BIOLOGIA PLANTARUM 51 (2): 351-354, 2007
351
BRIEF COMMUNICATIONProline accumulation induced by excess nickel in detached rice leaves
Y.C. LIN and C.H. KAO*Department of Agronomy, National Taiwan University, Taipei 106, Taiwan, Republic of China
Abstract
The regulation of proline accumulation in detached rice leaves exposed to excess NiSO4 was investigated. NiSO4
treatment increased proline and Ni contents but had no effect on relative water content, indicating that proline accumulation in Ni-exposed detached rice leaves was due to Ni uptake per se, rather than to water stress. Proline accumulation caused by NiSO4 was related to protein hydrolysis, a decrease in proline dehydrogenase activity, and a
decrease in proline utilization. It seems that an increase in the content of ammonia and an increase in the activities of Δ1-pyrroline-5-carboxylate reductase and ornithine-δ-aminotransferase play minor if any role in Ni-induced proline
accumulation in detached rice leaves.
Additional key words: glutamic acid, ornithine, ornithine-δ-aminotransferase, Oryza sativa, proline dehydrogenase, Δ1-pyrroline- 5-carboxylate reductase.
⎯⎯⎯⎯
Nickel (Ni) is an essential element for plant growth (Brown et al. 1987). In general, there is much more concern about Ni toxicity in crop plants. Critical toxicity levels in crop species are in the range of > 10 μg g-1(d.m.)
in sensitive, and 50 μg g-1(d.m.) in moderately tolerant
species (Marschner 1995). At toxic concentrations Ni interferes with plant growth and metabolism (Mishra and Kar 1974). Some plant species are adapted to grow on soil containing high content of Ni. Such plants were named Ni hyperaccumulators. Recently, shoot cultures of
Alyssum markgraffi, endemic Ni hyperaccumulating
species, were established and maintained on Murashige and Skoog medium (Vinterhalter and Vinterhalter 2005). Nakazawa et al. (2004) demonstrated that histidine is involved in Ni-tolerance and detoxification of Ni in symplast in Ni-tolerant tobacco cells.
Many plants accumulate proline when treated with toxic concentrations of heavy metals (Alia and Pardha Saradhi 1991, Bassi and Sharma 1993a,b, Costa and Morel 1994, Schat et al. 1997, Metha and Gaur 1999, Chen et al. 2001, Rai 2002). Metha and Gaur (1999) reported that excess Ni caused proline accumulation in
Chlorella vulgaris. However, this is the only report
describing proline accumulation under Ni stress so far. It
is not known whether other plant systems also show proline accumulation in response to Ni.
Proline accumulation in plant tissues has been suggested to result from 1) an increase in proline biosynthesis, 2) a decrease in proline degradation, 3) an increase in protein hydrolysis, 4) a decrease in proline utilization (Charest and Phan 1990, Yoshiba et al. 1997). In plants, proline is synthesized from glutamic acid via Δ1-pyrroline-5-carboxylate (P5C) by two enzymes, P5C
synthetase and P5C reductase (P5CR, EC 1.5.1.2) (Delauney and Verma 1993,Yoshiba et al. 1997). Plants also synthesize proline from ornithine by ornithine-δ- aminotransferase (OAT, EC 2.6.1.13). On the other hand, the content of proline also depends on its degradation, which is catalysed by the enzyme proline dehydrogenase (PDH, EC 11.5.99.8) (Yoshiba et al. 1997).
In detached rice leaves, it has been shown that proline accumulates during dark-induced senescence (Wang et al. 1982), by excess copper (Chen et al. 2001), by NaCl (Lin
et al. 2002), by water stress (Hsu et al. 2003, Yang and
Kao 1999), and by exogenous abscisic acid (Yang et al. 2000). This paper reports the results of an investigation into the regulation of proline accumulation in detached rice leaves exposed to excess NiSO4.
⎯⎯⎯⎯
Received 10 June 2005, accepted 25 March 2006.
Abbreviations: d.m. - dry mass; f.m. - initial fresh mass; GABA - γ-aminobutyric acid; OAT - ornithine-δ-aminotransferase; P5C - Δ1-pyrroline-5-carboxylate; P5CR - Δ1-pyrroline-5-carboxylate reductase; PDH - proline dehydrogenase; RWC - relative water content.
