行政院國家科學委員會專題研究計畫 成果報告
T 淋巴細胞免疫反應調控基因的辨認與研究(3/3)
研究成果報告(完整版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2320-B-002-003- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學醫學院免疫學研究所 計 畫 主 持 人 : 繆希椿 計畫參與人員: 博士班研究生-兼任助理:林伯修、曹孝瑋 碩士班研究生-兼任助理:顏偉峰、蔡佩芸、謝婉云、林俊國 處 理 方 式 : 本計畫可公開查詢中 華 民 國 96 年 10 月 31 日
T 淋巴細胞免疫反應調控基因的辨認與研究(3/3) 關鍵詞: T 淋巴細胞,基因調控及免疫反應
中文摘要
T 淋巴細胞特異的轉錄因子:T-bet、GATA3 與 c-MAF 在 Th1/Th2 的發育上是 必需的。T-bet 是 Th1 細胞分化的主要調控者,然而 GATA3 與 c-MAF 則在 Th2 的 發育上扮演決定性的角色。其它因子,如細胞激素、多肽媒介者 Eta-1、化學激 素 MCP-1、訊息傳導及活化者 4(STAT4)及 STAT6 也會影響 T 細胞的分化。然而許 多實驗指出,一些其他未被確定的因子也參與 T 細胞的分化。所以,我們計畫有 效率的篩選和辨識這些參與 T 細胞分化的因子,並在體內及體外模式中研究他們 在 Th1/Th2 的發育與免疫反應中的角色。目前我們已成功地從酵母雙雜交系統篩 選出與 c-MAF 交互作用的分子。我們的研究著重在探討分子與細胞機轉上,這些 c-MAF 交互作用的分子如何影響 c-MAF 的功能,進而調節 T 細胞分化與免疫反應。 前趨致癌基因 Ets-1 在發炎及抗發炎免疫反應中扮演特定性的角色。Ets-1 已經被證明在調控 T 淋巴細胞中扮演重要的角色,其中包括 T 細的發育、分化以 及細胞凋亡。而近來也發現 Ets-1 也參與了調節輔助型 T 細胞所分泌的細胞激 素,像是介白素 2、介白素 4、介白素 10 及γ干擾素。雖然有關 Ets-1 的分子調 控已經被廣泛的研究,但 sumoylation 在 Ets-1 的調節機制中所擔任的功能卻仍 然不是很清楚。我們證明了破壞 Ets-1 上的 SUMO 結合區,並不會影響 Ets-1 和 T-bet 對γ干擾素的轉錄作用。利用 Ets-1 最小啟動子所進行的螢光素酶分析實 驗,我們更進一步證明了即使將 Ets-1 上的 sumoylation 位置利用定點突變破壞 掉,Ets-1 本身的轉錄能力也不會有所改變。而即使在加入了 SUMO 連接酶中的 PIAS 蛋白後,野生型和 SUMO 區突變型的 Ets-1,其轉錄能力亦仍然相同。也許 sumoylation 並不影響 Ets-1 的轉錄能力,但既然 Ets-1 已經被證明的確可以被 SUMO 所結合修飾,此 SUMO 化的 Ets-1 在免疫上的功能更需要我們去進一步探索 的。
Identification and Study of Genes that Regulate T Lymphocyte Differentiation and Immune Response (3/3)
Keywords: T Lymphocyte, Gene Regulation and Immune Response
T lymphocyte specific transcription factors T-bet, GATA3 and c-MAF have essential roles in the Th1/Th2 development. T-bet is the master regulator for the development of the Th1 cell lineage, while GATA3 and c-MAF are crucial for the development of the Th2 cell lineage. Other factors, such as cytokines, polypeptide mediator Eta1, chemokine MCP-1, signal transducer and activator of transcription 4 (STAT4), and STAT6 also influence the T lymphocyte differentiation. However, many studies indicate that some other factors involved in Th differentiation are yet to be identified. Therefore, we intend to effectively screen and identify the factors crucial for T lymphocyte differentiation and study their roles in Th1/Th2 and immune response in vitro and in vivo. We have screened successfully the c-MAF-interacting proteins by yeast-two-hybrid system. Currently, we focus to study their roles on the function of c-MAF. Thus, we hope to provide a better insight of the molecular and cellular mechanisms by which genes regulate T lymphocyte differentiation and immune response.
Ets-1 has been demonstrated to play some roles in the regulation of T lymphocytes, which including T cell development, differentiation and apoptosis. Recently, Ets-1 has been reported to be a modulator of T helper cell cytokine secreting profiles which include interleukin-2, interleukin-4, interleukin-10 and interferon-γ. Although the regulation of Ets-1 has been extensively studied, the role of sumoylation on the function of Ets-1 remains elusive. Here, we demonstrated that abolishing the consensus SUMO conjugation motif in Ets-1 did not alter the transcription of IFNγ by Ets-1 and T-bet. Furthermore, we demonstrated that the transactivation ability of Ets-1 on the reporter carrying Ets-1-binding region had been no changed when we mutated the sumoylation sites. Moreover, wild-type and sumoylated Ets-1 have the similar transactivation ability in the presence of PIAS proteins, SUMO E3 ligase. These results imply that sumoylation may play no role on Ets-1’s transcriptional activities. Therefore, the functions of sumoylated Ets-1 need to be further investigated.
Introduction
The cytokines have major roles in Th differentiation. Recent studies demonstrate the cytokines, mainly IFNγ and IL4, influence the differentiation of Th cells by controlling subsets of specific transcription factors, T-bet and GATA3, in a directional and positive feedback manner. Receptors for Th1 or Th2 cytokines also involve in the decision of Th1 or Th2 lineage commitment. For example, the IL12 receptor β chain is the one of the key factors for the Th1-lineage pathway. Other factors, such as cytokines, polypeptide mediator Eta1, chemokine MCP-1, signal transducer and activator of transcription 4 (STAT4), and STAT6 also influence the T lymphocyte differentiation.
