行政院國家科學委員會專題研究計畫 成果報告
結合運動及可食多醣體調節人體周邊血液單核球細胞促進
HL-60 白血病細胞株分化之研究(I)
研究成果報告(精簡版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2413-H-028-003- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣體育學院體育學系 計 畫 主 持 人 : 陳裕鏞 共 同 主 持 人 : 江啟銘、陳裕仁、廖慧芬 計畫參與人員: 研究生:王識豪、施怡甄、簡宸妮 處 理 方 式 : 本計畫涉及專利或其他智慧財產權,2 年後可公開查詢中 華 民 國 96 年 11 月 29 日
Competitive cyclists possess greater immunomodulatory response
against surface antigen expression of human hepatitis B virus
Yu-Yawn Chen
aa
Department and Graduate School of Physical Education, National Taiwan College of Physical Education, Taichung, Taiwan
*Corresponding author. Phone: (886) 2 2809 4661 ext. 2301; Fax: (886) 2 2809 6180 E-mail: [email protected]
Department of Radiation Oncology, Mackay Memorial Hospital 92 Chung San North Road, Taipei 104, Taiwan
Abstract
To evaluate whether competitive cyclists with moderate training have higher immunity against hepatitis B virus than sedentary controls was present purpose. Human peripheral blood mononuclear cells, from competitive cyclists and sedentary controls, were stimulated by phytohemagglutinin (PHA) to prepare conditioned media (MNC-CM) for assessment of the inhibitory effects on hepatitis B surface antigen (HBsAg) expression in human hepatoma Hep3B cells. Our results showed that the inhibitory effect against the relative HBsAg expression of cyclists’ MNC-CM was greater than that of control. For example, the cyclists’ MNC-CM prepared from 5 μg/ml PHA decreased HBsAg expression to 61.5% whereas that of controls remained at 83.8%. The levels of cytokines such as IFN-γ, TNF-α, IFN-α and IL-1β in the MNC-CM from the cyclists increased in a PHA dose-dependent manner and were higher than those in the control group. Antibody neutralization of cyclists’ MNC-CM and addition of recombinant cytokines into controls’ MNC-CM indicated that IFN-γ, TNF-α and IFN-α had synergistic effects against HBsAg expression. Our results suggest that the immunomodulatory response to suppress HBsAg expression in competitive cyclists with moderate training is greater than that in sedentary controls.
Introduction
The prevalence of chronic hepatitis B, with more than 350 million patients in the world and 3 million in Taiwan, is progressively increasing worldwide (Kao and Chen, 2002; Kane, 1996). Moreover, hepatitis B virus (HBV) is highly correlative to the formation of hepatocellular carcinoma (Beasley, 1988). The clinical outcome of hepatocellular carcinoma is poor, especially in patients with unresectable disease while diagnosis. Clearly, it is important to develop better strategies against HBV infection.
People who exercise regularly are often thought to have resistance to many diseases, including virus infection. However, some epidemiological investigations have shown higher incidences of upper respiratory infection after prolonged, strenuous exercise during competition season (Nieman et al., 1990; Peters et al., 1983, 1993). By contrast, moderate exercise resulted in a decreased upper respiratory tract infection rate (Nieman et al., 1990). The stimulating effects on the immune system against virus can be enhanced by exercise with moderate intensity. For example, after moderate exercise intervention, elderly subjects who were immunized with trivalent influenza vaccine responded with a higher antibody titer to influenza in their blood than that seen in controls (Kohut et al., 2004). The resistance to infection of herpes simplex virus type 1 was also proposed to be associated with moderate exercise in mice (Davis et al., 2003).
The attempt of this study was to examine the difference from immune reaction against HBV activity between competitive cyclists and sedentary controls. PBMNC were isolated and stimulated with different concentrations of PHA, mimicking the immune
reaction induced in vivo. In this study we proposed that moderate exercise may have greater immunomodulatory response to inhibit gene expression of HBV in competitive cyclists. Whether this effect was exerted by the secreted cytokines from PBMNC was also tested.