Acknowledgement: This work was supported by the National Science Council of the Republic of China. * Corresponding author; fax: (+886) 02 23620879, e-mail: [email protected]
Y.C. LIN, C.H. KAO
352
Rice (Oryza sativa L. cv. Taichung Native 1) was cultured in a stainless net floating on Kimura B nutrient solution (pH 4.2) in a 500-cm3 beaker (Hsu and Kao
2005). The nutrient solution was replaced every 3 d. Rice plants were grown for 12 d in a greenhouse, under natural sunlight and the day/night temperature of 30/25 °C. The apical 3 cm of the third leaf of 12-d-old seedlings was used for the experiment. A group of 10 segments floated in a Petri dish containing 10 cm3 of distilled water served
as controls and NiSO4 (10 mM). All samples were kept at
temperature at 27 °C and irradiance of 40 μmol m-2 s-1 for
12, 24, 36, and 48 h.
Relative water content (RWC) was determined by the method of Weatherley (1950). For determination of proline, glutamic acid, ornithine, γ-aminobutyric acid (GABA), NH3, and total amino acids, leaf samples were
extracted with 2 % sulfosalicylic acid and the homo-genates was centrifuged at 15 000 g for 20 min. The supernatant was used directly for amino acid analysis (amino acid analyzer, Beckman 63000, Palo Alto, USA). For protein determination, leaf segments were homoge-nized in 50 mM sodium phosphate buffer (pH 6.8). The homogenate was centrifuged at 17 600 g for 20 min, and the supernatants were used for determination of protein by the method of Bradford (1976). Enzymes were extracted and assayed as described previously (Chen
et al. 2001). For determination of Ni, leaf samples were
dried at 65 °C for 48 h. Dried materials were ashed at 550 °C for 4 d. Ash residues were incubated with 17.5 % (v/v) HNO3 and 17.5 % (v/v) H2O2 at 72 °C for more than
10 h, and dissolved in double distilled water for 3 h. Ni content was then quantified using an atomic absorption spectrophotometer (Shimadzu AA-6800, Kyoto, Japan).
Amino acids, protein, and NH3 were expressed on the
basis of initial fresh mass (f.m.). Enzyme activity was expressed on the basis of protein. Ni was expressed on the basis of dry mass (d.m.). Statistical differences between measurements (n = 4) for different treatments or for different times were analyzed following the Duncan’s multiple range test or Student’s t-test.
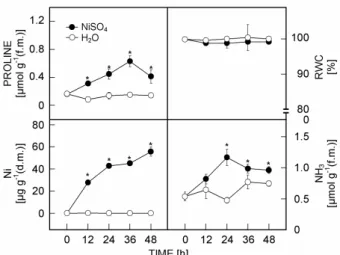
Proline content in detached rice leaves exposed to NiSO4 (10 mM) increased significantly with the increase
of incubation time (Fig. 1). It is clear that accumulation of proline induced by NiSO4 was evident 12 h after
treatment (Fig. 1).
To be sure that the described proline accumulation was related to an increase in the leaf Ni content, Ni concentrations were determined in detached rice leaves treated with either water or 10 mM NiSO4 (Fig. 1). Ni
content in control leaves remained unchanged during 48 h in the light. However, Ni content in NiSO4-treated
detached rice leaves increased with increasing duration of incubation. It is obvious that the increase in Ni content in NiSO4-treated detached rice leaves was evident 12 h after
treatment.
Rabe (1990) postulated that any stress causing reduced growth or impaired plant health will result in ammonia accumulation early and suggested that the
ammonia detoxification results in the accumulation of nitrogen containing compounds, such as putrescine and proline. Exogenous NH4Cl, methionine sulfoximine and
phosphinothricin, which caused an accumulation of ammonia in detached rice leaves, increased proline content (Yang and Kao 2000, Tsai et al. 2003). In the present study, we also observed that NiSO4 treatment
resulted in an ammonia accumulation (Fig. 1). However, Ni-induced ammonia accumulation was observed only after the onset of proline accumulation (Fig. 1). It appears that ammonia plays a minor if any role in proline accumulation by NiSO4 in detached rice leaves.
Fig. 1. The contents of proline, Ni, and ammonia and relative water content (RWC) in detached rice leaves floating on water (H2O) or NiSO4 (10 mM) solution for 12, 24, 36, and 48 h. Means ± SD, n = 4. Asterisks represent values that are significantly different between H2O and NiSO4 treatments at
P < 0.05 determined by Student’s t-test.
Fig. 2. The contents of total amino acids and protein in detached rice leaves floating on water (H2O) or NiSO4 (10 mM) solution for 12, 24, 36, and 48 h. Means ± SD, n = 4. Asterisks represent values that are significantly different between H2O and NiSO4 treatments at P < 0.05 determined by Student’s t-test.