Further studies have shown that T lymphocyte specific transcription factors T-bet, GATA3 and c-MAF have essential roles in the Th1/Th2 development. T-bet is the master regulator for the development of the Th1 cell lineage, while GATA3 and c-MAF are crucial for the development of the Th2 cell lineage. ROG, a new member of the POZ family of transcriptional repressors, was initially cloned as a GATA-3 interacting protein. In vitro, ROG can serve as a GATA-3-dependent transcriptional repressor. ROG transcripts can be detected in both Th1 and Th2 cells. Furthermore, over-expression of ROG in Th clones inhibits cytokine production.
However, many studies indicate that other factors involved in Th differentiation are yet to be identified. Therefore, in this proposal, we intend to effectively screen and identify the factors crucial for T lymphocyte differentiation by using the yeast
two-hybrid system and study their roles in Th1/Th2 and immune response in vitro and in vivo.
Grenningloh et al. have identified that Ets-1 as a functional cofactor of T-bet and is essential for Th1 responses. In the absence of Ets-1, IFN-γ production by Th1-cell is decrease and the production of some of the Th2 cytokines, such as IL-10, are increase. They also demonstrated that the Ets-1 is recruited to the IFN-γ promoter and enhances the function of T-bet. But in their experiments, they have been unable to demonstrate the existence of an Ets-1/T-bet complex in Th cells by
coimmunoprecipitation. This implies that Ets-1 may not directly interact with T-bet, but through another interacting protein to modulate the function of T-bet. Previous studies about Ets-1 have focus on its interacting proteins and phosphorylation of Ets-1. Phosphorylation of Ets-1 has been demonstrated that can alter the DNA binding
abilities of Ets-1. Hahn et al. and Macauley et al. have demonstrated that Ets-1 can interact with the SUMO E2 conjugating enzyme, Ubc9 and further sumoylated within its N-terminal. These researches broaden our point of view that the sumoylation of Ets-1 may play some roles in the regulation of IFNγ within Th1 cells. Thus, there are
three aims in this study. First, we would like to ask whether the Ets-1 is sumoylated in T helper cells. Second, we need to define the specific E3 ligase for Ets-1 sumoylation. Finally, the roles of sumoylated Ets-1 in T helper cells need to be discovered.
Specific aim 1: To identify the interacting proteins of T lymphocyte specific transcription factors
T lymphocyte specific transcription factors T-bet, GATA3 and c-MAF have major roles in the Th1/Th2 development. T-bet is the master regulator for the development of the Th1 cell lineage, while GATA3 and c-MAF are crucial for the development of the Th2 cell lineage. ROG is a transcriptional repressor of GATA3. Over-expression of ROG inhibits both Th1 and Th2 cytokines production in Th1 and Th2 clones. How do these factors regulate the differentiation of T cells and immune response? Are any other factors involved to mediate their function? To get a better picture, we plan to conduct a comprehensive screening to identify the interacting protein of these T lymphocyte specific transcription factors by using the yeast two-hybrid system.
Experimental design: To identify the interacting proteins of T lymphocyte specific transcription factor c-Maf
The studies described in this aim focus on the isolation of the interacting proteins of c-Maf. c-Maf is crucial for the development of the Th2 cell lineage. I will attempt to clone comprehensively possible interacting partner(s) with the yeast two-hybrid system by using different domains of c-Maf as baits and cDNA libraries from mouse CD4 and CD8 clones as preys.
Results:
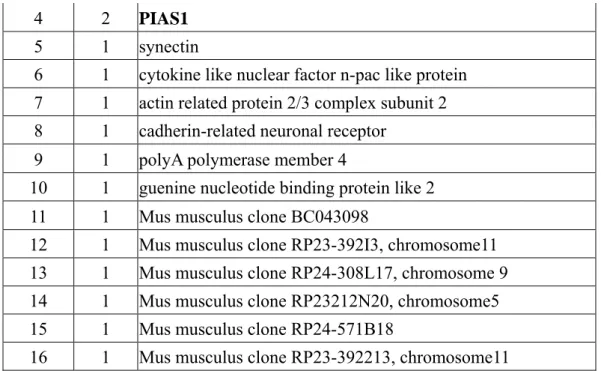
(A) We have obtained 90 positive colonies from two screenings. Reconfirmation and identification of these colonies have been performed (Tables 1 and 2).
