以去氧核醣核酸之多型性來描繪川崎症; Genetic polymorphisms in Kawasaki disease patients
55
0
0
全文
(2) 中文摘要 儘管出生人口越來越少,但川崎症有逐年增加的趨勢,每年約 6000 名新病例被報告。川 崎症屬於全世界的疾病,尤好發於幼小孩子。其診斷依據是臨床表現,約有百分之二十有冠狀 動脈瘤併發症。 川崎症至目前其致病機轉仍不明,有人推測其致病原因為多因性,直到目前也沒有很好預 測指標可以預測川崎症的發生。因此我們將探討基因多形性(gene polymorphism)在川崎症所 扮演的角色。 Angiotensin- I converting enzyme(ACE) DD genotype 是造成一些心臟疾病如缺血性心臟 疾病,肥大性心肌病變,特異擴大心肌病變的危險因子。而在川崎症病人會製造多種發炎物 質,包括 interleukin-1(IL-1)、IL-4、IL-6、tumor necrosis factor-alpha。所以我們以聚合鏈反 應的方法研究川崎症和基因多形性的關係,探討的基因包括 ACE I/D、A-240T、G2350A、 interleukin-I ß (IL-1ß) promoter、IL-1 ß exon 5、IL-1 receptor antagonist、IL-4、IL-6、tumor necrosis factor-a 和 transforming growth factor-ß1。此研究目的除了欲了解 gene polymorphisms 在川崎症病人與正常人的分布是否不同,也想探討那一個 gene type 或 allele 影響川崎症病人合併動脈瘤的產生。 在此研究,統計方法是根據卡方檢定。結果,ACE gene I/D、A-240T、G2350A、IL-1 ß promoter、IL-1 ß exon 5、IL-1 receptor antagonist、IL-4、IL-6、tumor necrosis factor-a 和 transforming growth factor-ß1 對於川崎症病人合併動脈瘤的形成無影響,但 D/D genotype, G2350G genotype 及 G2350A 的 G allele 在川崎症病人的發生比例較正常控制組有明顯較低 的現象,IL-1 Ra I/II genotype 和 II allele 在川崎症病人的發生比例,較正常控制組有明顯較 高的現象,因此這些 genotype 和 allele 的存在和川崎症形成有關。. 關鍵詞:川崎症、基因多形性、冠狀動脈瘤. 2.
(3) 英文摘要 More than 6000 newly diagnosed cases are reported every year, and the number has been increasing year by year despite the decreasing birth rate. The disorder occurs worldwide, the illness occurs predominantly in young children. The diagnosis of Kawasaki disease is based on demonstration of characteristic clinical signs. Approximately 20% of untreated patients develop coronary artery aneurysms. The cause of the illness remains unknown. One hypothesis is that multifactors cause Kawasaki disease. Due to the lack of predictive markers for Kawasaki disease, we are going to detect the role of gene polymorphism in Kawasaki disease. Genotype DD of angiotensinogen I converting enzyme (ACE) polymorphism may be a risk factor for several diseases such as ischemic heart disease, hypertrophic cardiomyopathy and idiopathic dilated cardiomyopathy. In Kawasaki disease patients, the production of various inflammatory cytokines, including interleukin-1 (IL-1), IL-4, IL-6 and tumor necrosis factor-alpha is increased. We examined the gene polymorphism of Kawasaki disease patients using PCR. The polymorphisms of the gene including ACE I /D, A-240T, G2350A, interleukin-I ß (IL-1 ß) promoter, IL -1 ß exon 5, IL-1 receptor antagonist, IL -4, IL-6, tumor necrosis factoralpha and TGF-beta 1 were examined. In this study, we investigated the gene polymorphism among children with Kawasaki disease and also for an appropriately matched control group. We also investigated the relationship of frequencies of the alleles and genotype of these genes and coronary aneurysm formation. In this study, the data were analyzed using chisquare test. We have found that the DD genotype; G2350G of the ACE gene polymorphism and the G allele at G2350A are present at a significant lower frequency among patients suffering Kawasaki disease. Kawasaki patients are significantly more likely to posses an I/II genotype and II allele of the IL-1 Ra. There appears to be no significant association between. 3.
(4) ACE gene I/D, A-240T , G2350A, IL -1 ß promoter, IL -1 ß exon 5 , IL-1 receptor antagonist , IL-4 , IL-6 , tumor necrosis factor-alpha 和 transforming growth factor-ß1 gene polymorphisms and the potential for the formation of a coronary artery aneurysm formation for Kawasaki disease sufferers. In conclusion, the genotype and allele of ACE gene polymorphisms and IL -1 Ra gene polymorphisms are associated with Kawasaki disease.. Key words : kawasaki disease, genetic polymorphism, coronary artery aneurysm. 4.
(5) 誌. 謝. 本人自母校醫學系畢業後,就一直從事小兒科的醫療工作,至今已拾玖年; 隨著醫學的進步,深感基礎醫學學識欠缺不足,故興起再進修的念頭。於民國九 十年,我終於一償宿願,進入母校醫學研究所就讀。 在這兩年裡,過著臨床醫療與研究所兩頭忙的日子,實在感謝小兒腸胃團隊 的體恤,讓我有更多的時間專心於課業上。而這二年的研讀中,最要感謝蔡輔仁 教授的教導,使本論文得以順利完成;另外要謝謝陳汶吉博士不厭其煩的講解, 使我收穫良多。而研究所的每一位師長,因為您們的傳道、授業、解惑,使這二 年得以過得充實,最後要多謝家人的支持,我才得以無後顧之憂完成學業。. 5.
(6) 目. 錄. 中文摘要. ---------------------------------------------------------------------. 2. 英文摘要. ---------------------------------------------------------------------. 3. 誌謝辭. ---------------------------------------------------------------------. 5. 目錄. ---------------------------------------------------------------------. 6. 表目錄. ---------------------------------------------------------------------. 7. 圖目錄. ---------------------------------------------------------------------- 10. 主文. ---------------------------------------------------------------------- 13. 前言. ---------------------------------------------------------------------- 13. 材料方法. ---------------------------------------------------------------------- 18. 結果. ---------------------------------------------------------------------- 22. 討論. ---------------------------------------------------------------------- 24. 參考文獻. ---------------------------------------------------------------------- 32. 圖表. ---------------------------------------------------------------------- 40. 簡單自述. ---------------------------------------------------------------------- 55. 著作權聲明 ---------------------------------------------------------------------- 56. 6.
(7) 表目錄 Table 1 Main characteristics of IL-1ß promoter, IL -1 ß exon 5, IL-1 Ra, IL-4 intron 3, IL-4 promoter and IL -6 promoter gene polymorphisms and techniques. Table 2. Distribution of ACE I/D genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 3. Distribution of ACE (A-240T) genotypes and allelic frequencies for patients suffering Kawasaki disease and also for healthy control subjects. Table 4. Distribution of ACE (G2350A) genotypes and allelic frequencies for patients suffering Kawasaki disease and also healthy control subjects. Table 5. Phenotype frequencies of the polymorphism in the ACE I/D gene in patients with and without coronary-artery aneurysm. Table 6. Phenotype frequencies of the polymorphism in the ACE (A-240T) gene in patients with and without coronary-artery aneurysm. Table 7. Gene frequencies of the polymorphism in the ACE (A-240T) gene in patients with and without coronary-artery aneurysm. Table 8. Phenotype frequencies of the polymorphism in the ACE (G2350A) gene in patients with and without coronary-artery aneurysm. 7.
(8) Table 9. Distribution of I L-1 ß promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 10. Phenotype and alleleic frequencies of the polymorphism in the IL -1 ß promoter gene in patients with and without coronary-artery aneurysm. Table 11. Distribution of I L-1 ß exon 5 genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 12. Phenotype and alleleic frequencies of the polymorphism in the IL-1 ß exon 5 gene in patients with and without coronary-artery aneurysm. Table 13. Distribution of I L-1 ß Ra genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 14. Phenotype and alleleic frequencies of the polymorphism in the IL -1 Ra gene in patients with and without coronary-artery aneurysm. Table 15. Distribution of I L-4 promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 16. Phenotype and alleleic frequencies of the polymorphism in the IL -4 promoter gene in patients with and without coronary-artery aneurysm. 8.
(9) Table 17. Distribution of I L-4 intron 3 genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 18. Phenotype and alleleic frequencies of the polymorphism in the IL -4 intron 3 gene in patients with and without coronary-artery aneurysm. Table 19. Distribution of IL -6 promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 20. Phenotype and alleleic frequencies of the polymorphism in the IL -6 promoter gene in patients with and without coronary-artery aneurysm. Table 21. Distribution of TNF-a promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 22. Phenotype and alleleic frequencies of the polymorphism in the TNF-a gene in patients with and without coronary-artery aneurysm. Table 23. Distribution of TGF-ß 1 genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Table 24. Phenotype and alleleic frequencies of the polymorphism in the TGF-ß 1 gene in patients with and without coronary-artery aneurysm. 9.