PROLINE ACCUMULATION INDUCED BY NICKEL
353
No differences in RWC were observed betweenNi-treated leaves and control leaves (Fig. 1). It appears that Ni-induced proline accumulation is not attributed to the development of water stress. Previously, we also observed that proline accumulation in Cu-exposed detached rice leaves and argued that this was due to Cu uptake per se, rather than to water stress (Chen et al. 2001).
The decrease in protein content was faster in NiSO4-treated detached rice leaves than in control leaves
(Fig. 2). Therefore, protein degradation might contribute to NiSO4-induced proline accumulation in detached rice
leaves. This suggestion is supported further by the observation that the content of total amino acids was higher in detached rice leaves exposed to NiSO4 solution
than in control leaves (Fig. 2). It is generally considered that glutamic acid and ornithine can contribute to the accumulation of proline (Delauney and Verma 1993). The decreased or low content of glutamic acid was observed in NiSO4-treated detached rice leaves (Fig. 3),
suggesting that glutamic acid is converted to proline in rice leaves exposed to excess NiSO4. Since
γ-amino-butyric acid (GABA) content in NiSO4-treated detached
rice leaves was observed to be higher than those in water-treated detached rice leaves (Fig. 3), therefore the decreased content of glutamic acid also resulted from glutamic acid being metabolized to GABA (Ireland and Lea 1999). The increase in glutamine content and decrease in glutamic acid in NiSO4-treated detached rice
leaves (Fig. 3) also suggest that NiSO4 inhibits the
conversion of glutamine to glutamic acid, a step catalyzed by glutamic acid synthase (Ireland and Lea 1999). Although ornithine may contribute to the accumulation of proline (Delauney and Verma 1993), slightly higher content of ornithine in NiSO4-treated detached rice leaves
was observed to occur after proline accumulation (Fig. 3).
Fig. 3. The contents of glutamic acid, glutamine, γ-aminobutyric acid (GABA), and ornithine in detached rice leaves floating on water (H2O) or NiSO4 (10 mM) solution for 12, 24, 36, and 48 h. Means ± SD, n = 4. Asterisks represent values that are significantly different between H2O and NiSO4 treatments at
P<0.05 determined by Student’s t-test.
It appears that ornithine plays no important role in Ni-induced proline accumulation in rice leaves.
To determine the role of biosynthetic pathway for proline accumulation caused by NiSO4, the effect of
NiSO4 on P5CR and OAT activities was examined. It is
obvious that clear differences in P5CR and OAT activities between NiSO4- and water-treated detached rice
leaves was observed at the later stage of incubation (Fig. 4), suggesting that P5CR and OAT play minor role in proline accumulation induced by NiSO4. It has been
reported that PDH catalyzed proline oxidation (Hare et al. 1999). In the present study, PDH activity was found to decrease significantly in detached rice leaves exposed to excess Ni (Fig. 4), suggesting that decrease in proline oxidation (or degradation) contributes to proline accumulation in detached rice leaves under NiSO4 stress
condition.
Fig. 4. The activities of P5CR, OAT, and PDH in detached rice leaves floating on water (H2O) or NiSO4 (10 mM) solution for 12, 24, 36, and 48 h. Means ± SD, n = 4. Asterisks represent values that are significantly different between H2O and NiSO4 treatments at P < 0.05 determined by Student’s t-test.
A decrease in proline utilization may also contribute to the accumulation of proline in NiSO4-treated rice
leaves. To test this possibility, detached rice leaves were pretreated with 50 mM ornithine for 3 h to increase the endogenous proline content and then transferred to water and NiSO4 for 24 h in the light; proline contents were
then determined. It was found that the proline content was lower in the absence of NiSO4 than in the presence of
NiSO4 (data not shown), suggesting that proline in
detached rice leaves treated with NiSO4 is utilized less
Y.C. LIN, C.H. KAO
354
in detached rice leaves by NaCl has also been described previously (Lin et al. 2002).
The physiological significance of proline accumula-tion in intact rice leaves is not fully understood. Studying the effect of NiSO4 stress on enzyme activities involved
in proline metabolism in intact rice leaves could provide valuable information on the physiological significance of
its accumulation. Regulation of proline accumulation in detached rice leaves under NiSO4 stress as we reported
here is not necessarily similar to that in intact rice leaves. However, the results of the present work do provide some basic information which should be valuable for our future studies
References
Ali, Pardha Saradhi, P.: Proline accumulation under heavy metal stress. - J. Plant Physiol. 138: 504-508, 1991.
Bassi, R., Sharma, S.S.: Changes in proline content accompanying the uptake of zinc and copper by Lemna minor. - Ann. Bot. 72: 151-154, 1993a.
Bassi, R., Sharma, S.S.: Proline accumulation in wheat seedlings exposed to zinc and copper. - Phytochemistry 33: 1339-1342, 1993b.