Table 1. The candidates of c-Maf-interacting proteins obtaining by yeast-two-hybrid using c-maf (138bp) as a bait and CD4+ AE7 library as a prey
Group Repeat Gene
1 30 UBC9
2 3 snRNP70
4 2 PIAS1
5 1 synectin
6 1 cytokine like nuclear factor n-pac like protein 7 1 actin related protein 2/3 complex subunit 2 8 1 cadherin-related neuronal receptor
9 1 polyA polymerase member 4
10 1 guenine nucleotide binding protein like 2 11 1 Mus musculus clone BC043098
12 1 Mus musculus clone RP23-392I3, chromosome11 13 1 Mus musculus clone RP24-308L17, chromosome 9 14 1 Mus musculus clone RP23212N20, chromosome5 15 1 Mus musculus clone RP24-571B18
16 1 Mus musculus clone RP23-392213, chromosome11
Table 2. The candidates of c-Maf-interacting proteins obtaining by yeast-two-hybrid using c-Maf (hinge domain) as a bait and CD8+ L3 library as a prey
Group Repeat Gene
1 5 Mus musculus RIKEN cDNA 2010311D03 gene
(2010311D03Rik), mRNA
2 5 Mus musculus integrin beta 7, mRNA
3 3 Mus musculus adrenodoxin mRNA, complete cds;Mus musculus ferredoxin 1 (Fdx1), mRNA
4 3 Mouse protein tyrosine phosphatase (70zpep) mRNA, complete cds 5 3 Mus musculus endothelial cell-specific molecule 1 (Esm1), mRNA 6 2 Mus musculus golgi phosphoprotein 3-like, mRNA 7 3 Mus musculus heat shock protein 8 (Hspa8), mRNA
8 1 Mus musculus cleavage and polyadenylation specific factor 2 (Cpsf2)
9 1 Mus musculus S-adenosylhomocysteine hydrolase (Ahcy), mRNA 10 1 Mus musculus replication protein A1(Rpa1), mRNA
11 1 Mus musculus topoisomerase (DNA) II beta binding protein, mRNA 12 1 Mus musculus protein kinase, cAMP dependent regulatory, type I, alpha 13 1 Mus musculus polymerase (RNA) II (DNA directed) polypeptide J 14 1 Mus musculus similar to Speer1-ps1 protein
15 1 Mus musculus proteasome (prosome, macropain) subunit, beta type 7 16 1 Mus musculus spectrin alpha 2, mRNA
17 1 Mus musculus 0 day neonate head cDNA, RIKEN full-length enriched library, clone:4833426G12
18 1 Mus musculus kidney CCL-142 RAG cDNA
19 1 Mus musculus CH-rich interacting match of PLAG1 mRNA, complete cds Mus musculus ring finger and CHY zinc finger domain protein containing 1 20 1 Mus musculus ring finger and CHY zinc finger domain containing 1 21 1 Mus musculus BAC clone RP23-257F7 from 7, complete sequence 22 1 Mus musculus NADH dehydrogenase 6, mitochondrial,
mRNA
23 1 Mus musculus RIKEN cDNA 2010012C16 gene (2010012C16Rik), mRNA (B) c-Maf and PIAS1 interact directly in vivo
293T cells were transfected with myc-tagged PIAS1 and HA-tagged c-Maf or both. Cells were lysed after 24hr, and cell extracts were immunoprecipitation by anti-HA (A) or anti-Myc (B) antibodies and then immunobloting by anti-Myc (A) or anti-HA (B) antibodies respectively.
A
B
IP-c-Maf (HA), IB-PIAS1 (Myc):
HA-c-
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-f
(HA)
HA-c-Maf
+ +
Myc-PIAS1 + −
c-Maf 50 kD Ig band
PIAS1
IP-c-
(HA), IB-PIAS1 (Myc):
HA-c-Maf
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-f
(HA)
HA-c-
+ +
Myc-PIAS1 + −
c-IP-c-
(HA), IB-PIAS1 (Myc):
HA-c-
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-Maf (HA)
HA-c
+ +
Myc-PIAS1 + −
c- 50 kD Ig band PIAS1A
B
IP-c-Maf (HA), IB-PIAS1 (Myc):
HA-c-
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-f
(HA)
HA-c-Maf
+ +
Myc-PIAS1 + −
c-Maf 50 kD Ig band
PIAS1
IP-c-
(HA), IB-PIAS1 (Myc):
HA-c-Maf
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-f
(HA)
HA-c-
+ +
Myc-PIAS1 + −
c-IP-c-
(HA), IB-PIAS1 (Myc):
HA-c-
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-Maf (HA)
HA-c
+ +
Myc-PIAS1 + −
c- 50 kD Ig band
PIAS1
IP-c-Maf (HA), IB-PIAS1 (Myc):
HA-c-
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-f
(HA)
HA-c-Maf
+ +
Myc-PIAS1 + −
c-Maf
IP-c-Maf (HA), IB-PIAS1 (Myc):
HA-c-
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-f
(HA)
HA-c-Maf
+ +
Myc-PIAS1 + −
c-Maf 50 kD Ig band
PIAS1
IP-c-
(HA), IB-PIAS1 (Myc):
HA-c-Maf
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-f
(HA)
HA-c-
+ +
Myc-PIAS1 + −
c-IP-c-
(HA), IB-PIAS1 (Myc):
HA-c-
+ −
Myc-PIAS1 + +
IP-PIAS1 (Myc), IB-c-Maf (HA)
HA-c
+ +
Myc-PIAS1 + −
c- 50 kD Ig band
PIAS1
(C) Effect of c-Maf-induced transactivation by PIAS1
2×105 293T cells were transfected with 50ng of IL-4 promoter-luciferase reporter together with 10, 50 or 100ng of c-maf respectively, and increasing amounts of PIAS1 as indicated. Cells were lysed after 24hr, and cell extracts were assayed by luciferase activity. Fold activation was calculated as the increase in luciferase activity by c-maf and PIAS1, and compared with basal reporter gene activity. All luciferase data were correlated by co-transfected Renilla level.
(D) Effect of PIAS1 on c-maf transactivation ability
2×105 293T cells were transfected with 50ng of IL-4 promoter-luciferase reporter and 25ng of wild type, K29R, K33R or K328R, K29,33R or K29,33,328R c-Maf
c-maf, together with 0, 10, 25 or 50ng PIAS1 respectively. Cells were lysed after 24hr, and cell extracts were assayed by luciferase activity. Fold activation was calculated as the increase in luciferase activity by c-Maf and compared with basal reporter gene activity. All luciferase data were correlated by co-transfected Renilla level.
