaling, can reduce invasive activity of breast cancer (30). Epidermal growth factor receptor (EGFR) vIII-mediated migration and transformation in U87MG (PTEN mutant) glioblastoma cells is down-regulated by the effects of signal regulatory protein alpha1 (SIRPalpha1) on the activation loop of SHP-2/FAK/GRB2/SOS-1/MAPK (31). Knockdown of Rho A inhibits the expression of p-JNK and phospho-c-Jun (p-c-Jun), reduces MMP2 activity and cell invasion in human glioma U251 cells in hypoxic conditions (32). ROCK-dependent signaling pathways involves the glioma migration, and antidromic effects on glioma migration are made by selective knockdown of either ROCK1 or ROCK2 (33). ROCK1 knockdown inhibits cell proliferation, but ROCK2 knockdown promotes it. PEITC inhibited the protein levels of Ras, uPA, Rho A, GRB2 (Fig. 6A), p38, JNK, p-ERK, p65 (Fig. 6B), SOS1, Rock1 (Fig. 6C) in a dose-dependent manner after cells were treated with 2, 4 μM PEITC for 48 h. PEITC decreased the mRNA levels of Rho A (Fig. 7D) in a dose- and time-dependent manner. Taken together, PEITC may inhibit the migration and invasion of GBM 8401 cells through reduction in the protein levels of Ras, uPA, Rho A, GRB2, p-p38, p-JNK, p-ERK, p65, SOS1, Rock1, and the mRNA levels of Rho A.
In conclusion, our experiments indicated PEITC had potent anti-cancer activities through the inhibition of migration and invasion in GBM 8401 cells. PEITC decreased the expressions of MMP-2, MMP-7, MMP-9, MMP-13, Ras, uPA, Rho A, GRB2, p-p38, p-JNK, p-ERK, p65, SOS1, Rock1 in GBM 8401 cells in vitro (Fig. 8). PEITC may have the therapeutic potentials, and our findings disclose the possible
molecular mechanisms and signaling pathways of the anti-cancer properties of PEITC for human brain glioblastoma cells.
Acknowledgments This work was supported by grant TCVGH-1044903B from Taichung Veterans General Hospital, Taichung, Taiwan
Conflict of interest The authors declare no conflict of interest.
References
1. Stupp R, Mason WP, van den Bent MJ, et al.: Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med 352: 987-996, 2005.
2. Lun M, Lok E, Gautam S, Wu E and Wong ET: The natural history of extracranial metastasis from glioblastoma multiforme. J Neurooncol 105: 261-273, 2011.
3. Yang LJ, Zhou CF and Lin ZX: Temozolomide and radiotherapy for newly diagnosed glioblastoma multiforme: a systematic review. Cancer Invest 32: 31-36, 2014.
4. Chaichana KL, Jusue-Torres I, Navarro-Ramirez R, et al.: Establishing percent resection and residual volume thresholds affecting survival and recurrence for patients with newly diagnosed intracranial glioblastoma. Neuro Oncol 16: 113-122, 2014.
5. Moon YJ, Brazeau DA and Morris ME: Dietary phenethyl isothiocyanate alters gene expression in human breast cancer cells. Evid Based Complement Alternat Med 2011, 2011.
6. Antosiewicz J, Ziolkowski W, Kar S, Powolny AA and Singh SV: Role of reactive oxygen intermediates in cellular responses to dietary cancer chemopreventive agents. Planta Med 74: 1570-1579, 2008.
7. Chen YR, Han J, Kori R, Kong AN and Tan TH: Phenylethyl isothiocyanate induces apoptotic signaling via suppressing phosphatase activity against c-Jun N-terminal kinase. J. Biol. Chem. 277: 39334-39342, 2002.
8. Hu R, Kim BR, Chen C, Hebbar V and Kong AN: The roles of JNK and apoptotic signaling pathways in PEITC-mediated responses in human HT-29 colon adenocarcinoma cells. Carcinogenesis 24: 1361-1367, 2003.
9. Jakubikova J, Bao Y and Sedlak J: Isothiocyanates induce cell cycle arrest, apoptosis and mitochondrial potential depolarization in HL-60 and multidrug-resistant cell lines. Anticancer Res 25: 3375-3386, 2005.
10. Kang L and Wang ZY: Breast cancer cell growth inhibition by phenethyl isothiocyanate is associated with down-regulation of oestrogen receptor-alpha36. J Cell Mol Med 14: 1485-1493, 2010.
11. Telang U, Brazeau DA and Morris ME: Comparison of the effects of phenethyl isothiocyanate and sulforaphane on gene expression in breast cancer and normal mammary epithelial cells. Exp Biol Med (Maywood) 234: 287-295, 2009.
12. Tseng E, Scott-Ramsay EA and Morris ME: Dietary organic isothiocyanates are cytotoxic in human breast cancer 7 and mammary epithelial MCF-12A cell lines. Exp Biol Med (Maywood) 229: 835-842, 2004.