Methods
Subjects
After review by the Human Ethics Committee of Chinese Culture University, twelve competitive cyclists and fourteen sedentary controls (SC) were recruited to participate in this study with their informed consents. All subjects were male with a mean age of 22.9 ± 2.7 years (range19 to 26 years old), a mean height of 173 ± 4 cm, and a mean mass of about 65.2 ± 3.4 kg. There was no difference in these parameters between the cyclist group and sedentary controls (Table 1). During the experimental period, no subject participated in any competitions or intensive training programs. The athletes cycled underwent no more than 120 km per week for 4 months prior to the study. Before blood collection, the cyclists in the moderate training group (CMT) underwent a moderate training program for 4 weeks. They exercised on a bicycle ergometer with work intensity at an individually estimated V‧O2max of
65% for 60 min every day between 8 and 11 o’clock A.M. in an average room temperature at 24 ºC. 60 min of exercise was divided into four 15-min intervals with 5 min of rest in each interval. During the experimental period, the SC group subjects stayed in the same place and were seated to watch a video. The subjects were provided with a breakfast consisting both of 240 ml of orange juice and milk, one egg and six slices of white toast 1 hour before the moderate training program. All subjects refrained from ingesting any drugs, alcoholic drinks, caffeinated beverages, smoking and vegetarian diet for 4 weeks prior to the study, as well as during the experimental period. The subjects had no clinical illnesses or surgical treatment during the 4 weeks prior to blood collection and the experimental period. All of the subjects
are neither HBV carriers nor hepatitis B surface antigen (HBsAg) positive, which were examined by physician. Each fasting blood sample was taken between 8 and 9 o’clock A.M. after the subjects had rested quietly for at least 48 hours. Serum albumin concentrations were determined by the Bromcresol Green method.
Measurements of cardiopulmonary fitness
Test of maximal oxygen uptake (V‧O2max) were completed two weeks before experimental period using an initial incremental maximal exercise test. In brief, maximal oxygen uptake and ventilation ability were measured by a system for cardiopulmonary exercise testing (Q-plus IW/Corival 400, Seattle, WA, USA) while the subjects were exercising on their own bicycles mounted on and electronically braked cycle ergometer (Lode Excalibur, Quinton Instruments, Seattle, WA, USA). The subjects warmed up by loading set at 75 W for 2 min and then began to increase loading set at 25 W every 2 min until volitional fatigue. A polar pacer heart rate monitor was used to record the heart rate.
Preparation of mononuclear cell-conditioned media (MNC-CM)
Human MNC were taken from the peripheral blood of each healthy volunteer with informed consent. The cells were separated by density centrifugation (400 g, 30 min) in a Ficoll-Hypaque solution (1.077 g/ml) as previously described (Chiang et al., 2000). MNC were washed three times in phosphate-buffered saline, suspended in RPMI-1640 medium (Gibco/BRL, Grand Island, NY, USA) containing 10% heat-inactivated autoserum, and seeded on autoserum-coated culture plates. Cells were cultured at an initial concentration of 1
x 106 cells/ml in 10% heat-inactivated fetal calf serum (Hyclone, Logan, UT) with 50 mg/l penicillium (Gibco/BRL), 100 mg/l streptomycin (Gibco/BRL), 1,000 micromole/l L-glutamine (Gibco/BRL), and RPMI 1640 medium containing a series of levels (0, 1.25, 2.5 and 5 μg/ml) of phytohemagglutinin (PHA; Difco Lab., Detroit, MI, USA) at 37 oC in a fully humidified incubator with 5% CO2. After cultivation for 24 hours, the collected aliquot was filtered through a 0.45 μ membrane to remove MNC and then stored at -80 oC until use (Wang et al., 1997). This was termed conditioned media (CM). Subsequently, PHA-MNC-CM prepared with 5 μg/ml PHA for 24 hours, were used to observe the relative HBsAg expression in Hep3B cells using the method described below. For cytokine assay, MNC-CM prepared with 5 μg/ml PHA was collected. Hep3B cell medium, in which a volume of only 30% PBS with 5 μg/ml PHA replaced the MNC-CM, was termed the untreated control group.
PHA is a natural mitogen of T lymphocytes isolated from plants. It has been used as an immune stimulant in various studies regarding immune response and effect. For example, in our previous studies, we stimulated PBMNC with gradient concentrations of PHA to mimic various degree of immune reaction for evaluation of drug-induced (Chen and Chang, 2004; Chen, 1997), exercise-mediated immunomodulation (Chiang et al., 2000; Liao et al., 2006). The basis of this model is that PHA stimulates production and release of various cytokines from PBMNC including those relate to anti-viral and anti-tumor immunity. Among which, IFN-γ, TNF-α and IFN-α have been shown as effector molecules acting on anti-HBV immunity (Suri et al., 2001; Heike et al., 2000; Geneva-Popova et al., 1999). It follows that we use PHA as an immune stimulant and cytokine levels in PHA-MNC-CM as
a measure of anti-HBV effectors.