0 50 100 150 200 0ng 5ng 10ng 50ng 100ng
PIAS1
re
la
tiv
e
fo
ld
c-maf 10ng c-maf 50ng c-maf 100ng 0 20 40 60 80 100 120 140 0ng 10ng 25ng 50ng PIAS1 fo ld in d uc tio n Wt K29R K33R K328R(E) Effect of UBC9 on c-maf transactivation ability
2×105 293T cells were transfected with 50ng of IL-4 promoter-luciferase reporter and 25ng c-maf, together with 0, 10, 25 or 50ng UBC9 respectively. Cells were lysed after 24hr, and cell extracts were assayed by luciferase activity. Fold activation was
calculated as the increase in luciferase activity by c-maf and compared with basal reporter gene activity. All luciferase data were correlated by co-transfected Renilla level. 0 20 40 60 80 100 120 140 0ng 10ng 25ng 50ng PIAS1 fo ld i nd uc tio n WT K29,33R K29,33,328R 0 20 40 60 80 100 120 140 0ng 10ng 25ng 50ng UBC9 fo ld i nd uc tio n UBC9
Conclusion:
We have effectively screened and identified the factors which interacting with Th2 specific transcription factor c-Maf. Currently, we are studying whether the c-Maf is post-modified in vivo and its role on the regulation of IL-4, a Th2 specific cytokine.
Reference:
Ashkar, S., Weber, G. F., Panoutsakopoulou, V., Sanchirico, M., Jansson, M.,
Zawaideh, S., Rittling, S. R., Denhardt, D. T., Glimcher, M. J., and Cantor, H. (2000). Eta-1 (osteopontin): an early component of type-1 (cell-mediated) immunity. Science 287, 860-864.
Dai, M. S., Chevallier, N., Stone, S., Heinrich, M. C., McConnell, M., Reuter, T., Broxmeyer, H. E., Licht, J. D., Lu, L., and Hoatlin, M. E. (2002). The effects of the Fanconi anemia zinc finger (FAZF) on cell cycle, apoptosis, and proliferation are differentiation stage-specific. J Biol Chem 277, 26327-26334.
Finotto, S., De Sanctis, G. T., Lehr, H. A., Herz, U., Buerke, M., Schipp, M., Bartsch, B., Atreya, R., Schmitt, E., Galle, P. R., et al. (2001). Treatment of allergic airway inflammation and hyperresponsiveness by antisense-induced local blockade of GATA-3 expression. J Exp Med 193, 1247-1260.
Glimcher, L. H., and Murphy, K. M. (2000). Lineage commitment in the immune system: the T helper lymphocyte grows up. Genes Dev 14, 1693-1711.
Gu, L., Tseng, S., Horner, R. M., Tam, C., Loda, M., and Rollins, B. J. (2000). Control of TH2 polarization by the chemokine monocyte chemoattractant protein-1. Nature 404, 407-411.
Ho, I. C., and Glimcher, L. H. (2002). Transcription: tantalizing times for T cells. Cell 109 Suppl, S109-120.
Ho, I. C., Hodge, M. R., Rooney, J. W., and Glimcher, L. H. (1996). The
proto-oncogene c-maf is responsible for tissue-specific expression of interleukin-4. Cell 85, 973-983.
Ho, I. C., Vorhees, P., Marin, N., Oakley, B. K., Tsai, S. F., Orkin, S. H., and Leiden, J. M. (1991). Human GATA-3: a lineage-restricted transcription factor that regulates the expression of the T cell receptor alpha gene. EMBO J 10, 1187-1192.
Kageyama, Y., Koide, Y., Yoshida, A., Uchijima, M., Arai, T., Miyamoto, S., Ozeki, T., Hiyoshi, M., Kushida, K., and Inoue, T. (1998). Reduced susceptibility to
collagen-induced arthritis in mice deficient in IFN-gamma receptor. J Immunol 161, 1542-1548.
required for mediating responses to IL-4 and for development of Th2 cells. Immunity 4, 313-319.
Kaplan, M. H., Whitfield, J. R., Boros, D. L., and Grusby, M. J. (1998a). Th2 cells are required for the Schistosoma mansoni egg-induced granulomatous response. J
Immunol 160, 1850-1856.
Kaplan, M. H., Wurster, A. L., and Grusby, M. J. (1998b). A signal transducer and activator of transcription (Stat)4-independent pathway for the development of T helper type 1 cells. J Exp Med 188, 1191-1196.
Kim, J. I., Ho, I. C., Grusby, M. J., and Glimcher, L. H. (1999). The transcription factor c-Maf controls the production of interleukin-4 but not other Th2 cytokines. Immunity 10, 745-751.
Ko, L. J., Yamamoto, M., Leonard, M. W., George, K. M., Ting, P., and Engel, J. D. (1991). Murine and human T-lymphocyte GATA-3 factors mediate transcription
through a cis-regulatory element within the human T-cell receptor delta gene enhancer. Mol Cell Biol 11, 2778-2784.
Kuperman, D., Schofield, B., Wills-Karp, M., and Grusby, M. J. (1998). Signal transducer and activator of transcription factor 6 (Stat6)-deficient mice are protected from antigen-induced airway hyperresponsiveness and mucus production. J Exp Med 187, 939-948.
Miaw, S. C., Choi, A., Yu, E., Kishikawa, H., and Ho, I. C. (2000). ROG, repressor of GATA, regulates the expression of cytokine genes. Immunity 12, 323-333.
Miaw, S.-C., Kang, B., White, I., and Ho, I. C. (2004). A repressor of GATA-mediated negative feedback mechanism of T cell activation. J Immunol 172.
Mosmann, T. R., Cherwinski, H., Bond, M. W., Giedlin, M. A., and Coffman, R. L. (1986). Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J Immunol 136, 2348-2357.
Mosmann, T. R., and Coffman, R. L. (1989). TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol 7, 145-173.
Pai, S. Y., Truitt, M. L., Ting, C. N., Leiden, J. M., Glimcher, L. H., and Ho, I. C. (2003). Critical roles for transcription factor GATA-3 in thymocyte development. Immunity 19, 863-875.