(10) 圖目錄. Figure 1. Angiotensin-1-converting enzyme gene polymorphism shown on 3% agarose electrophoresis. The polymorphic region was amplified by PCR, resulting in a fragment at lane II (495 bp),a heterozygote at lane ID (495bp and 208bp)and a homozygote at lane DD (208 bp).M=Marker, 100-bp-ladder.. Figure 2. PCR based restriction analysis of the angiotensin-1-converting enzyme gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a heterzygote at lane A/T,a cuttable length at lane T/T, (114 bp and 23 bp), and a uncuttable fragment at lane A/A (137 bp). The 23 bp band was obscure during the electrophoresis process.. Figure 3. PCR based restriction analysis of the angiotensin-1-converting enzyme gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a uncuttable fragment at lane AA (122bp), and a heterozygote at lane AG,and a cuttable fragment at lane GG (103bp and 19 bp).The 19 bp band was obscure during the electrophoresis process.. Figure 4. PCR based restriction analysis of the interleukin-1 ß promoter gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a uncuttable fragment at lane TT (304bp), and a heterozygote at lane CT,and a cuttable fragment at lane CC (190bp and 114bp).The 114bp band was obscure during the electrophoresis process.. 10.
(11) Figure 5. PCR based restriction analysis of the interleukin-1 ß exon 5 gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a uncuttable fragment at lane E2 (249bp), and a heterozygote at lane E1E2, and a cuttable fragment at lane E1E1 (135bp and 114bp). The 114bp band was obscure during the electrophoresis process.. Figure 6. PCR based restriction analysis of the interleukin-1 Ra gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in an 86 bp copies. The variable number of tandem repeats was obscure during the electrophoresis process.. Figure 7. PCR based restriction analysis of the interleukin-4 promoter gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a uncuttable fragment at lane TT (252bp), and a heterozygote at lane CT,and a cuttable fragment at lane CC (192bp and 60bp).The 60bp band was obscure during the electrophoresis process.. Figure 8. PCR based restriction analysis of the interleukin-4 intron 3 gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a 70 bp copies. The variable number of tandem repeats was obscure during the electrophoresis process.. Figure 9. PCR based restriction analysis of the interleukin-6 promoter gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR,. 11.
(12) resulting in a uncuttable fragment at lane GG (299bp), and a heterozygote at lane CG,and a cuttable fragment at lane CC (238bp and 61bp).The 61bp band was obscure during the electrophoresis process.. Figure 10. PCR based restriction analysis of the tumor necrosis factor alpha gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a uncuttable fragment at lane AA (117bp), and a heterozygote at lane AG,and a cuttable fragment at lane GG (97bp and 20bp).The 20bp band was obscure during the electrophoresis process.. Figure 11. PCR based restriction analysis of the transforming growth factor ß1 gene polymorphism shown on 3% agarose electrophoresis. The polymorphism region was amplified by PCR, resulting in a uncuttable fragment at lane TT (120bp), and a heterozygote at lane CT,and a cuttable fragment at lane CC (74bp and 46bp).The 46bp band was obscure during the electrophoresis process.. 12.
(13) 主文 :. Genetic polymorphisms in Taiwanese children with Kawasaki disease. Introduction. Kawasaki disease (KD) is an acute febrile vasculitis of childhood first described by Dr. Tomisaku Kawasaki in Japan in 1967(1). The disorder occurs worldwide, with Asians at highest risk. Kawasaki disease has replaced acute rheumatic fever as the leading cause of acquired heart disease in children. The cause of the illness remains unknown. It is estimated that at least 3000 cases are diagnosed annually in the United States. The incidence of Kawasaki disease in Asian children is substantially higher than in other racial groups, but the illness occurs worldwide in all ethnic groups. In Japan, more than 150000 cases have been reported since the 1960s.the illness occurs predominantly in young children; 80% of patients are younger than 5 yr (2), and only occasionally are teenagers and adults affected. Approximately 15 to 25% of untreated patients will develop damage to the coronary arteries as a result of intense inflammation in the vessel wall (3, 4, 5).. A number of immunoregulatory changes are observed in children with KD that may contribute to the pathogenesis of the disease. Serum levels of several immune mediators are. 13.
(14) increased, including IL -1, IL-2, IL-4, IL-6, IL-8 and TNF-a (6, 7, 8, 9, 10). In vitro studies using vascular endothelial cells had demonstrated that IL-1 and TNF-a induce the expression of surface antigens that render the cells susceptible to lysis by IgG or IgM antibodies in the sera of children with acute KD (11, 12). TNF-alpha levels are elevated in the majority of children during the acute phase of the disease (13, 14, 15).. The stimulus in KD for the increased serum levels of IL -1, IL-4, IL-6, and TNF-a remains unknown. Genetic polymorphisms influence the magnitude of the cytokine response after an inflammatory stimulus. To determine whether gene polymorphisms might play a role in KD, we analyzed the gene including ACE I/D, A-240T, G2350A, interleukin-I ß (IL-1 ß), IL-1 receptor antagonist, IL -4, IL-6, tumor necrosis factor-alpha and TGF-beta1 in Taiwanese children with KD and control populations.. Angiotensin converting enzyme (ACE) is a zinc metalloprotease that catalyzes the hydrolysis of carboxyterminal dipeptides from oligopeptide substrates (16). ACE is produced mainly by vascular endothelium (17), but a high concentration of ACE can also be detected in the kidney, the brain, the testis, and epithelial surfaces (16). ACE converts angiotensin I to angiotensin II, a potent vasoconstrictor and stimulator of aldosterone secretion by the adrenal gland.. Circulating ACE levels show extensive interindividual variability and are highly genetically determined (18, 19, 20, 21). An insertion/deletion (I/D) dimorphism, due to the presence or absence of a 287 base pair alμ-type sequence in the 16th intron of the ACE gene, has been shown to cosegregate with serum and tissue ACE activities, and major locus inheritance explains best the findings that the D allele is associated with elevated ACE. 14.
(15) Levels (19,21). These findings have been recurrently confirmed by several investigators in populations of various ethnic origins (22, 23, 24, 25, 26, 27). Thus, the ACE gene is viewed as a quantitative trait locus that modulates circulating ACE levels, and the ACE I/D dimorphism is a marker that is in linkage disequilibrium with functional variants located in the ACE gene (28).. The D allele is associated with myocardial infarction, increase plaque instability, stent restenosis, LVH, ischemic or idiopathic cardiomyopathy, coronary artery disease, and other cardiovascular diseases, including diabetes mellitus and diabetic nephropathy.. The ACE A-240T polymorphism is located in the 5’ section of the gene, accounting for 6 % of the total variance in ACE. The ACE G2350A polymorphism is in intron 17, accounting for 19 % of the variance.. Cytokines with polymorphic gene sequences are potential markers of disease severity since their gene products are involved in KD pathogenesis. Differences in severity between individuals could be related to different levels of cytokine production or functional differences resulting from polymorphisms in their genes.. IL-1 is one of the most potent proinflammatory agents, and it has a central role in joint inflammation and destruction. The genes for IL -1 are located on chromosome 2, in close linkage with the IL-1 receptor antagonist (IL -1 Ra) gene (29). Different polymorphisms have been described in the IL -1 ß gene, and at least 2 of them could influence the protein production: one is located within the promoter region (30), the other in exon 5 (31).. 15.
(16) The IL-1 receptor antagonist (IL -1 Ra) is its natural competitive inhibitor, acting by occupancy of IL-1 cell surface receptor without triggering signal transduction (32). IL -1 Ra plays a role as an important regulator of inflammation and is currently being evaluated in clinical trials. Five alleles of the IL -1Ra gene have been reported, corresponding to 2, 3, 4, 5, and 6 copies of and an 86-basepair sequence repeat located in intron 2 (33). Because 3 potential protein-binding sites are located in this 86-bp sequence, the number of repeats may influence gene transcription and protein production. Moreover, dimers have been shown to be associated with a higher IL -1 Ra secretion by mononuclear cells in vitro, which is activated by granulocyte-macrophage colony-stimulating factor (34).. IL-4 is a cytokine with antiinflammatory properties, its production is increased in KD patients. The gene for IL -4 has been mapped to the q arm (q23-31) of chromosome 5 (35). A functional polymorphism, representing a C-to-T substitution at position -590, has been recently described in the promoter region of IL -4 (36). Another polymorphism has been located in the third intron, and is composed of a variable number of tandem repeats of a 70bp sequence (37).. IL-6 is a multifunctional cytokine. A recently described G/C polymorphism at position-174 in the human IL-6 promoter (38) has been reported to be associated with an altered IL-6 promoter activity and with different plasma levels of IL -6 in healthy men (39). The C allele─and more so the CC genotype—will result in a lower IL-6 expression after a given inflammatory stimulus compared with the GG genotype.. Tumor necrosis factor alpha (TNFa) is a central mediator of the immunological response and the location of the gene within the major histocompatibility complex has. 16.
(17) prompted much speculation about the role of TNF a alleles in inflammatory diseases. A G to A transition polymorphism at position -308 of the TNFa promoter/enhancer region has been described. The -308 A allele was shown to be strongly associated with human leukocyte antigen (HLA)-DR3, known to be related to a TNFa high producer phenotype.. Transforming growth factor ß1 (TGF-ß1) is a multifunctional cytokine. It regulates cell growth, differentiation and matrix production. TGF-ß1 is to maintain a normal immune function. TGF-ß1gene is located on chromosome 19q13, contains seven exons that give rise to a precursor protein of 390 amino acids, which is proteolytically processed to generate the mature protein of 112 amino acids. Seven TGF-ß1 polymorphisms have been reported. Three were localized in the 5-flanking region of the TGF-ß1gene (at positions-988, -800,and 509), three were in the coding region (codons 10, 25,and 263), and an insertion was in the 5untranslated region at position +72.. The etiology of Kawasaki disease is unknown. Several reports have indicated that this illness is linked to genes (40, 41). Gene polymorphisms have important implications in human genetic studies. Due to the lack of predictive markers in Kawasaki patients, we hope to investigate the relationship between Kawasaki disease-suffering patients and gene polymorphisms.. 17.