Hepatoma cell cultures
Hep3B/C16 (Hep3B), a moderately differentiated human hepatocellular carcinoma cell line, which is integrated in the HBV genome in its chromosome and stably produces HBsAg, was taken as the cell model for studying HBV replication in our study (Knowles et al., 1980; Twist et al., 1981). The Hep3B cells were cultured at an initial concentration of 1 x 106 cells/ml in Dulbecco's modified Eagle’s medium (DMEM) containing 10% fetal calf serum (Gibco/BRL), 1 X 105 I.U./l penicillin, 100 mg/l streptomycin, and 1,000 micromole/l L-glutamine, in a humidified 5% CO2 incubator at 37 oC. The 30% volumes of conditioned medium from MNC and fresh DMEM plus 10% fetal calf serum were added and incubated for 3 days in a humidified 5% CO2 incubator at 37 oC. In our pretest experiment, we found that the addition of 30% CM is optimal for Hep3B cell to grow. The final concentration of PHA was 0 - 1.5 μg/ml in the MNC-CM, which had no direct effect on the relative HBsAg expression of Hep3B cells tested in our preliminary work. The cell viability was assessed by a tetrazolium dye colorimetric MTT test (Mosmann, 1983)and expressed as follows: MTT value of experimental group/MTT value of untreated control group.
Assay for relative HBsAg expression
Hep3B cells cultured in DMEM with 10% fetal bovine serum for 24 hrs were transferred to serum-free DMEM with or without 30% (v/v) MNC-CM and incubated for 48 hrs. The secreted HBsAg in the culture medium was measured by commercial ELISA kits (General
Biological, Taipei, Taiwan, Republic of China). The determined optical density (O.D.) values of the Enzyme-Linked Immunosorbent Assay (ELISA) kits during measurement were normalized with cell numbers. PHA does not interfere with the HBsAg assay. The detection of HBsAg in serum of patients indicated a current HBV infection and the risk for developing compensated cirrhosis and hepatocellular carcinoma (Chisari, 1997; Lee, 1997; Yang et al., 1996). Thus, the HBsAg is a useful index to evaluate the viral activity (Czaja, 1979). The relative HBsAg expression was determined by following formula: (HBsAg/MTT) from PHA-MNC-CM / (HBsAg/MTT) from the untreated control group culture media. The (HBsAg/MTT) from the untreated control group culture media was treated as 100% expression. The measurement of secreted viral antigens from HBV host cells, such as HBsAg secretion from Hep3B cells has been applied in various investigations to evaluate the effect of cytokine (Lin et al., 2003) drugs (Chen et al., 1996; Chen et al.,
1995)on HBV activity.
Assay for cytokines
To detect the effects of cytokines on the reduction of relative HBsAg expression in Hep3B incubated with PHA-MNC-CM, commercial ELISA kits for IL-1β, TNF-α, IFN-α, and IFN-γ (R&D Systems, Minneapolis, MN, USA) were used and determined at a wavelength of 450 nm according to the method described by Wang et al. (1997). The correlation coefficients (r2) for the standard curves of three cytokines were between 0.998-0.999. Three separate experiments were each tested in duplicate.
Antibody neutralization
PHA-MNC-CM from the cyclists group (termed as CMT-PHA-MNC-CM) were pre-incubated at 37 °C for 90 min with various cytokine-neutralizing antibodies including anti-IFN-γ (30.0 μg/ml, is 10 folds more over than the concentration as near 100% of
neutralization), anti-TNF-α (2.4 μg/ml), anti-IFN-α (1.0 μg/ml) and anti-IL-1β (5.1 μg/ml) in combination or alone. PHA-MNC-CM from the sedentary control group was termed SC-PHA-MNC-CM. Viable cells were counted after 48 hours of incubation with added antibodies of the cytokines. Three separate experiments were each tested in duplicate.
Addition of IFN-γ, TNF-α and IFN-α into SC-PHA-MNC-CM
SC-PHA-MNC-CM were pre-incubated at 37 °C for 24 hours with various cytokines including IL-1β (500 pg/ml), TNF-α (1200 pg/ml), IFN-α (700 pg/ml), and IFN-γ (1000
pg/ml) or their combinations.
Statistical analysis
Results are presented as mean ± SEM. Differences between the treatment groups, which consisted of matched samples, was assessed by Student’s t test. A confidence level of 5% (p < 0.05) was considered significant.