Pandolfi, P. P., Roth, M. E., Karis, A., Leonard, M. W., Dzierzak, E., Grosveld, F. G., Engel, J. D., and Lindenbaum, M. H. (1995). Targeted disruption of the GATA3 gene causes severe abnormalities in the nervous system and in fetal liver haematopoiesis. Nat Genet 11, 40-44.
Sherman, M. A., Nachman, T. Y., and Brown, M. A. (1999). Cutting edge: IL-4 production by mast cells does not require c-maf. J Immunol 163, 1733-1736.
Szabo, S. J., Dighe, A. S., Gubler, U., and Murphy, K. M. (1997). Regulation of the interleukin (IL)-12R beta 2 subunit expression in developing T helper 1 (Th1) and Th2 cells. J Exp Med 185, 817-824.
Szabo, S. J., Kim, S. T., Costa, G. L., Zhang, X., Fathman, C. G., and Glimcher, L. H. (2000). A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell 100, 655-669.
Szabo, S. J., Sullivan, B. M., Stemmann, C., Satoskar, A. R., Sleckman, B. P., and Glimcher, L. H. (2002). Distinct effects of T-bet in TH1 lineage commitment and IFN-gamma production in CD4 and CD8 T cells. Science 295, 338-342.
Specific aim 2: To study the role of the transcription factor Ets-1 in immune response
Subjects and Methods
Clone full length Mus musculus Ets-1 cDNA
The coding region was amplified from a pFlag-Ets1 plasmid (a gift from Dr. I-Chen Ho)(Grenningloh et al., 2005) by polymerase chain reaction (PCR). Because the lack of Ets-1 start codon in the pFlag-Ets-1 plasmid, we added the additional nucleotides ATG in the forward primer to rebuild the complete Ets-1 cDNA. In addition to the start codon, we introduced restriction endonuclease sites in the designed primers for manipulation. The restriction endonuclease sites we choose in this pair of primers are Sal I sites.
Forward primer: 5’-TATAGTCGACATGAAGGCGGCCGTCGATCT-3’ Reverse primer: 5’-TTCGGTCGACCTAGTCAGCATCCGGCTTTA-3’.
The PCR amplified full-length Ets-1 cDNA fragments were digested by Sal I and clone into pBluescript II KS(-) vector (Stratagene, USA) and was designated pBSIIKS(-)-Ets1(1323). The construct was further confirmed by restriction enzyme mapping and sequencing.
Construction of Ets-1 minimal reporter for luciferase assay
The ETS binding sequence (EBS) specific for Ets-1 was designed based on the previous study (Reddy et al., 2003). The designed binding sequence contains two copies of EBS and flanked by XhoI restriction enzyme sites.
Forward primer 5’-CTAGCCTCGAGTGCCGGAAATGTAGTGCCGGAAATGT-3’ Reverse primer 5’-TCGAACATTTCCGGCACTACATTTCCGGCACTCGAGG-3’ The synthesized primers were annealed and cloned into the luciferase vector pGL3 (Promega) to generate 2XEBS-luciferase reporter and 4XEBS-luciferase reporter. The two constructs were further confirmed by sequencing.
Clone GST fusion Ets-1 for polyclonal antibody production
To purify Ets-1 protein for the production of polyclonal antibody, I chose GST fusion protein system for this purpose. Because the homology of the ETS DNA binding domain within ETS family proteins, I used a truncated form of Ets-1 (999bp) to generate the GST-Ets1(999) fusion protein to eliminated the cross activities of Ets-1 antiserum. The 999 base pair Ets-1 was cloned from pBSIIKS(-)-Ets1(1323) by forward primer 5’-TATAGTCGACATGAAGGCGGCCGTCGATCT-3’ reverse primer 5’-CACAGGTCGACCTACCCACTTCCTGTGTAGCC-3’ primer pairs and cloned into pGEX-4T-1 vector (Amersham Biosciences, USA) to produce
sequencing.
Clone full length Mus musculus PIAS1, PIAS3, PIASx and PIASy
Full length PIAS1, PIAS3, PIASx and PIASy were amplified by PCR from cDNA library of type 1 T helper cell or CD8 T cell and cloned into pcDNA3.1 (Invitrogen) expression vector.
Primer design For PIAS1
Forward primer 5’-TTCTCGAGAGATGGCGGACAGTGC-3’
Reverse primer 5’-ACCTCGAGTCAGTCCAATGAGATAATGTCT-3’ For PIAS3
Forward primer 5’-ACCTCGAGACATGGTGATGAGTTTCC-3’ Reverse primer 5’-GACTCGAGTCAGTCCAAGGAAATGAC-3’ For PIASx
Forward primer 5’-ATCTCGAGAAATGGCGGATTTCGAGGAG-3’ Reverse primer 5’-CCCTCGAGTTAGTCCAAAGAGATGATGTCA-3’ For PIASy
Forward primer 5’-GCGAATTCATGGCGGCAGAGCTGG-3’ Reverse primer 5’-TACTCGAGTCAGCACGCGGGCAC-3’ Sumoylation sites prediction
Typically lysine residues subject to SUMO modification were found within a SUMO modification consensus motif, KxE (where is a large hydrophobic residue and x is any residue). Full length Ets-1 cDNA sequence was searched for the sumoylation sites. The sites which might be modified by SUMO-1 were predicted by ELM program (http://elm.eu.org) and SUMOplotTM program (http://www.abgent.com). Four putative sumoylation sites were found which located at amino acid 14-17, 199-202, 226-229 and 435-438 of Ets-1.
Site-directed mutagenesis
The four conserved lysine residues in predicted SUMO-1 interacting sites of Ets-1 were changed to arginine by site-directed mutagenesis (QuikChange® Site-Directed Mutagenesis Kit, Stratagene). Using the pBSIIKS(-)-Ets1 (1323bp) as the template, we generated four Ets-1 mutants.