(18) MATERIALS AND METHODS Subjects We collected Kawasaki disease patients from the department of paediatrics at the China Medical college Hospital from July 1997 to December 2002. The 107 patients suffering Kawasaki disease included in our study group comprised 65 boys and 42 girls, aged from 0.2 to 3.2 years of age, with a mean age of 1.7 years, all of whom fulfilled the appropriate diagnostic criteria for Kawasaki disease. All patients received treatment with intravenous gamma globulin (2gm/Kg/day for one day plus oral aspirin at 80mg/Kg/day). Participant blood samples were obtained on the day of admission. Twodimensional echocardiography was used to detect the presence of any coronary-artery lesions. Coronary arteries with diameters of 4mm or greater were classified as abnormal. A total of 29 patients developed coronary-artery aneurysm, whilst 78 patients revealed no evidence of coronary-artery aneurysm. We also studied 107 healthy children (80 boys and 27 girls aged 0.4 to 3.0 years, with a mean age of 1.8 years) acting as the control group. Control samples were tested in parallel with patient samples. In this study, we investigated the gene including ACE I/D, A-240T, G350A, interleukin-I ß (IL-1 ß) promoter, IL -1ß exon 5, IL-1 receptor antagonist, IL -4, IL-6, tumor necrosis factor-alpha and TGF-beta 1 in all subjects. This study was carried out with approval from the Human study Committee of the China Medical College Hospital. Information was obtained from family of patients. The genomic DNA was prepared from peripheral blood. Purified genomic DNA was amplified by PCR amplication. PCR products were analyzed by electrophoresis on agarous gel and each allele was recognized according to its size. Determination of the genotype of the I/D, A-240T and G2350A polymorphisms of the. 18.
(19) ACE gene Polymerase chain reactions (PCRs) appropriate for demonstrating the I/D polymorphism of the ACE gene were carried out in a total volume of 50ul, containing genomic DNA, 2-6pmol of each primer, 1 X Taq polymerase buffer (1.5mM MgCl2), and 0.25 units of Ampli Taq DNA polymerase (Perkin Elmer, Foster City, Calif., USA). The primers for the insertion/deletion (I/D) polymorphism of ACE gene were forward 5-TGGAGACCACTCCCATCCTTTCT-3 and backward 5-CAGGTCTTCATATTTCCGATGTGG-3. The primers were designed according to the sequences of human ACE mRNA, completed cds (JO4144) and H. sapiens ACE gene (intron 16) (X62855), which were available from the web site (http://www.ncbi.nih.gov/). The PCR amplifications were performed using the Gene Amp PCR System 2400 programmable thermal cycler (Perkin Elmer). The cycling conditions for I/D were set as follows: one cycle at 94ºC for three minutes, 30 cycles at 94ºC for 30 seconds, 60º C for 30 seconds, and 72ºC for 60 seconds, and one final cycle of extension at 72ºC for ten minutes. Through electrophoresis analysis on 3% agarose gel, the I/D polymorphism in intron 16 of the ACE gene was categorized as the PCR fragment of the insertion type (495 bp in length) and the deletion type (208 bp in length; Figure 1). The A-240T and G2350A polymorphisms were typed by the restriction fragment length polymorphism method. The primer of A-240T was F:TCGGGCTGGGAAGAAGATCGAG, R:GAGAAAGGGCCTCCTCTCTCT The restriction endonuclease was Xbal. The fragment size was 137/114 (Figure 2). The primer for G2350A was F: CTGACGAATGTGATGGCCGC, R: TTGATGAGTTCCACGTATTTCG, and the restriction endonuclease was BstUI. The fragment size was 122/103. (Figure 3) Determination of the genotype of the polymorphisms of the IL-1 gene, IL-4 gene, IL-6 gene PCR was used to identify the genotypic pattern of all the different cytokine genes. Details. 19.
(20) for each polymorphism analysis are given in Table (1) and Figure (4, 5, 6, 7, 8, 9) Determination of the genotype of the polymorphisms of the TNF-a gene TNF-a gene polymorphism loci involves a G to A transition at position -308 in the 5-flanking promoter region. The sequences of the primers were as following: 5’-AGGCAATAGGTTTTGAGGGCCAT-3’, and 5’-ACACTCCCCATCCTCCCGGCT-3’ 50ng of genomic DNA was mixed with 20 pmol of each PCR primer in a total volume of 25µl containing 10mM Tris-HCL, PH 8.3, 50mM KCL, 2.0mM MgCL2, 0.2 mM each deoxyribonucleotide triphosphate, and 1 unit of Ampli taq DNA polymerase. PCR was programmed as follows: 35 cycles of 95 ℃ and 60℃ for 15 sec and 30 sec, respectively. (Figure 10) Determination of the genotype of the polymorphisms of the TGF-ß1 gene Genotyping of C-509T polymorphisms of the TGF-ß1 gene Genomic DNA was prepared from peripheral blood leukocytes using a genomic DNA isolation Kit. PCRs were carried out to a total volume of 50ul containing genomic DNA, 2-6 pmol of each primer, 1X Taq polymerase buffer (1.5 mM MgCl2), and 0.25 unit of Pro-Taq DNA polymerase. The C-509Tpolymorphisms of the TGF-ß1 gene were typed using the restriction fragment length polymorphism (RFLP) method. Primer sequences forward : GGAGAGCAATTCTTACAGGTG backward :TAGGAGAAGGAGGGTCTGTC .. The PCR amplifications were performed in the GeneAmp PCR System 2400. programmable thermal cycler. The cycling conditions were set as follows: one cycle at 94℃ for 3 min, 30 cycles at 94℃for 30 s, 60℃for 30 s, and 72℃ for 60 s, and one final cycle of extension at 72℃for 10 min. PCR fragments through DdeI digestion reaction and analyzed. 20.
(21) by 3% agarose gel electrophoresis. A non-digested fragment was a single band of 120 bp (TT), the C-509 genotype was digested as 46 and 74 bp in length (CC), and heterozygotes (CT) showed three fragments of 120, 46 and 74 bp. (Figure 11) STATISTICAL METHODS The Chi-square test was used to compare the genotype and the allelic frequency distribution of the ACE gene for patients with Kawasaki disease and also for control subjects. A Chi-square test was also used to compare groups revealing the presence of a coronaryartery aneurysm as compared to those who did not. A value of p < 0.05 was considered to represent significant difference between tested populations.. 21.
(22) RESULTS The distribution of the I/D, A-240T and G2350A polymorphism of the ACE gene is revealed in Tables 2, 3 and 4. The analysis of ACE D/D genotype distribution in the control group and KD group revealed16.8% and 5.6% respectively. The analysis of ACE 2350 G/G genotype distribution in the control group and KD group revealed 16% and 5.6% respectively. The analysis of ACE D/D genotype and ACE 2350GG genotype distribution revealed a statistically-significant difference between KD patients and controls. As shown in Tables 5, 6, 7 and 8, no significant difference as regards I/D, A-240T and G2350A polymorphism of the ACE gene between patients with and without coronary-artery aneurysm was observed. The distribution of the IL -1 ß promoter, IL -1 ß exon 5 and IL -1 Ra polymorphism gene is revealed in Tables 9, 10, 11, 12, 13, 14. The analysis of genotype and allelic frequency distribution of IL -1 ß promoter and IL -1 ß exon 5 revealed no significant difference between Kawasaki-disease patients and controls. The I/II genotype and II allele of the IL -1 Ra gene were found with a significantly higher frequency in KD patients compared with controls. The distribution of genotype and alleic frequency of IL -1 gene polymorphisms between patients with and without coronary-artery aneurysm was no significant difference. The distribution of the polymorphisms of IL -4 promoter and IL -4 intron 3 gene is revealed in Tables15, 17. The analysis of genotype and alleic frequency distribution revealed no significant difference between Kawasaki-disease patients and controls. As shown in Tables 16, 18, no significant difference as regards IL -4 gene polymorphisms between patients with and without coronary-artery aneurysm was observed. The distribution of the IL -6 promoter gene polymorphisn is revealed in Tables 19. The analysis of genotype and alleic frequency distribution revealed no significant difference between Kawasaki-disease patients and controls. As shown in Tables 20, no significant. 22.
(23) difference as regards IL -6 gene polymorphism between patients with and without coronaryartery aneurysm was observed. The distribution of the TNF-a gene polymorphism is revealed in Tables 21. The distribution in control group revealed 6.8% A-308 allele homozygote, 29.1 heterozygote and 64.1 % G -308 allele homozygote. The genotype distribution in KD group reveled 15.9 % A308 allele homozygote, 22.4 % heterzygote and 61.7 % G-308 allele. The allelic frequencies in the control group were 21.4% A and 78.6% G. The analysis of genotype and alleic frequency distribution revealed no significant difference between Kawasaki-disease patients and controls. As shown in Tables 22, no significant difference as regards TNF-a gene polymorphism between patients with and without coronary-artery aneurysm was observed. The distribution of the TGF-ß1 gene polymorphism is shown in Tables 23. The analysis of genotype and alleic frequency distribution revealed no significant difference between Kawasaki-disease patients and controls. As shown in Tables 24, no significant difference as regards TGF-ß1 gene polymorphism between patients with and without coronary-artery aneurysm was observed.. 23.