Results
The maximal oxygen uptake (V‧O2max)
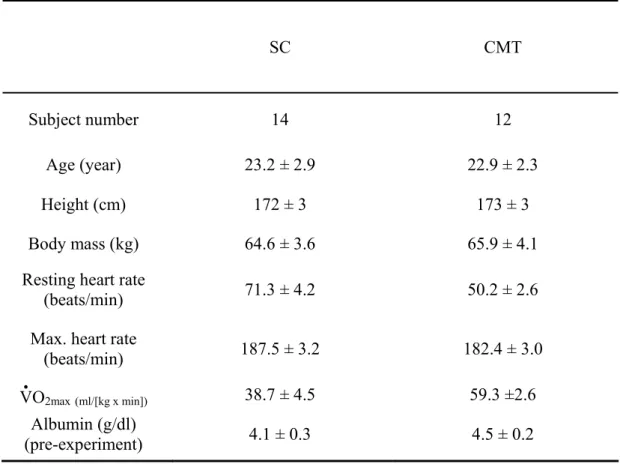
As shown in Table 1, the mean V‧O2max of the SC group was 38.7 ± 4.5 ml/kg/min (range 35.6
- 42.1) and of CMT group was 59.3 ml/kg/min (range 56.8 – 62.2), having a significant difference (p < 0.001). There was no significant difference in age, height and body mass between groups (p > 0.05). There were no significant changes in mean V‧O2max (data not
shown) or albumin level in the CMT group after the experimental intervention.
Comparison of the reduction of relative HBsAg expression in Hep3B cells stimulated by PHA-MNC-CM from different groups
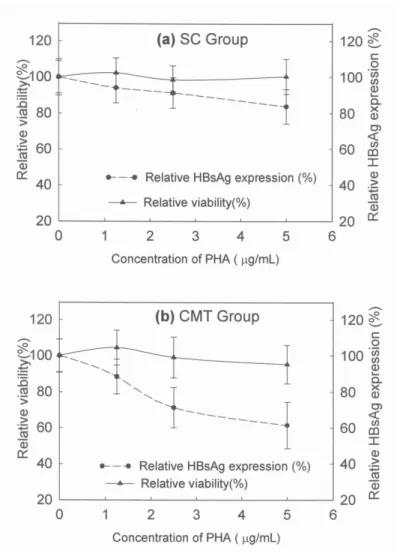
The relative HBsAg expression in Hep3B cells after stimulation with PHA-MNC-CM decreased with increasing doses of PHA, and a dose-response effect was much more obvious in the CMT group. For example, in comparison with the SC group, there was a much lower relative HBsAg expression of 61.5% of HBV in Hep3B cells incubated with CMT-PHA-MNC-CM prepared by stimulation with PHA at the same 5 μg/ml (Figure. 1b). No obvious cytotoxcity to Hep3B cells was observed in both groups when the administered concentrations were lower than 5 μg/ml. However, no significant reduction of relative HBsAg expression in Hep3B was observed in the SC group (Figure. 1a). After stimulation by PHA at 5 μg/ml for 24 hours, the amounts of IFN-γ, IL-1β, TNF-α and IFN-α in CMT-PHA-MNC-CM all dramatically increased (1345 ± 196 pg/ml, 830 ± 189 pg/ml, 1937 ± 314 pg/ml and 947 ± 175 respectively), compared to those in SC-PHA-MNC-CM (425 ±
146 pg/ml, 352 ± 103 pg/ml, 756 ± 238 pg/ml and 326 ± 118 pg/ml, respectively).