Bradford, M.M.: A rapid and sensitive method for the quantitation of microgram quantities for protein utilizing the principles of protein-dye binding. - Anal. Biochem. 72: 248-254, 1976.
Brown, P.H., Welch, R.M., Cary, E.E.: Nickel: a micronutrient essential for higher plants. - Plant Physiol. 85: 801-803, 1987.
Charest, C., Phan, C.T.: Cold acclimation of wheat (Triticum aestivum): properties of enzymes involved in proline metabolism. - Physiol. Plant. 80: 159-168, 1990.
Chen, C.T., Chen, L.M., Lin, C.C., Kao, C.H.: Regulation of proline accumulation in detached rice leaves exposed to excess copper. - Plant Sci. 160: 283-290, 2001.
Costa, G., Morel, J.-L.: Water relations, gas exchange and amino acid content in Cd-treated lettuce. - Plant Physiol. Biochem. 32: 561-570, 1994.
Delauney, A.J., Verma, D.P.S.: Proline biosynthesis and osmoregulation in plants. - Plant J. 4: 215-223, 1993. Hare, P.D., Cress, W.A., Van Staden, J.: Proline synthesis and
degradation: a model system for elucidating stress-related signal transduction. - J. exp. Bot. 50: 413-434, 1999. Hsu, Y.T., Kao, C.H.: Abscisic acid accumulation and cadmium
tolerance in rice seedlings. - Physiol. Plant. 124: 74-80, 2005.
Ireland, A.J., Lea, P.J.: The enzyme of glutamine, glutamate, asparagines, and aspartate metabolism. - In: Singh, B.K. (ed.): Plant Amino Acids. Pp. 49-109. Marcel Dekker, New York 1999.
Lin, C.C., Hsu, Y.T., Kao, C.H.: The effect of NaCl on proline accumulation in rice leaves. - Plant Growth Regul. 36: 275-285, 2002.
Marschner, H.: Mineral Nutrition of Higher Plants. - Academic Press, New York 1995.
Metha, S.K., Gaur, J.P.: Heavy-metal-induced proline
accumulation and its role in amelioating metal toxicity in Chlorella vulgaris. - New Phytol. 143: 253-259, 1999. Mishra, D., Kar, M.: Nickel in plant growth and metabolism. -
Bot. Rev. 40: 395-452, 1974.
Nakazawa, R., Kameda, Y., Ito, T., Ogita, Y., Michihata, R., Takenaga, H.: Selection and characterization of nickel-tolerant tobacco cells. - Biol. Plant. 48: 497-502, 2004. Rabe, E.: Stress physiology: the functional significance of the
accumulation of nitrogen-containing compounds. - J. hort. Sci. 65: 231-243, 1990.
Rai, V.K.: Role of amino acids in plant response to stresses. - Biol. Plant. 45: 481-487, 2002.
Schat, H., Sharma, S.S., Vooijs, R.: Heavy metal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris. - Physiol. Plant. 101: 477-482, 1997.
Tsai, Y.-C., Hsu, Y.T., Kao, C.H.: Proline accumulation induced by phosphinothricin in rice leaves. - Biol. Plant. 46: 213-320, 2003.
Vinterhalter, B., Vinterhalter, D.: Nickel hyperccumulation in shoot cultures of Alyssum markgrafii. - Biol. Plant. 49: 121-124, 2005.
Wang, C.Y., Cheng, S.H., Kao, C.H.: Senescence of rice leaves VII. Proline accumulation is senescing excised leaves. -Plant Physiol. 69: 1348-1349, 1982.
Weatherley, P.E.: Studies on the water relations of cotton plant. I. The field measurement of water deficit in the leaves. - New Phytol. 49: 81-97, 1950.
Yang, C.-W., Kao, C.H.: Ammonium in relation to proline accumulation in detached rice leaves. - Plant Growth Regul.
30: 139-144, 2000.
Yang, C.-W., Kao, C.H.: Importance of ornithine-δ-aminotransferase to proline accumulation caused by water stress in detached rice leaves. - Plant Growth Regul. 27: 189-192, 1999.
Yang, C.-W., Wang, J.W., Kao, C.H.: The relation between accumulation of abscisic acid and proline in detached rice leaves. - Biol. Plant. 43: 301-304, 2000.
Yoshiba, Y., Kiyosue, T., Nakashima, K., Yamaguchi-Shinozaki, K., Yamaguchi-Shinozaki, K.: Regulation of levels of proline as an osmolyte in plants under water stress. - Plant Cell Physiol. 38: 1095-1102, 1999.