(24) DISCUSSION Tomisaki first described Kawasaki disease in 1967 (42). The exact cause of Kawasaki disease would appear to still be largely unknown. Serious complications to the coronary artery may occur as a result of this disease (43). Kawasaki disease is also one of the leading causes of acquired heart disease(s) in the pediatric field. In a number of previous reports, the genotype DD of the ACE gene may have constituted a risk factor for heart disease including coronary-artery disease, thus, here, we have attempted to establish the relationship between Kawasaki disease-suffering patients and ACE gene polymorphisms. The human ACE gene has been cloned and localized to chromosome 17q23 (44). A 287 bp insertion/deletion polymorphism in intron 16 has been identified for use as a genetic marker (45). The A-240T polymorphism is on the promoter region of the ACE gene. The G2350A polymorphism is located in intron 17. Circulating levels of ACE are under substantial genetic control, there being ample evidence to suggest that the I/D polymorphism is in strong linkage disequilibrium with a major gene effect at the ACE gene locus, this locus being suggested to control up to 44% of the variability in ACE levels (46). Philippe. et al. (1998) analyzed the distribution of mean serum ACE activities according to ACE I/D genotypes for different age groups of Emirati subjects, these authors finding that the correlation between the ACE I/D dimorphism and circulating serum ACE activities was able to be quite easily documented. Mean ACE levels were lowest amongst II homozygotes, intermediate for ID heterozygotes, and highest amongst DD homozygotes (47). Further studies have shown that the T allele of the A-240T polymorphism and the G allele of the G2350A polymorphism were associated with an increase in circulating ACE concentration (48, 49). ACE is a zinc metalloprotease that cleaves angiotensin I into the angiotensin II molecule which is a promoter molecule for vasoconstriction. Further, ACE is produced mainly by vascular endothelial cells (50), it being frequently reported that the vascular lesions that. 24.
(25) typically arise amongst sufferers of Kawasaki disease are associated with endothelial-cell damage of, particularly, small and medium blood vessels the ACE level depression arising subsequent to blood-vessel endothelial-cell injury (51). Falcini et al. (1996) reported ACE values were significantly lower amongst active Kawasaki-disease sufferers than was the case for healthy children, it being suggested that the decreased ACE levels under such circumstances may be linked to a diffuse vascular inflammatory process (52). Since we didn’t attempt to measure the ACE activity level amongst our test-group individuals, the relationship between the ACE gene polymorphism and ACE levels was not able to be clearly elucidated. For this case-controlled study, the ACE gene polymorphisms noted were hypothesized to be associated with the coronary artery aneurysms associated with Kawasaki disease. Our results suggest no evidence of an association between ACE gene polymorphisms and the formation of coronary-artery aneurysms. This result differs from that contained in the report of Takeuchi in which Kawasaki-disease patients also suffering coronary-artery aneurysm reflected the presence of the genotype II ACE polymorphism more frequently than was the case for non-aneurysm sufferers amongst a Japanese subpopulation (53). In our study, the distribution of the DD genotype, G2350G of the ACE gene polymorphism amongst Kawasaki-disease patients appeared to differ to that for the control group, there existing significant difference between both groups (Tables 2 & 4). In this study, the increased risk of Kawasaki disease associated with the A allele of the G2350A polymorphism is also observed. According to the interpretation of Frossard (1998) and Zhu (2001) data, it would appear that expression of the DD genotype of the ACE gene and the G allele of the G2350A polymorphism were associated with an increase in ACE activity (47,49).These fragments is located on an intron and thus cannot affect the expression of mRNA directly. It is hypothesized that the insertion/deletion and G2350A fragments are in. 25.
(26) linkage disequilibrium with a still unknown DNA fragment that acts as a silencer fragment. Our study group of Kawasaki-disease patients did reveal a lower level of the DD genotype and the G allele of the G2350A polymorphism than was the case for the control group, for which case, the ACE concentration should be lower than is the case for the control case. The ACE inhibition reduced free radical expression, however, its role in modulating the inflammatory response has not been clearly defined. Alternatively, renin-angiotensin systems may influence KD pathogenesis via more global effects in other tissues. For example, vascular wall might modulate the response to systemic inflammation. All such hypotheses are worthy of further study. Contrasting our patients suffering coronary aneurysm with those who did not, we do not appear to demonstrate any difference in distribution of the ACE-gene polymorphism between these two groups; such a result suggesting that ACE activity does not relate to severity of KD. For all 107 Kawasaki-disease patients, the distribution of the I/D ACE genotype was II:0.28, I/D:0.66 and DD:0.05, such results being comparable with the results of Takeuchi (1997), who reported for 36 Japanese Kawasaki-disease patients, that the distribution of the I/D ACE genotype was II:0.42, I/D:0.50 and DD:0.08 (53). The allele frequencies were 0.61 for the I allele and 0.39 for the D allele amongst our Kawasaki-disease patients (Table 2), similar to corresponding levels of 0.66 for the I allele and 0.34 for the D allele amongst the Japanese sub-population as reported by Takeuchi in 1997 (53). If the Kawasaki-disease patient in Asia reflects a similar ACE gene polymorphism, then further, more-expansive studies would appear to be needed. Studies of cytokines have been reported that proinflammatory cytokines such as IL -1, IL-4, IL-6 and TNF-a are elevated in the sera of KD patients (12, 13). These results suggest that cytokines play an important role in the onset of this disease. The inflammatory reaction produced by a number of factors, including the type of stimulus, the dose of stimulus, and. 26.
(27) genetic characteristics of the host. The stimulus for the inflammatory reaction in children with KD remains unknown. Some cytokine polymorphisms might be related to KD. Interleukin 1 belongs to a cytokine family modulating cellular proliferation and has the capacity to induce other cytokines. It is a primary mediator of the inflammatory response and has been shown to induce prostaglandin synthesis (54). The IL -1 genes are associated with several immunoinflammatory diseases (55). Interleukin I exists in 2 forms, IL-1α and IL-1β, which are encoded by distinct genes but share the same receptors and biological properties( 56). The loci for IL -1α and IL-1β are located on the proximal region of the long arm of chromosome 2 ( 57). The IL -1β polymorphism has been correlated with IL-1β expression( 58). Different polymorphisms have been described in the IL -1ß gene, and at least 2 of them could influence protein production : one located in the promoter region at position511(IL-1β-511) and the other in exon 5(59, 60). The allele E2 of IL-1 ß exon 5 has been described to be associated with an IL -1ß high secretor gene. The IL-1Ra is structurally related to IL -1 a and IL-1βand competes with these molecules for occupation of IL -1 cell surface receptors. The presence of the IL-1Ra allele II was associated with enhanced IL -1βproduction in vitro(61). The type II IL-1Ra allele has been previously found in association with a variety of autoimmune diseases: alopecia areata, lichen sclerosus, systemic lupus erythematosus, ulceractive colitis, and late-onset psoriasis. These genes code several proteins that may be key components in the pathogenesis of KD. In this study, we observed that the IL-1 Ra genotype I/II and II allele are associated with higher susceptibility to KD. These observations suggest that genes that contribute to regulating the level of IL-1 production may be useful in predicting the occurrence of KD. The IL-1 Ra genotype and II allele may be involved in the formation of KD through a complex pathway. Such as change the pathway of signal transduction between cells and then change the function of transcription. Endogenous production of IL -1 Ra is an important anti27.
(28) inflammatory mechanism in human disease. IL -1 Ra binds to IL -1 receptor with an avidity equal to that of IL-1, so fails to stimulate the cells. In our KD patients, an imbalance exists in this system, because the relative levels of production of IL -1 Ra are not adequate to effectively block the proinflammatory effects of IL -1.The IL-1ß promoter and IL -1ß exon 5 gene polymorphisms are not useful in predicting the susceptibility to KD. Previous report (55) showed that IL-1 genes may have a role in the severity of the disease rather than in susceptibility to the disease itself. But in this study, we observe no association of IL -1 gene polymorphisms between patients suffering coronary aneurysm with those who did not. IL-4 is a helper T cell type 2 cytokine involved in the promotion of humoral immunity. The IL-4 -590T allele has been shown to be associated with an enhanced IL -4 activity as measured by IgE production in Jurkat cells. This suggests that a C- to- T exchange at position -590 enhances IL-4 production or activity by T cells. In the study by Hunt et al., the frequency of the T allele of the C/T polymorphism of the IL -4 gene was 14 % in the control subjects. In our study, we found that the frequency of the T allele was 77.7% in controls to be different to Hunt et al. The difference in the result may reflect several factors. There may be genuine geographical differences in the data sets, however, this is unlikely as all controls. IL-4 intron 3 RP1 and RP2 alleles are located in a polymorphic region in the third intron and are characterized by 2 and 3 tandem repeats, respectively, of a 70-bp sequence (37). More recently, a third allele has been reported, including 4 tandem repeats, but we do not observe this rare allele in our population. Many of the polymorphisms have been related to different levels of cytokine production, but the functional incidence of the intron 3 polymorphism of the IL -4 gene is not known. It can be hypothesized that distinct numbers of variable number of tandem repeats (VNTR) copies may affect the transcriptional activity of the IL-4 gene. The human gene for IL-4 has been mapped to the q arm of chromosome 5, in a cluster of cytokine genes which code proteins that are important in the control of different. 28.