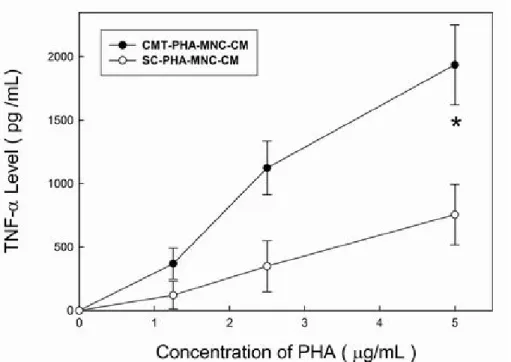
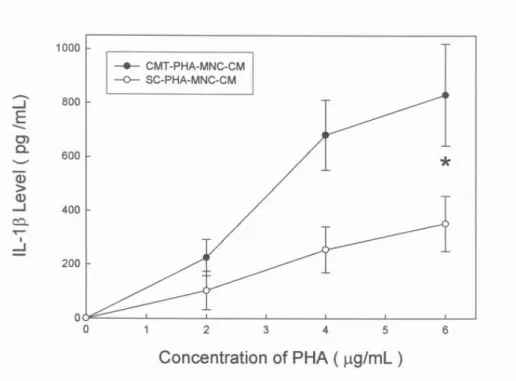
The significant augmentation of secreted IFN-γ from 58 ± 41 to 1345 ± 196 pg/ml occurred as the amount of PHA increased from 0 μg/ml to 5 μg/ml in CMT-PHA-MNC-CM (Figure. 2). The production of IFN-γ in CMT-PHA-MNC-CM prepared by stimulating with 5 μg/ml PHA was about 3 folds higher than that in the SC-PHA-MNC-CM. Although the increment of TNF-α production was much higher than that of IFN-γ, the trend in the increment was similar. The concentration of TNF-α in CM from the CMT group increased exponentially, ranging from about 369 ± 122 to 1937 ± 314 pg/ml (Figure. 3). The TNF-α production in CMT-PHA-MNC-CM was about 2 fold higher than that in the SC-PHA-MNC-CM. The concentrations of IFN-α in CMT-PHA-MNC-CM increased exponentially, ranging from about 80 ± 59 to 947 ± 175 pg/ml (Figure. 4). With a similar trend, the concentration of IL-1β in CMT-PHA-MNC-CM increased exponentially, ranging from about 225 ± 67 to 830 ± 189 pg/ml (Figure. 5).
Discussion
In this study, we demonstrated that PBMNC isolated from competitive cyclists possessed greater immunomodulatory response against HBsAg expression of HBV in an ex
vivo experimental model. This effect may be attributed to much more secretion of cytokines,
namely TNF-α, IFN-α and IFN-γ.
The exercise intensity of cyclists assessed in our study was moderate. There was much lower relative HBsAg expression of HBV in Hep3B cells incubated with
CMT-PHA-MNC-CM than in SC-PHA-MNC-CM. Lowder also pointed out that moderate exercise training inhibited activity of influenza virus in vivo (Lowder et al., 2005 ). Thus, moderate intensity exercise might be a suitable way to enhance anti-viral immunity.
CM was prepared by removal of MNC after stimulation by various concentrations of PHA. This suggests that there may be soluble mediators produced by MNC capable of inhibiting HBsAg expression in Hep3B cells. In our study, the secreted levels of IFN-γ, TNF-α, IFN-α and IL-1β of the CMT-PHA-MNC-CM were much higher than those of the SC-PHA-MNC-CM. The CMT-PHA-MNC-CM neutralized by anti-IFN-γ, anti-TNF-α and anti-IFN-α antibodies (but not anti-IL-1β) partially lost the inhibitory activity against relative HBsAg expression. Both these results of experiments of antibody neutralization and cytokines supplement show that IFN-γ, TNF-α, IFN-α, but not IL-1β, might be the soluble mediators contributing to the greater inhibitory activity of CMT-MNC-CM. Furthermore, our results showed that moderate exercise training, such as that for cyclists, are safe for cultured MNC because there was no cytotoxicity to MNC observed (data not shown).