13. Chou YC, Chang MY, Wang MJ, et al.: PEITC induces apoptosis of Human Brain Glioblastoma GBM8401 Cells through the extrinsic- and intrinsic -signaling pathways. Neurochem. Int. 81: 32-40, 2015.
14. Gupta P, Adkins C, Lockman P and Srivastava SK: Metastasis of Breast Tumor Cells to Brain Is Suppressed by Phenethyl Isothiocyanate in a Novel Metastasis Model. PLoS One 8: e67278, 2013.
15. Lai KC, Hsu SC, Kuo CL, et al.: Phenethyl isothiocyanate inhibited tumor migration and invasion via suppressing multiple signal transduction pathways in human colon cancer HT29 cells. J Agric Food Chem 58: 11148-11155, 2010.
16. Lin CC, Chen JT, Yang JS, et al.: Danthron inhibits the migration and invasion of human brain glioblastoma multiforme cells through the inhibition of mRNA expression of focal adhesion kinase, Rho kinases-1 and metalloproteinase-9. Oncol Rep 22: 1033-1037, 2009.
17. Lu HF, Lai TY, Hsia TC, et al.: Danthron induces DNA damage and inhibits DNA repair gene expressions in GBM 8401 human brain glioblastoma multiforms cells. Neurochem. Res. 35: 1105-1110, 2010.
18. Shang HS, Chang JB, Lin JH, et al.: Deguelin inhibits the migration and invasion of U-2 OS human osteosarcoma cells via the inhibition of matrix metalloproteinase-2/-9 in vitro. Molecules 19: 16588-16608, 2014.
19. Lu KW, Chen JC, Lai TY, et al.: Gypenosides inhibits migration and invasion of human oral cancer SAS cells through the inhibition of matrix metalloproteinase-2 -9 and urokinase-plasminogen by ERK1/2 and NF-kappa B signaling pathways. Hum Exp Toxicol 30: 406-415, 2011.
invasion in human osteosarcoma U-2 OS cells through suppressing the matrix metalloproteinase-2/-9, protein kinase B (PKB) and PKC signaling pathways. Food Chem Toxicol 50: 1734-1740, 2012.
21. Ho CC, Huang AC, Yu CS, et al.: Ellagic acid induces apoptosis in TSGH8301 human bladder cancer cells through the endoplasmic reticulum stress- and mitochondria-dependent signaling pathways. Environ. Toxicol. 29: 1262-1274, 2014.
22. Lin HJ, Su CC, Lu HF, et al.: Curcumin blocks migration and invasion of mouse-rat hybrid retina ganglion cells (N18) through the inhibition of MMP-2, -9, FAK, Rho A and Rock-1 gene expression. Oncol Rep 23: 665-670, 2010. 23. Gupta B, Chiang L, Chae K and Lee DH: Phenethyl isothiocyanate inhibits
hypoxia-induced accumulation of HIF-1alpha and VEGF expression in human glioma cells. Food Chem 141: 1841-1846, 2013.
24. Lee DH, Kim DW, Lee HC, Lee JH and Lee TH: Phenethyl isothiocyanate sensitizes glioma cells to TRAIL-induced apoptosis. Biochem. Biophys. Res. Commun. 446: 815-821, 2014.
25. Kim YS and Joh TH: Matrix metalloproteinases, new insights into the understanding of neurodegenerative disorders. Biomol Ther (Seoul) 20: 133-143, 2012.
26. Chintala SK, Tonn JC and Rao JS: Matrix metalloproteinases and their biological function in human gliomas. Int J Dev Neurosci 17: 495-502, 1999. 27. Inoue A, Takahashi H, Harada H, et al.: Cancer stem-like cells of glioblastoma
characteristically express MMP-13 and display highly invasive activity. Int J Oncol 37: 1121-1131, 2010.
28. Zhao Y, Lyons CE, Jr., Xiao A, et al.: Urokinase directly activates matrix metalloproteinases-9: a potential role in glioblastoma invasion. Biochem. Biophys. Res. Commun. 369: 1215-1220, 2008.
29. Ruiz-Ontanon P, Orgaz JL, Aldaz B, et al.: Cellular plasticity confers migratory and invasive advantages to a population of glioblastoma-initiating cells that infiltrate peritumoral tissue. Stem Cells 31: 1075-1085, 2013.
30. Xu Y, Zhang H, Lit LC, et al.: The kinase LMTK3 promotes invasion in breast cancer through GRB2-mediated induction of integrin beta(1). Sci Signal 7: ra58, 2014.
31. Kapoor GS and O'Rourke DM: SIRPalpha1 receptors interfere with the EGFRvIII signalosome to inhibit glioblastoma cell transformation and migration. Oncogene 29: 4130-4144, 2010.
32. Tong JJ, Yan Z, Jian R, Tao H, Hui OT and Jian C: RhoA regulates invasion of glioma cells via the c-Jun NH2-terminal kinase pathway under hypoxia.
Oncol Lett 4: 495-500, 2012.
33. Mertsch S and Thanos S: Opposing signaling of ROCK1 and ROCK2 determines the switching of substrate specificity and the mode of migration of glioblastoma cells. Mol. Neurobiol. 49: 900-915, 2014.