(29) aspects of immune response (62). Our data shows that the C-T promoter and intron 3 RP1/RP2 polymorphism of the IL-4 gene do not confer protection against the development of KD and coronary aneurysm formation. IL-6 is a multifunctional cytokine expressed in many tissues, including adipose tissue, skeletal muscle and hypothalamus. We examined the IL -6 gene promoter, G/C polymorphism at position -174 has been reported to be associated with an altered IL -6 promoter activity and with different plasma levels of IL -6 in healthy men (39). The recently described C-174G promoter polymorphism of the IL -6 gene has been found to influence plasma IL -6 levels in patients with systemic-onset juvenile chronic arthritis (63) and in patients with primary sjören’s syndrome. Although KD patients demonstrate a drastic increase in serum IL -6 during acute phase, there were no significant differences in the nucleotide sequence between the KD patients and normal control group. The stimulus in KD for the increased serum levels of TNF-a remains unknown. The gene coding for TNF-a lies within the MHC on chromosome 6 (64). A number of genetic polymorphisms upstream from the coding sequence for TNF-a have been described. The presence of the A allele at the –308 site is associated with elevated TNF-a production in response to endotoxin in whole blood cell cultures (65). An increased frequency of the TNFa-308 A allele has been associated with poor outcome after certain infections. We tested the hypothesis that children with KD may have a higher frequency of the A allele at the TNF-a308 sites, which is associated with elevated serum levels of TNF-a after an inflammatory stimulus. The results of our study demonstrate that the TNF-a-308 gene A allele is not associated with KD and aneurysm formation. Transforming growth factor-beta (TGF-β) is a multifunctional cytokine involved in many cellular processes, such as cell proliferation, embryonic development, tumorigenesis, wound healing, fibrosis, and immune and inflammatory cell responses (66,67). Its. 29.
(30) immunoregulatory effects include the inhibition of immunocompetent cells and the regulation of cytokine production (66). KD is an acute febrile illness and one of the most important forms of systemic vasculitis occurring in early children. Vasculitis associated with KD is characterized by initiation of a pro-inflammatory cytokine cascade that results in dramatic immune activation. Matsubara et al. reported the decrease in the concentrations of TGF-β1 in the sera of patients with KD (68). The decrease in TGF-β1 may result in an increase in TNF-α, because TGF-β1 can suppress TNF-α production from macrophage/monocytes. The mechanisms leading to the decreased TGF-β1 levels remain unknown. We postulated the possible role of TGF-β1 to be involved in the pathogenesis of KD. In our case-controlled study, the TGF-β1 gene C-509T polymorphism, the frequencies of the T allele and TT genotype in the control population were 54 and 31 %, respectively- somewhat different from those in the Caucasian population (24 and 5 %, respectively) (69). The distribution of the C509T polymorphism was studied in the KD and control groups, and studied in the patients suffering coronary aneurysm and those who did not. The results showed no evidence of any association of TGF-β1 gene C-509T with KD. In conclusion, we have found that the DD genotype; G2350G of the ACE gene polymorphism and the G allele at G2350A are present at a significantly-lower frequency amongst patients suffering Kawasaki disease than is the case for controls .The I/II genotype and II allele of IL-1 Ra are associated with KD. These data illustrate that a different gene regulation of KD pathogenesis. It is highly unlikely that polymorphism alone represents the susceptibility gene for the development of disease. In addition, there appears to be no significant association between ACE gene, IL -1 gene, IL-4 gene, IL-6 gene, TNF-a gene and TGF-ß1 polymorphisms and the potential for the formation of a coronary-artery aneurysm. 30.
(31) formation for Kawasaki-disease sufferers.. 31.
(32) References 1.Kawasaki T.: Acute febrile mucocutaneous syndrome with lymphoid involvement with specific desquamation of the fingers and toes in children.Jpn J Allerg 1967;16:178-222 2.Yanagawa H, Kawasaki T, and Shigematsu I. Nationwide survey on Kawasaki disease in Japan. Pediatrics 1987; 80:58-62 3.Kawasaki T, Kosaki F, Okawa S, Shigematsu I, Yanagawa H. A new infantile acute febrile mucocutaneous lymph node syndrome prevailing in Japan. Pediatrics 1974; 54:271-276 4. Melish ME, Hicks RM, Larson EJ. Mucocutaneous lymph node syndrome in the Unites states. Am J Dis Child 1976;130: 599-607 5.Kato H, Koike S, Yamamoto M, Ito Y, Yano E. Coronary aneurysms in infants and young children with acute febrile mucocutaneous lymph node syndrome. J Pediatr 1975; 86: 892898 6. Furukawa S, Matsubara T, Jujoh K et al. Peripheral blood monocyte/macrophages and serum tumor necrosis factor in Kawasaki disease. Clin immunol immunopathol 1988; 48:247-251 7. Kim DS. Serum interleukin-6 in Kawasaki disease. Yonsei Med J 1992; 33:183-188 8. Lang BA, Silverman ED, Laxer RM et al. Spontaneous tumor necrosis factor production in Kawasaki disease. J Pediatr 1989;115:939-943 9. Lang BA, Silverman ED, Laxer RM et al. Serum-soluble interleukin-2 receptor levels in Kawasaki disease. J Pediatr 1990; 116:592-596 10. Maury CPJ, Salo E, and Pelkonen P. Elevated circulating tumor necrosis factor-alfa in patients with Kawasaki disease. J Lab Clin Med 1989; 113:651-654 11. Leung DYM, Collins T, Lapierre LA, Geha RS, Pober JS. Immunoglobulin M antibodies present in the acute phase of Kawasaki syndrome lyse cultured vascular endothelial cells stimulated by gamma interferon. J Clin Invest 1986; 77:1428-1435. 32.
(33) 12.Leung DYM, Geha RS, Newberger JW, Burns JC, Fiers W, Lapierre LA, Pober JS. Two monokines, interleukin-1 and tumor necrosis factor, render cultured vascular endothelial cells susceptible to lysis by antibodies circulating during Kawasaki disease. J Exp Med 1986;164: 1958-1972 13. Maury CP, Salo E, Pelkonen P. Elevated circulating tumor necrosis factor-alpha in patients with Kawasaki disease. J Lab Clin Med 1989; 113:651-654 14. Matsubara T, Furukawa S, Yabuta K. Serum levels of tumor necrosis factor, interleukin 2 receptor, and interferon-gamma in Kawasaki disease involved coronary-artery lesions. Clin Immunol Immunopathol 1990; 56: 29-36 15. Lang BA, Silverman ED, Laxer RM, Lau AS. Spontaneous tumor necrosis factor production in Kawasaki disease, J Pediatr 1989; 115:939-943 16. Erdos EG, Skidgel RA. The unusual substrate specificity and the distribution of human angiotensin I converting enzyme. Hypertension 1986; 8:134-138 17. Caldwell PRB, Segal BC, Hsu KC, Das M, Soffer RL. Angiotensin converting enzyme: vascular endothelial localization. Science 1976; 191:1050-1051 18. Cambien F, Alhenc-Gelas F, Herbeth B, Andre JL, Rakotovao R, Gonzales MF, et al. Familial resemblance of plasma angiotensin-converting enzyme level: the Nancy study. Am J Hum Genet 1988; 43:774-780 19. Rigat B, Hubert C, Alhene-Gelas F, Cambien F, Corvol P, Soubrier F. An insertion/deletion polymorphism in the angiotensin I-converting enzyme gene accounting for half the variance of serum enzyme levels. J Clin invest 1990; 86:1343-1346 20. Alhenc-Gelas F, Richard J, Courbon D, Warnet JM, Corvol P. Distribution of plasma angiotensin I-converting enzyme levels in healthy men:Relationship to environmental and hormone parameters. J Lab Clin Med 1991; 117:33-39 21. Tiret L, Rigat B, Visvikis S, Breda C, Corvol P, Cambien F, et al. Evidence from combined. 33.
(34) segregation and linkage analysis, that a variant in the angiotensin-converting enzyme (ACE) gene controls plasma ACE levels. Am J Hum Genet 1992; 51:197-205 22. Harrap Sb, Davidson HR, Connor JM, Soubrier F, Corvol P, Fraser R, et al. The angiotensin-converting enzyme gene and predisposition to high blood pressure. Hypertension 1993; 21:455-460 23. Marre M, Bernadet P, Gallois Y, Savagner F, Guyene T-T, Hallab M, et al. Relationships between angiotensin-converting enzyme gene polymorphism, plasma levels, and diabetic retinal and renal complications. Diabetes 1994; 43:384-388 24. Foy CA, McCormack LJ, Knowler WC, Barrett JH, Catto A, Grant PJ. The angiotensinconverting enzyme gene I/D polymorphism and ACE levels in Pima Indians. J Med Genet 1996; 33:336-337 25. Winkleman BR, Nauck M, Klein B, Russ AP, Bohm BO, Siekmeier R, et al. Deletion polymorphism of angiotensin-converting enzyme gene is associated with increased plasma angiotensin-converting enzyme activity but not with increased risk for myocardial infarction and coronary artery disease. ANN Intern Med 1996; 125:19-25 26. Chiang FT, Lai ZP, Chern TH, Tseng CD, Hsu KL, Lo HM. Lack of association of the angiotensin-converting enzyme gene polymorphism with essential hypertension in a Chinese population. Am J Hypert 1997; 10:197-201 27. Morris BJ, Zee RYL, Schrader AP. Different frequencies of angiotensin-converting enzyme genotypes in older hypertensive individuals. J Clin Invest 1994; 94:1085-1089 28. Villard E, Soubrier F. Molecular biology and genetics of the angiotensin I-converting enzyme: potential implications in cardiovascular disease. Cardiovasc Res 1996; 3:25-29 29. Steinkasserer A, Spurr NK, Cox S, Jeggo P, Sim RB. The human IL -1 receptor antagonist gene (IL1RN) maps to chromosome 2q14-q21, in the region of the IL-1a and IL-1ß loci. Genomics 1992; 13:654-657. 34.