Primers design sumoylation site 1 (K15R) Forward primer 5’-CGACTCTCACCATCATCAGGACAGAAAAAGTGGATCTCG-3’ Reverse primer 5’-CGAGATCCACTTTTTCTGTCCTGATGATGGTGAGAGTCG-3’ sumoylation site 2 (K200R)
Forward primer 5’-CGGAAGAACTCCTGTCCCTCAGGTATGAGAACGACTACCC-3’ Reverse primer 5’-GGGTAGTCGTTCTCATACCTGAGGGACAGGAGTTCTTCCG-3’ sumoylation site 3 (K227R) Forward primer 5’-GCAGACAGACTACTTTGCCATCAGGCAAGAGGTGTTAACTCC-3’ Reverse primer 5’-GGAGTTAACACCTCTTGCCTGATGGCAAAGTAGTCTGTCTGC-3’ sumoylation site 4 (K436R)
Forward primer 5’-GCTGCACGCCATGCTGGATGTAAGGCCGGATGC-3’ Reverse primer 5’-GCATCCGGCCTTACATCCAGCATGGCGTGCAGC-3’ The constructs were checked by sequencing.
The wild type and four sumoylation site mutants of Ets-1 were then cut from pBSIIKS(-)-Ets1(1323), pBSIIKS(-)-Ets1.K15R, pBSIIKS(-)-Ets1.K200R,
pBSIIKS(-)-Ets1.K227R and pBSIIKS(-)-Ets1.K436R, respectively and cloned into the multiple cloning site of pcDNA3.1, a mammalian expression vector. The
constructs were checked by restriction enzyme mapping and sequencing. HEKHEK293T cells transfection
All the transfections were done by using the FuGENE 6 Transfection Reagent (Roche) according to the manufacturer’s instructions.
Luciferase assay
One day before transfection, HEKHEK293T cells were seeded into 24-well cell culture plate, 105/well. On the day of transfection, the DNA mixtures were prepared and transfected into HEK293T cells by FuGENE 6 Transfection Reagent (Roche). One day after the transfection, the culture medium of each well was replaced by 50 μl serum free medium/Dual-GloTM Luciferase Reagent (Promega) 1:1 mixture. Ten minutes later, the cells were lysed and 40 μl of the cell lysates were transfer to 96-well black plate. After 10 minutes waiting, the firefly luminescence was measured by luminometer. Then 20 μl Dual-GloTM Stop & Glo® Reagent was also added to each
well and the Renilla luminescence was measured after 10 minutes incubation. The firefly luminescence of luciferase was normalized by the luminescence of Renilla following the equation: luminescence of firefly/luminescence of Renilla. The relative luciferase activities were determined and subjected to statistic analysis.
Results
Ets-1 is comparable
Previous study (Grenningloh et al., 2005) had shown that with the help of Ets-1, T-bet can transactivate IFNγ promoter more efficiently. Since Ets-1 can interact with UBC9 (Hahn et al., 1997) and can be sumoylated in vitro (Macauley et al., 2006), this sumoylation reaction may be involved in the regulation of the functional synergy between T-bet and Ets-1.
To investigate the effect of sumoylaton on Ets-1 in T-bet transactivate IFNγ promoter, we mutated four predicted sumoylation sites within Ets-1 base on database search for the well known SUMO consensus sequence. Then lucifease assay was performed to examine the effects of the mutated Ets-1.
When we cotransfect T-bet with wild type or mutated Ets-1 in a IFN
promoter-luciferase system, the effects caused by four kinds of mutated Ets-1 were similar to wild type Ets-1 and showed no significant difference.
These results suggested that sumoylation of Ets-1 may not be involved in transactvating IFNγ promoter by T-bet.
The role of PIAS proteins in IFNγ transactivation by T-bet
The PIAS family proteins are the major E3 ligase for sumoylation reaction and probably determine the specificity of sumoylation. Some researches have indicated that PIAS proteins alone can complete this sumoylation reaction. Since Ets-1 could be sumoylated in vivo, it’s interesting to examine the functional synergy between Ets-1 and T-bet in the presence of PIAS proteins.
Our luciferase reporter experiments showed that the transcriptional activity of T-bet in IFNγ-promoter reporter system was blocked in the presence of hPIASy, mPIAS1, mPIAS3 and mPIASxβ and the synergy effect by sumoylation site mutated Ets-1 were comparable to wild type Ets-1.
According to our experiment, the PIAS proteins can inhibit the transactivating function of T-bet. But whether the PIAS proteins can interact with Ets-1 and further influence the Ets-1 function remains to be clarify and can not be tested in the IFN reporter luciferase system.
The Ets-1 minimal reporter luciferase assay system
Sumoylation may influence the protein-protein interactions and protein-DNA binding affinities. In order to address this issue, we constructed an Ets-1 minimal reporter system which can be specifically bound by Ets-1 (ETS-specific binding sequence, EBS) and transactivate downstream gene, luciferase. We generated 2XEBS-luciferase and 4XEBS-luciferase reporters and performed the luciferase assay in the presence of different dosage of wild type Ets-1. Both the 2XEBS-Luciferase reporter and
4XEBS-Luciferase reporter had the abilities to be transactivated by wild-type Ets-1 and showed a dosage dependent manner. Since the responsiveness of the
2XEBS-luciferase reporter was more sensitive than 4XEBS-Luciferase reporter and a much reduced background, we chose 2XEBS-Luciferase reporter for further
experiments.