Figures legends
Figure 1. PEITC and TMZ did not induce marked cell morphological changes and did not decrease the percentage of viable GBM 8401 cells. Cells were treated with different concentrations (0.5-4 μM) of PEITC for 24, 48 h and cells morphological changes were examined under phase contrast microscope at ×200 (A). Cells were harvested to calculate the percentage of viable cells by flow cytometric assay (B). The values presented are the mean ± S. D. (n = 3) from three independent experiments. There was no significant difference between PEITC-treated (experimental group) or TMZ-treated (positive control group) and vehicle control group. C: control, TMZ: temozolomide.
Figure 2. Effects of PEITC on the migration of GBM 8401 cells in scratch wound healing migration assay. GBM 8401 cells were placed on the plate and a wound line was made with a pipette tip. The cells were incubated with 0, 2, 4 μM PEITC or 500 μM TMZ for 0 and 48 h. Cell migration was assessed by microscopy at the indicated time points at ×100 (A). Migration inhibited rates were counted at ×100 (B). *P<0.05, significant difference between PEITC-treated or TMZ-treated groups and the control.
Figure 3. PEITC inhibited the migration of GBM 8401 cells. GBM 8401 cells (2.5 × 104 cells/filter) were treated with 0, 2, 4 μM PEITC or 500 μM TMZ for 0 and 48 h. Cells on the Transwell filter migrated to the lower surface of the filter were stained with crystal violet, and were photographed under a light microscope at ×200 (A).
Cells from the lower chamber were counted at ×200 (B). *P<0.05, significant difference between PEITC-treated or TMZ-treated groups and the control.
Figure 4. PEITC inhibited the migration of GBM 8401 cells. GBM 8401 cells (2.5 × 104 cells/filter) were treated with 0, 2, 4 μM PEITC or 500 μM TMZ for 48 h. Cells on the Transwell filter penetrated through the matrigel to the lower surface of the filter were stained with crystal violet, and were photographed under a light microscope at ×200 (A). Cells from the lower chamber were counted at ×200 (B). *P<0.05, significant difference between PEITC-treated or TMZ-treated groups and the control.
Figure 5. Effects of PEITC on the enzyme activities of MMP-2 in GBM 8401 cells. Cells were treated with 0, 2, 4 μM PEITC or 500 μM TMZ for 24, 48 h. Cells were harvested for examining MMP-2 activities from each treatment that were determined by Gelatin zymography assay as described in Materials and Methods.
Figure 6. Effects of PEITC on the levels of proteins associated with migration and invasion in GBM 8401 cells. Cells were treated with 0, 2, 4 μM PEITC or 500 μM TMZ for 48 h. The proteins levels from each sample were determined by SDS-PAGE and Western blotting. A: Ras, uPA, Rho A, GRB2; B: p-p38, p-JNK, p-ERK, p65 and C: SOS1, Rock1, MMP-2, MMP-9, MMP-13.
Figure 7. PEITC inhibits the levels of mRNA expression in GBM 8401 cells. Cells were treated with 0, 2, 4 μM PEITC or 500 μM TMZ for 24 and 48 h. RNA samples were reverse-transcribed to obtain cDNA for real-time PCR. The ratios of gene expressions related to that for GAPDH are presented in A: MMP-2, B: MMP-7, C:
MMP-9 and D: Rho A. Data represents the mean ± S.D. of three experiments. Significantly different between PEITC treatment and control groups (*P<0.05, #P<0.05).
Figure 8. The possible signaling pathways of PEITC-inhibited migration and invasion in human glioblastoma multiforme GBM 8401 cells.
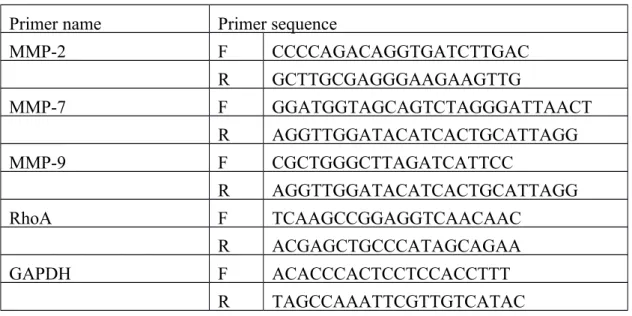
Table 1. Primers sequence used for real-time PCR
Primer name Primer sequence
MMP-2 F CCCCAGACAGGTGATCTTGAC R GCTTGCGAGGGAAGAAGTTG MMP-7 F GGATGGTAGCAGTCTAGGGATTAACT R AGGTTGGATACATCACTGCATTAGG MMP-9 F CGCTGGGCTTAGATCATTCC R AGGTTGGATACATCACTGCATTAGG RhoA F TCAAGCCGGAGGTCAACAAC R ACGAGCTGCCCATAGCAGAA GAPDH F ACACCCACTCCTCCACCTTT R TAGCCAAATTCGTTGTCATAC
MMP, Matrix metalloproteinase; RhoA, RAS homologue gene family member A; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; F, forward primers; R, reverse primers.