(35) 30. Di Giovine FS, Takhsh E, Blakemore AIF, Duff GW. Single base polymorphism at -511 in the human interleukin-1 ß gene. Hum Mol Genet 1992; 1:450 31. Pociot F, Molvig J, Wogegensen L, Worsaae H, Nerup J. A Taq I polymorphism in the human interleukin-1ß (IL-1 ß) gene correlates with IL -1ß secretion in vitro. Eur J Clin Invest 1992; 22:396-402 32. Arend W. Interleukin I receptor antagonist: a new member of the interleukin 1 family. J Clin Invest 1991; 5: 1445-1451 33. Tarlow JK, Blakemore AIF, Lennard A, Solari R, Hughes HN, Steinkasserer A, et al. Polymorphism in human IL-1 receptor antagonist gene intron 2 is caused by variable numbers of an 86 base pair tandem repeat. Hum Genet 1993; 91:403-404 34. Danis VA, Millington M, Hyland VJ, Grennan D. Cytokine production by normal human monocytes: inter-subject variation and relationship to an IL -1 receptor antagonist (IL -1Ra) gene polymorphism. Clin Exp Immunol 1995; 99:303-310 35. Le Beau MM, Lemons RS Espinosa R, Larson RA, Arai N, Rowley JD. Interleukin -4 and interleukin-5 map to human chromosome 5 in a region encoding growth factors and receptors and are deleted in myeloid leukemias with a del(5 q). Blood 1989; 73:647-650 36. Rosenwasser LJ, Klemm DJ, Dresback JK, Inamura H, Mascali JJ, Klinnert M, et al. Promoter polymorphisms in the chromosome 5 gene cluster in asthma and atopy. Clin Exp Allergy 1995; 25:74-78 37. Mout R, Wilemze R, Landegent JE. Repeat polymorphisms in the interleukin-4 gene. Nucleic Acids Res 1991; 19:3763 38.Olomolaiye O, Wood NA, Bidwell JL. A novel NIaIII polymorphism in the human IL -6 promoter. Eur J Immunogenet 1998; 25:267 39. Fishman D, Faulds G, Jeffery R, Mohamed9Ali V, Yudkin JS, Humphries S, Woo P. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and. 35.
(36) plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J Clin Invest 1998; 102:1369-1376 40. Takeuchi K, Yamamoto S, Kataoka S, Kakihara T, Tanaka A, Sato S, Uchiyama M. High incidence of angiotensin I converting enzyme genotype II in Kawasaki disease patients with coronary aneurysm. Eur J Pediatr 1997; 156:266-268 41. Kamizono S, Yamada A, Higuchi T, Kato H, Itoh K. Analysis of tumor necrosis factor-a production and polymorphisms of the tumor necrosis factor-a gene in individuals a history of Kawasaki disease. Pediatr Int 1999; 41:341-345 42. Kawasaki T. Acute febrile mucocutaneous syndrome with lymphoid involvement with specific desquamation of fingers and toes in children: clinical observations of 50 patients. Allerg 1967; 16:178-222 43. Fujiwara H, Hamashima Y. Pathology of the heart in Kawasaki disease. Pediatrics 1978; 61:100-7 44. Mattei MG, Hubert C, Alhenc-gelas F, Roekel N, Corvol P, Soubrier F. Angiotensin I converting enzyme gene is on chromosome 17. Cytogenet Cell Genet 1989; 51:1041 45. Rigat B, Hubert C, Alhenc-Gelas F, Cambien F, Corvol F, Corvol P, Soubrier F. An insertion/deletion polymorphism in the angiotensin I converting enzyme gene accounting for half the variance of serum enzyme levels. J Clin Invest 1990; 86:1343-6 46.Tiret L, Rigat B, Visvikis S, Breda C, Corvol P,Cambien F, Soubrier F. Evidence, from combined segregation and linkage analysis, that a variant of the angiotensin I converting enzyme (ACE)gene controls plasma ACE levels . Am J Hum Genet 1992; 51:197-205 47. Frossard PM, Lestringant GG, Obineche EN, Hill SH. The angiotensin-converting enzyme (ACE) gene insertion/deletion dimorphism tracks with higher serum ACE activities in both younger and older subjects. Ann Saudi Med 1998; 18: 289-392 48. Foy CA, Rice GI, Ossei-Gerning N, Mansfield MW, Grant PJ. Angiotensin I converting. 36.
(37) enzyme (ACE) gene polymorphism in patients characterized by coronary angiography. Hum Genet 1997; 100:420-5 49. Zhu X, Bouzekri N, Southam L, Cooper RS, et al. Linkage and association analysis of angiotensin I converting enzyme (ACE) gene polymorphisms with ACE concentration and blood pressure. Am J Hum genet 2001; 68:1139-48 50. Caldwell PRB, Segal BC, Hsu KC, Das M, Soffer RL. Angiotensin converting enzyme: vascular endothelial localization. Science 1976; 191:1050-1 51. Matucci Cerinic M, Jaffa A, Kahaleh BM. Angiotensin converting enzyme: An in vivo and in vitro marker of endothelial injury. J Lab Clin Med 1992; 120:428-33 52. Falcini F, Matucci Cerinic M, Ermini M, et al. Nerve growth factor circulating levels are increased in Kawasaki disease: correlation with disease activity and reduced angiotensin converting enzyme levels. J Rheumatol 1996; 23:1798-1802 53. Takeuchi K, Yamamoto K, Kataoka S, Kakihara T, Sato S, Uchiyama M. High incidence of angiotensin I converting enzyme genotype II in Kawasaki disease patients with coronary aneurysm. Eur J Pediatr 1997; 156:266-8 54. Rossi V, Breviario F, Ghezzi P, Dejana E, Montovani A. Prostacyclin synthesis induced by vascular cells by interleukin-1. Science 1985; 229:174-176 55. Demeter J, Messer G, Ramisch S, et al. Polymorphism within the second intron of the IL 1 receptor antagonist gene in patients with hematopoietic malignancies. Cytokines Mol Ther 1996; 2:239-242 56. Robertson SA, Seamark RF. The role of cytokines in gestation. Crit Rev Immunol 1994; 14:239-292 57. Copeland NG, Silan CM, Kingsley DM, et al. Chromosomal location of murine and human IL-1 receptor genes. Genomics 1991; 9:44-50 58. Mark LL, Haffajee AD, Socransky SS, et al. Effect of the interleukin-1 genotype on. 37.
(38) monocyte IL-1 beta expression in subjects with adult periodonitis. J Periodontal Res 2000; 35:172-177 59. Moos V, Rudwaleit M, Herzog V, Hohlig K, Sieper J, Muller B. Association of genotypes affecting the expression of interleukin-1 beta or interleukin-1 receptor antagonist with osteoarteritis. Arthritis Rheum 2000; 43:2417-2422 60. Pociot F, Molvig J, Wogensen L, et al. A Taq I polymorphism in the human interleukin-1ß secretion in vitro. Eur J Clin Invest 1992; 22:396-402 61. Santtila S, Savinainen K, Hurme M. Presence of the IL -1 RA allele 2(IL1RN 2) is associated with enhanced IL -1 beta production in vitro. Scand J Immunol 1998; 47:195198 62. Saltman DL, Dolganov GM, Warrington JA, Wasmuth JJ, Lovett M. A physical map of 15 loci on human chromosome 5 q23-q33 by two-color fluorescence in situ hybridization. Genomics 1993; 16:726-732 63. Fishman D, Faulds G, Jeffery R, Mohamedali V, Yudkin JS, Humphries S, Woo P. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and association with systemic-onset juvenile chronic arthritis. J Clin Invest 1998; 102:1369-1376 64. Nedwin GE, Naylor SL, Sakaguchi AY, Smith D, Jarrett-Nedwin J, Pennica D, Goeddel DV, Gray PW. Human lymphotoxin and tumor necrosis factor genes: structure, homology and chromosomal localization. Nucleic Acid Res 1985; 13:6361-6373 65. Louis E, Franchimont D, Piron A, Gevaert Y, Schaaf-Lafontaine N, Roland S, Mahieu P, Malaise M De Groote D, Louis R, Belaiche J. Tumor necrosis factor (TNF) gene polymorphism influences TNF-a production in lipopolysaccharide (LPS)-stimulated whole blood cell culture in healthy humans. Clin Exp Immunol 1998;113:401-406 66. McCartney-Francis NL, Wahl SM. Transforming growth factor ß: a matter of life and death.. 38.
(39) J Leukoc Biol 1994; 55:401-409 67. Border WA, Noble NA. Transforming growth factor ß in tissue fibrosis. N Engl J Med 1994; 10:1286-1292 68. Matsubara T, Umezawa Y, Tsuru S, Motohashi T, Yabuta K, Furukawa S. Decrease in the concentrations of transforming growth factor-beta 1 in the sera of patients with Kawasaki disease. Scand J Rheumatol 1997; 26:314-317 69. Awad MR, El-Gamel A, Hasleton P, Turner DM, Sinnott PJ, Hutchinson IV. Genotypic variation in the transforming growth factor- ß1 gene. Transplantation 1998; 66:1010-1020. 39.