The transactivation abilities of wild type or sumoylation sites mutated Ets-1
The 2XEBS-Luciferase reporter is useful for directly examining the transcriptional activities of Ets-1 sumoylaiton site mutants. We cotransfected wild type or mutated Ets-1 and 2XEBS-Luciferase reporter in HEKHEK293T cells and luciferase assay was performed. The results showed that wild type and Ets-1 mutants can induce similar luciferase activities. We concluded that the transcriptional activities of Ets-1 sumoylation site mutants were comparable to wild type Ets-1 in 2XEBS-luciferase system.
The transcriptional activities of wild type Ets-1 in the presence of PIAS proteins. Next, we examined the transactivation ability of Ets-1 in the presence of PIAS family proteins. We cotransfected mEts-1 with hPIASy, mPIAS1, mPIAS3, mPIASx and mPIASx into HEKHEK293T cells and proceed luciferase assay Interestingly, the results showed that hPIASy, mPIAS1, mPIAS3 and mPIASxβ had negative effects on Ets-1 in 2XEBS-luciferase system but mPIASxα can increase the transactivity of Ets-1. These data indicated that PIAS family proteins may play some roles in the regulation of Ets-1.
The transcriptional activities of Ets-1 mutants in the presence of PIAS family proteins .
The modified transcriptional activities of Ets-1 in 2XEBS-Luciferase reporter experiment caused by PIAS proteins could be a result of sumoylation on Ets-1 since PIAS proteins are E3 ligase of sumoylation reaction. We can answer this question by introducing sumoylation sites mutants of Ets-1 into the 2XEBS-Luciferase reporter system. In the presence of PIAS proteins, when wild type Ets-1 was replaced by mutated Ets-1, these mutants can not rescue the effects caused by PIAS proteins. We concluded that the reduced transcriptional activities of Ets-1 in 2XEBS-luciferase reporter system caused by hPIASy, mPIAS1, mPIAS3 and mPIASxβ or the increased activity of Ets-1 caused by mPIASxα were not related to the Ets-1 sumoylation site mutants.
Discussions
Traditionally, Ets-1 was recognized as a tumor oncogene and the majority of previous researches were focus on the roles of Ets-1 in embryogenesis and tumorigenesis. The role of Ets-1 in T helper cells has been described recently (Grenningloh et al., 2005). Ets-1 knockout type 1 T helper cells showed defects in the production of Th1
cytokines such as interleukin-2, interferon-γ and an upregulation of interleukin-10, which is a kind of Th2 cytokines. Further experiments showed that T-bet can
transactivate IFNγ more efficiently with the help of Ets-1. Since Ets-1 plays a cofactor role in T-bet transactivating IFNγ the regulation of Ets-1 may play a fine tune
mechanism with the Th1 major transcriptional factor, T-bet.
In our experiments, when we mutated the sumoylation sites of Ets-1, the T-bet helping effects of Ets-1 remained unchanged. Furthermore, when we added the SUMO
reaction E3 ligase, PIAS family proteins, the PIAS proteins had the abilities to reduce the transcriptional activity of T-bet significantly. However, Ets-1 seem has no roles in this inhibition effects. Two reasonable speculations are that PIAS proteins can directly effect the transcriptional activity of T-bet or the sumoylation reaction has no effects on Ets-1 incooperation with T-bet to transactivate IFNγ In other words, this reporter system is not suitable for examine the role of sumoylation on Ets-1.
Therefore, we examine directly the transcriptional activity of Ets-1 by sumoylation. The question we would like to ask is whether sumoylation played a role in affecting the transcriptional activity of Ets-1. Therefore, we designed an Ets-1 binging
sequence which can be specifically bound by Ets-1 and this minimal promoter can be used to transactivate downstream luciferase gene. The results showed that although we mutated the sumoylation sites within Ets-1, the transcriptional activities were very similar between wild type and mutant Ets-1. These results indicated the transcriptional activities of these Ets-1 sumoylation sites mutants have been unchanged. Sumoylation may play no roles in regulating the transcriptional activity of Ets-1. Other possibility is that the sumoylation machinery in HEKHEK293T cells is not enough or incomplete to cause significant effects. To strengthen the sumoylation reactions of transfected cells, we cotransfected PIAS proteins with wild type or mutated Ets-1. PIAS proteins alone can induce strong sumoylation reactions according to papers published before. In our experimental results, cotransfected wild type Ets-1 with PIAS proteins can significantly reduce or increase the activity of Ets-1. These results indicated that Ets-1 may interact with PIAS proteins directly or indirectly. However, it is possible that those PIAS proteins can directly interact with the Ets-1 minimal reporter. In our luciferase vector, there is a TATA binding sequence prior to the start codon of luciferase gene which is responsible for the basal transcriptional activity. There was paper reported that PIAS proteins can interact with TATA binding proteins (Prigge and Schmidt, 2006). This raises the possibility that PIAS proteins can directly affect the luciferase vector via the TATA binding proteins. The electro-mobility shift assay (EMSA) is needed to resolve this question or we, alternatively, can delete the TATA box of the vector and perform luciferase assay. By using the mutated Ets-1 together with PIAS proteins, the relative luciferase activities remained unchanged compare
with wild type Ets-1.
One issue in our experiment need to be addressed is that all the transfection experiments were done in the HEK293T cell line. HEK293T cell line is a human kidney epithelial cell line and may not completely mimic the T cell environment. The best way to study T cell functions is to use the primary T cells for this study. We can use the primary T cells from Ets-1 deficient mice to study the function of Ets-1 sumoylation sites mutants. We can put the wild type or mutated Ets-1 back to the Ets-1 KO T cells, and examines the functions of Ets-1 by their cytokines secreting profiles.
In our assay system, we cannot see any difference between the wild type and sumoylation sites mutated Ets-1 in terms of transactivation ability. One possible explanation is that the sumoylation sites of Ets-1 are not within the classical xE motif. Although one of the consensus sumoylaiton sites has been demonstrated as the major SUMO binding site, we cannot rule out the possibilities that other sumoylation sites play minor but important roles.
In order to demonstrate the relation between Ets-1 and sumoylation reaction, further experiments still need to be done. First, we need to demonstrate the interaction between Ets-1 and UBC9, the only identified SUMO E2 conjugating enzyme. Although the paper published in 1997 has proved the relationship between Ets-1 by yeast two-hybrid system and GST pull down assay, we still need to confirm the results by our yeast two-hybrid system and coimmunoprecipitation assay. On the other hand, since the SUMO specificity is determined by the SUMO E3 ligase, the PIAS proteins or other proteins, we need to define the E3 ligase specific for the sumoylation reaction of Ets-1. Similarly, yeast two-hybrid and coimmunoprecipitation assays are needed to confirm this speculation. Furthermore, the in vivo sumoylation of Ets-1 have to be confirmed. Three strategies can be used to detect the in vivo sumoylation of Ets-1. First, we can cotransfect Ets-1 with the basic sumoylation machinery, such as E1 activating enzymes and E2 conjugation enzyme, then we can immunoprecipitate Ets-1 and detect the sumoylation levels by standard Western blot analysis using SUMO antibodies in a easy transfected cell line such as HEK293T cells. Next, we have to perform same experiment with T cell line such as EL4 cell line. Finally, we need to apply the primary T cells to confirm the sumoylation with Ets-1, and this can be achieved by the retroviral transduction of wild type or sumoylation site mutated Ets-1 in to Ets-1-deficient T cells.
References
Barton, K., Muthusamy, N., Fischer, C., Ting, C. N., Walunas, T. L., Lanier, L. L., and Leiden, J. M. (1998). The Ets-1 transcription factor is required for the development of natural killer cells in mice. Immunity 9, 555-563.
Bories, J. C., Willerford, D. M., Grevin, D., Davidson, L., Camus, A., Martin, P., Stehelin, D., and Alt, F. W. (1995). Increased T-cell apoptosis and terminal B-cell differentiation induced by inactivation of the Ets-1 proto-oncogene. Nature 377, 635-638.
Dittmer, J. (2003). The biology of the Ets1 proto-oncogene. Mol Cancer 2, 29.
Ghysdael, J., Gegonne, A., Pognonec, P., Dernis, D., Leprince, D., and Stehelin, D. (1986). Identification and preferential expression in thymic and bursal lymphocytes of a c-ets oncogene-encoded Mr 54,000 cytoplasmic protein. Proc Natl Acad Sci U S A 83, 1714-1718.
Grenningloh, R., Kang, B. Y., and Ho, I. C. (2005). Ets-1, a functional cofactor of T-bet, is essential for Th1 inflammatory responses. J Exp Med 201, 615-626.
Johnson, E. S. (2004). Protein modification by SUMO. Annu Rev Biochem 73, 355-382.
Kola, I., Brookes, S., Green, A. R., Garber, R., Tymms, M., Papas, T. S., and Seth, A. (1993). The Ets1 transcription factor is widely expressed during murine embryo development and is associated with mesodermal cells involved in morphogenetic processes such as organ formation. Proc Natl Acad Sci U S A 90, 7588-7592.
Lavender, P., Cousins, D., and Lee, T. (2000). Regulation of Th2 cytokine gene transcription. Chem Immunol 78, 16-29.
Li-Weber, M., and Krammer, P. H. (2003). Regulation of IL4 gene expression by T cells and therapeutic perspectives. Nat Rev Immunol 3, 534-543.
Macauley, M. S., Errington, W. J., Scharpf, M., Mackereth, C. D., Blaszczak, A. G., Graves, B. J., and McIntosh, L. P. (2006). Beads-on-a-string, characterization of ETS-1 sumoylated within its flexible N-terminal sequence. J Biol Chem 281, 4164-4172.
Mackereth, C. D., Scharpf, M., Gentile, L. N., MacIntosh, S. E., Slupsky, C. M., and McIntosh, L. P. (2004). Diversity in structure and function of the Ets family PNT domains. J Mol Biol 342, 1249-1264.
Mullen, A. C., High, F. A., Hutchins, A. S., Lee, H. W., Villarino, A. V., Livingston, D. M., Kung, A. L., Cereb, N., Yao, T. P., Yang, S. Y., and Reiner, S. L. (2001). Role of T-bet in commitment of TH1 cells before IL-12-dependent selection. Science 292, 1907-1910.
structure of the avian erythroblastosis virus E26 transforming gene. Nature 306, 391-395.
Obika, S., Reddy, S. Y., and Bruice, T. C. (2003). Sequence specific DNA binding of Ets-1 transcription factor: molecular dynamics study on the Ets domain--DNA complexes. J Mol Biol 331, 345-359.
Ouyang, W., Lohning, M., Gao, Z., Assenmacher, M., Ranganath, S., Radbruch, A., and
Prigge, J. R., and Schmidt, E. E. (2006). Interaction of protein inhibitor of activated STAT (PIAS) proteins with the TATA-binding protein, TBP. J Biol Chem 281, 12260-12269.
Reddy, S. Y., Obika, S., and Bruice, T. C. (2003). Conformations and dynamics of Ets-1 ETS domain-DNA complexes. Proc Natl Acad Sci U S A 100, 15475-15480. Seeler, J. S., and Dejean, A. (2003). Nuclear and unclear functions of SUMO. Nat Rev Mol Cell Biol 4, 690-699.
Sementchenko, V. I., and Watson, D. K. (2000). Ets target genes: past, present and future. Oncogene 19, 6533-6548.
Teruyama, K., Abe, M., Nakano, T., Iwasaka-Yagi, C., Takahashi, S., Yamada, S., and Sato, Y. (2001). Role of transcription factor Ets-1 in the apoptosis of human vascular endothelial cells. J Cell Physiol 188, 243-252.