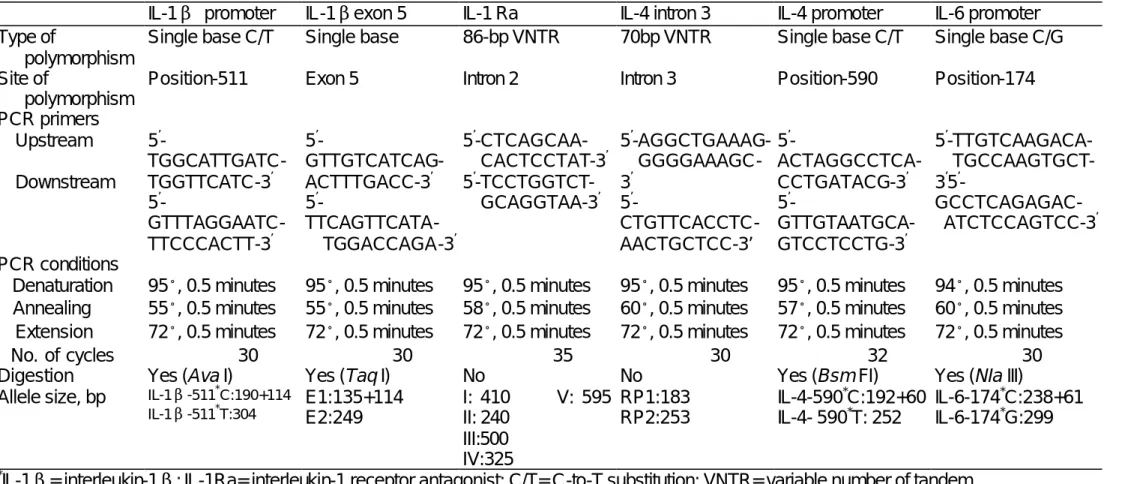
(40) Table Table 1. Main characteristics of cytokine gene polymorphisms and techniques used for screening * IL-1β promoter Single base C/T. Type of polymorphism Site of Position-511 polymorphism PCR primers Upstream 5’TGGCATTGATCDownstream TGGTTCATC-3’ 5’GTTTAGGAATCTTCCCACTT-3’ PCR conditions Denaturation 95。, 0.5 minutes Annealing 55。, 0.5 minutes Extension 72。, 0.5 minutes No. of cycles 30 Digestion Yes (Ava I) * IL-1β-511 C:190+114 Allele size, bp *. IL-1βexon 5 Single base. IL-1 Ra 86-bp VNTR. IL-4 intron 3 70bp VNTR. IL-4 promoter Single base C/T. IL-6 promoter Single base C/G. Exon 5. Intron 2. Intron 3. Position-590. Position-174. 5’-AGGCTGAAAGGGGGAAAGC3’ 5’CTGTTCACCTCAACTGCTCC-3’. 5’5’-TTGTCAAGACAACTAGGCCTCATGCCAAGTGCTCCTGATACG-3’ 3’5’5’GCCTCAGAGACGTTGTAATGCAATCTCCAGTCC-3’ ’ GTCCTCCTG-3. 5’5’-CTCAGCAAGTTGTCATCAGCACTCCTAT-3’ ’ ’ ACTTTGACC-3 5 -TCCTGGTCT’ 5GCAGGTAA-3’ TTCAGTTCATATGGACCAGA-3’ 95。, 0.5 minutes 55。, 0.5 minutes 72。, 0.5 minutes 30 Yes (Taq I) E1:135+114 E2:249. 95。, 0.5 minutes 95。, 0.5 minutes 95。, 0.5 minutes 94。, 0.5 minutes 58。, 0.5 minutes 60。, 0.5 minutes 57。, 0.5 minutes 60。, 0.5 minutes 72。, 0.5 minutes 72。, 0.5 minutes 72。, 0.5 minutes 72。, 0.5 minutes 35 30 32 30 No No Yes (Bsm FI) Yes (NIa III) I: 410 V: 595 RP1:183 IL-4-590*C:192+60 IL-6-174*C:238+61 IL-1β-511 T:304 II: 240 RP2:253 IL-4- 590*T: 252 IL-6-174*G:299 III:500 IV:325 * IL-1β= interleukin-1β; IL-1Ra= interleukin-1 receptor antagonist; C/T= C-to-T substitution; VNTR= variable number of tandem repeats; G/A=G-to-A substitution; PCR= polymerase chain reaction; SSOP= sequence-specific oligonucleotide probe. 40.
(41) Table 2. Distribution of ACE I/D genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects Kawasaki patients. Controls. P. Total=107 (%). Total=107 (%). Genotype ACE I/D. 0.008*. I/I. 30 (28.0). 37 (34.6). I/D. 71 (66.4). 52 (48.6). D/D. 6 (5.6). 18 (16.8). Allelic frequency ACE I/D. 0.693. I type. 131 (61.2). 126 (58.9). D type. 83 (38.8). 88 (41.1). *I: insertion. D: deletion Table 3. Distribution of ACE (A-240T) genotypes and allelic frequencies for patients suffering Kawasaki disease and also for healthy control subjects Kawasaki patients Total=90 (%). Controls. P. Total=100 (%). Genotype ACE (A-240T). 0.218. A/A. 36 (40.0). 29 (29.0). A/T. 46 (51.1). 57 (57.0). T/T. 8 (8. 9). 14 (14.0). Allelic frequency ACE (A-240T) Allele A Allele T. 0.113 118 (65.6) 62 (34.4). 41. 115 (57.5) 85 (42.5).
(42) Table 4. Distribution of ACE (G2350A) genotypes and allelic frequencies for patients suffering Kawasaki disease and also healthy control subjects Kawasaki patients. Controls. Total=107 (%). Total=100 (%). P. Genotype ACE(G2350A). 0.047*. A/A. 53 (49.5). 41 (41.0). A/G. 48 (44.9). 43 (43.0). G/G. 6 (5.6). 16 (16.0). Allelic frequency ACE(G2350A) Allele A Allele G. 0.046* 154 (72.0). 125(62.5). 60 (28.0). 75 (37.5). Table 5. Phenotype frequencies of the polymorphism in the ACE I/D gene in patients with and without coronary-artery aneurysm CAA -. P +. ACE. 0.404. I/I. 23. 7. I/D. 52. 19. D/D. 3. 3. *CAA : coronary artery aneurysm. 42.
(43) Table 6. Phenotype frequencies of the polymorphism in the ACE (A-240T) gene in patients with and without coronary-artery aneurysm. CAA. P. -. +. ACE. 0.197. AA. 29. 7. AT. 33. 13. TT. 4. 4. Table 7. Gene frequencies of the polymorphism in the ACE (A-240T) gene in patients with and without coronary-artery aneurysm. P. CAA -. +. ACE. 0.086. A. 91. 27. T. 41. 21. Table 8. Phenotype frequencies of the polymorphism in the ACE (G2350A) gene in patients with and without coronary-artery aneurysm CAA. P. -. +. ACE. 0.413. AA. 40. 13. AG. 35. 13. GG. 3. 3. 43.
(44) Table 9. Distribution of I L-1 ß promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Genotype. kawasaki p’t (n=107) n (%). normal (n=103). p value. n (%) 0.17 CC CT TT. 41(38.3) 45(42.1) 21(19.6). 27 (26.2) 52 (50.5) 24 (23.3). Allele frequency C T. 0.10 127(59.3) 87(40.7). 106 (51.5) 100 (48.5). Table 10. Phenotype and alleleic frequencies of the polymorphism in the IL -1 ß promoter gene in patients with and without coronary-artery aneurysm. Genotype. CAA(+) (n=29) n (%). CAA(-) (n=78) n (%). p value 0.75. CC CT TT. 10(34.5) 12(41.4) 7(24.1). 31(39.7) 33(42.4) 14(17.9). Allele frequency C T. 0.45 32(55.2) 26(44.8). 95(60.9) 61(39.1). 44.
(45) Table 11. Distribution of I L-1 ß exon 5 genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Genotype. kawasaki p’t (n=107) n (%). normal (n=103). p value. n (%) 0.27 a E1/E1 E1/E2. 105(98.1) 2(1.9). 98 (95.1) 5 (4.9). 212(99.1) 2(0.9). 201 (97.6) 5 (2.4). Allele frequency E1 E2 a:Fisher’s Exact Test. 0.28 a. Table 12. Phenotype and alleleic frequencies of the polymorphism in the IL -1 ß exon 5 gene in patients with and without coronary-artery aneurysm. Genotype. CAA(+) (n=29) n (%). CAA(-) (n=78) n (%). p value 0.47 a. E1/E1 E1/E2. 28(96.6) 1(3.4). 77(98.7) 1(1.3). 57(98.3) 1(1.7). 155(99.4) 1(0.6). Allele frequency E1 E2 a:Fisher’s Exact Test. 45. 0.47 a.
(46) Table 13. Distribution of I L-1 ß Ra genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Genotype. kawasaki p’t (n=107) n (%). normal (n=103). p value. n (%) 0.05 a I/I (410/410) I/II (410/240) II/II (240/240) I/IV (410/325) IV/IV (325/325) I/III(410/500). 89(83.3) 16(14.9) 1(0.9) 0 0. 95 (92.2) 6 (5.8) 1 (1.0) 0 1 (1.0). 1(0.9). 0. 195(91.1) 18(8.4) 1(0.5) 0. 196 (95.1) 8 (3.9) 0 2 (1.0). Allele frequency I/(410) II(240) III(500) IV(325) a:Fisher’s Exact Test. 0.05 a. Table 14. Phenotype and alleleic frequencies of the polymorphism in the IL -1 Ra gene in patients with and without coronary-artery aneurysm. Genotype. CAA(+) (n=29) n (%). CAA(-) (n=78) n (%). p value 0.14 a. I/I (410/410) I/II (410/240) II/II (240/240) I/IV (410/325) IV/IV (325/325) I/III(410/500). 24(82.8) 3(10.4) 1(3.4) 0 0. 65(83.3) 13(16.7) 0 0 0. 1(3.4). 0. 52(89.7) 5(8.6) 1(1.7) 0. 143(91.7) 13(8.3) 0 0. Allele frequency I/(410) II(240) III(500) IV(325) a:Fisher’s Exact Test. 46. 0.39 a.
(47) Table 15. Distribution of I L-4 promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Genotype. kawasaki p’t (n=107) n (%). normal (n=103). p value. n (%) 0.64 CC CT TT. 6(5.6) 42(39.3) 59(55.1). 6 (5.8) 34 (33.0) 63 (61.2). Allele frequency C T. 0.49 54(25.2) 160(74.8). 46 (22.3) 160 (77.7). Table 16. Phenotype and alleleic frequencies of the polymorphism in the IL -4 promoter gene in patients with and without coronary-artery aneurysm. Genotype. CAA(+) (n=29) n (%). CAA(-) (n=78) n (%). p value 0.51 a. CC CT TT. 1(3.4) 9(31.1) 19(65.5). 5(6.4) 33(42.3) 40(51.3). Allele frequency C T a:Fisher’s Exact Test. 0.20 11(19.0) 47(81.0). 43(27.6) 113(72.4). 47.
(48) Table 17. Distribution of I L-4 intron 3 genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Genotype. kawasaki p’t (n=107) normal (n=103) n (%) n (%). p value 0.66 a. RP1/RP1 RP1/RP2 RP2/RP2 RP0/RP1. 62(57.9) 38(35.6) 6(5.6) 1(0.9). 66 (64.1) 33 (32.0) 4 (3.9) 0. 163(76.2) 50(23.4) 1(0.4). 165 (80.1) 41 (19.9) 0. Allele frequency RP1 RP2 RP0 a:Fisher’s Exact Test. 0.38 a. Table 18. Phenotype and alleleic frequencies of the polymorphism in the IL -4 intron 3 gene in patients with and without coronary-artery aneurysm. Genotype. CAA(+) (n=29) n (%). CAA(-) (n=78) n (%). p value 0.30 a. RP1/RP1 RP1/RP2 RP2/RP2 RP0/RP1. 18(62.1) 8(27.6) 2(6.9) 1(3.4). 44(56.4) 30(38.5) 4(5.1) 0. 45(77.6) 12(20.7) 1(1.7). 118(75.6) 38(24.4) 0. Allele frequency RP1 RP2 RP0 a:Fisher’s Exact Test. 48. 0.30 a.
(49) Table 19. Distribution of IL -6 promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Genotype. kawasaki p’t (n=107) n (%). normal (n=103). p value. n (%) 1.00 a CC CG GG. 106(99.1) 1(0.9) 0. 34(100) 0 0 1.00 a. Allele frequency C G a:Fisher’s Exact Test. 213(99.5) 1(0.5). 68(100) 0. Table 20. Phenotype and alleleic frequencies of the polymorphism in the IL -6 promoter gene in patients with and without coronary-artery aneurysm. Genotype. CAA(+) (n=29) n (%). CAA(-) (n=78) n (%). p value 0.27 a. CC CG GG. 28(96.6) 1(3.4) 0. 78(100) 0 0. 57(98.3) 1(1.7). 156(100) 0. Allele frequency C G a:Fisher’s Exact Test. 0.27 a. Table 21. Distribution of TNF-a promoter genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects. Genotype. kawasaki p’t (n=107) n (%). normal (n=103). p value. n (%) 0.09 AA AG GG. 17(15.9) 24(22.4) 66(61.7). 7(6.8) 30(29.1) 66(64.1). Allele frequency A G. 0.17 58(27.1) 156(72.9). 44(21.4) 162(78.6). 49.
(50) Table 22. Phenotype and alleleic frequencies of the polymorphism in the TNFa gene in patients with and without coronary-artery aneurysm. Genotype. CAA(+) (n=29) n (%). p value. CAA(-) (n=78) n (%). 0.37 AA AG GG. 3(10.3) 5(17.3) 21(72.4). 14(17.9) 19((24.4) 45(57.7). Allele frequency A G. 0.10 11(19.0) 47(81.0). 47(30.1) 109(69.9). Table 23. Distribution of TGF-ß 1 genotypes and alleleic frequencies for patients suffering Kawasaki disease and also for healthy controls subjects Genotype. TGF-βF-1<-509> T/T C/T C/C Allelic frequency TGF-βF-1<-509> Allele T Allele C. Kawasaki patients total =107(%). Controls total = 100(%). P. 0.63 34(31.8) 54(50.5) 19(17.7). 31(31.0) 46(46.0) 23(23.0) 0.605. 122(58.0) 92(42.0). 108(54.0) 92(46.0). Table 24. Phenotype and alleleic frequencies of the polymorphism in the TGFß 1 gene in patients with and without coronary-artery aneurysm Genotype. Kawasaki patients total =107(%). TGF-βF-1<-509> T/T C/T C/C Allelic frequency TGF-βF-1<-509> Allele T Allele C. CAA(+) Total=29(%). CAA(-) total = 78(%). P. 0.934 34(31.8) 54(50.5) 19(17.7). 10(34.5) 14(48.3) 5(17.2). 24(30.8) 40(51.3) 14(17.9) 0.893. 122(58.0) 92(42.0). 34(58.6) 24(41.4). 50. 88(56.4) 68(43.6).
(51) Figure Fig. 1. ACE(intron16) M. II. ID. DD. 500bp. 495bp. 300bp 200bp 100bp. 208bp. Fig. 2. ACE(A-240T) A/T. T/T. A/A 137bp 114bp. Fig. 3. ACE(G2350A) AA AG GG AA 200bp 122bp 103bp 100bp. 51.
(52) Fig. 4. IL-1βpromoter TT. CT. CC. TT. M. 304bp 190bp 114bp. Fig. 5. IL-1β exon5 M E1/E2 E1/E1 E2/E2. 249bp 135bp 114bp. 52.
(53) Fig. 6. IL-1Ra M. M. 595bp 410bp 325bp 240bp. Fig. 7. IL-4 promoter. T/T. T/T. T/C. T/T. 252bp. T/C. C/C. 192bp. Fig. 8. IL-4 intron 3 M 500bp. 253bp 183bp 113bp. 53.
(54) Fig. 9. IL-6 promoter. GG. CG. CC. M. 299bp 238bp. Fig. 10. TNF-alpha A/A. G/G. A/G. G/G. 117bp 97bp. Fig. 11. TGF-ß1(-509) T/T. C/T. C/T. C/C. 120bp 74bp 46bp. 54.
(55) 簡單自述. 我出生於民風純樸、青山綠水的鄉城小鎮---彰化縣和美鎮,父 親是商人,母親是個家庭主婦,從小父母親疼愛子女,我們做子女 的都能體會父母的辛勞,也因為這樣從小就養成了獨立自主、積極 認真的個性。 年輕時求學過程非常順利,於民國六十五年進入中國醫藥學院 醫學系就讀。在大學時期,和同學相處融洽,我非常珍惜就讀醫學 院的機會,除了修習醫學相關科目外,也樂於學習新的事物。畢業 後返回母校附設醫院小兒科任職,歷任小兒科住院醫師、總醫師, 於民國七十八年升任小兒腸胃科主治醫師至今。然而,醫學進步一 日千里,這幾年來,母校在各項基礎醫學與臨床醫學的研究與發 展,投入極多的心力;為充實本身所學,期望能深入研究領域,擴 展更大的發展空間,於九十年進入臨床醫學研究所,接受更多的挑 戰與試鍊,希望能在醫學界學以致用。. .. 55.
(56)
數據



+7






Outline
相關文件
The percentage of positive LSGBs was significantly higher in patients in whom the biopsy was performed by or on the request of either the department of Rheumatology and Internal
pylori in pathogenesis of oral mucosal lesions or ulcerations is still unclear, it seems that patients with oral lesions as leukoplakia and oral lichen planus, and concurrent
However, in HIV-positive patients, plasma cell tumors may present at unusual sites and progress rapidly to involve multiple sites, including the soft tissues and viscera [19]..
According to his protocol, patients are categorised as Stage I if they exhibit exposed bone in a field of radiation that has failed to heal for at least 6 months and do not have
Genetic study of the BRAF gene reveals new variants and high frequency of the V600E mutation among Iranian ameloblastoma patients.. Maryam Soltani 1 | Mohammad Amin Tabatabaiefar 2,3
Radiomorphometric indices can be used to deter- mine the existence of a porous structure in the man- dible on panoramic images of patients who have scleroderma and may have a high
In conclusion, data from the present study demonstrat- ed that signs of carotid calcifications in panoramic radio- graphs are associated with future events of stroke and/or
A discussion of the Dapagli flozin and Prevention of Adverse Outcomes in Chronic Kidney Disease (DAPA-CKD) trial, the Effect of Sotagli flozin on Car- diovascular Events in Patients