Clinical studies pointed out that IFN-γ exert the HBsAg inhibitory effects (Bonino et al., 1986; Mahrle and Schulze, 1990). A similar study about active experienced older runners showed that moderate exercise can elevate the blood IFN-γ concentration after being stimulating with PHA (Shinkai et al., 1995). Both results of much higher IFN-γ in CMT-PHA-MNC-CM and lower relative HBsAg expression with anti-IFN-γ antibody neutralization have shown the important role played by IFN-γ. The degree of cell cytoplasmic HBV DNA reduction was highly correlated with the level of secretion of
IFN-γ in the supernatants of PBMNC medium (Schultz and Chisari, 1999; Sheron et al., 1991). TNF-α is considered as an activator of antiviral effects in the immune system. The administration of TNF-α disrupt the formation of cytoplasmic viral capsids to causes noncytopathic suppression of hepatitis B virus DNA replication in hepatocytes (Biermer et al., 2003). When TNF-α was added to Hep3B cells, a significant reduction of HBV replication was noted (Romero and Lavine, 1996). Both results of much higher TNF-α and lower relative HBsAg expression with anti-TNF-α antibody neutralization in CMT-PHA-MNC-CM have shown the critical role played by TNF-α. The anti-virus
activity exerted with increased secretion of IFN-α has been observed in other studies with similar experimental models. After being cultured with MNCs combined with antibody of IFN-α in Herpes simplex viruses (HSV)-infected human embryo lung fibroblasts, the level of viral suppression was reduced (Leibson et al., 1986). In fact, the effects of IFN-α on the inhibition of expression of HBV virus have been used in clinical therapy. The treatment of HBeAg-positive patients with IFN-α led to loss of HBeAg in 25 to 40% (Perrillo et al., 1990; Van Zonneveld et al., 2004). With both results of much higher IFN-α and lower relative HBsAg expression with anti-IFN-α antibody neutralization in CMT-PHA-MNC-CM, it showed that the obvious role played by IFN-α. Although the greater secretion of cytokines accompanied by reduced expression of HBsAg, no
cytotoxic effect of CMT-PHA-MNC-CM against the host HBV-harboring Hep3B cells was observed in our study. It indicates that the inhibitory activity of CMT-MNC-CM against HBsAg expression may not come from the cytotoxicity to Hep3B cells. It has been reported
that HBV-specific cytotoxic T lymphocytes can secrete both the TNF-α and IFN-γ to abolish HBV gene expression and replication without cytopathy (Chisari, 1997). The synergistic reduction of the inhibitory effect against HBsAg expression in CMT-PHA-MNC-CM by administration both of anti-TNF-α and anti-IFN-γ antibodies demonstrate that the interaction between them also attribute to the anti-viral activity. A combination of anti-TNF-α, anti-IFN-α and anti-IFN-γ exhibited the greatest blocking effect on inhibition against relative HBsAg expression in CMT-PHA-MNC-CM, therefore, it provides further support to speculate the main immunomodulatory anti-viral role played by above three cytokines during moderate exercise.
Whether the HBV-specific cytotoxic T lymphocytes play a role in the MNC we tested remains to be elucidated. The major reason why we used MNC, but not each single isolated cell lineages, in this study is that existence of all the various kinds of cells in MNC may represent a better simulation for the immune cell interaction inside human body.
In conclusion, by using an ex vivo antiviral immunity model, we found that competitive cyclists possessed greater immunomodulatory response against HBsAg expression of HBV. This effect may be attributed to much more secretion of cytokines, namely IFN-γ, TNF-α and IFN-α from human peripheral blood MNC.
Acknowledgements
This study was supported by MMH-94100 and MMH-E-95006 from Mackay Memorial Hospital, and NSC 95-2413-H-028-003 from the National Science Council of Republic of China.
References
Beasley, R. P. Hepatitis B virus. (1988) The major etiology of hepatocellular carcinoma.
Cancer 61, 1942-1956.
Biermer M, Puro R and Schneider RJ. (2003) Tumor necrosis factor alpha inhibition of hepatitis B virus replication involves disruption of capsid integrity through activation of NF-kappaB. Journal of virology 77, 4033-4042.
Biron, CA, Nguyen KB, Pien GC, Cousens LP and Salazar-Mather TP. (1999) Natural killer cells in antiviral defense: function and regulationby innate cytokines. Annual
review of immunology. 17, 189-220.
Bonino, F., Rosina, F., Rizzetto, M., Rizzi, R., Chiaberge, E., Tardanico, R., Callea, F. and Verme, G., (1986) Chronic hepatitis in HBsAg carriers with serum HBV-DNA and anti-HBe. Gastroenterology 90, 1268-1273.
Chen HC, Chou CK, Lee SD, Wang JC and Yeh SF. (1995) Active compounds from Saussurea lappa Clarks that suppress hepatitis B virus surface antigen gene expression in human hepatoma cells. Antiviral Res. 27(1-2), 99-109.
Chen HC, Chou CK, Sun CM and Yeh SF. (1997) Suppressive effects of destruxin B on hepatitis B virus surface antigen gene expression in human hepatoma cells.
Antiviral research. 34(3), 137-44.
Chen YJ, Shiao MS, Lee SS and Wang SY. (1997) Effect of Cordyceps Sinensis on the proliferation and differentiation of human leukemic U937 cells. Life sciences 60(25), 2349-59.
Chen YY and Chang HM. (2004) Antiproliferative and differentiating effects of polysaccharide fraction from Fu-Ling (Poria cocos) on human leukemic U937 and HL-60 cells source. Food and chemical toxicology 42(5),759-769.
Chetverikova LK, Frolov BA, Kramskaya TA and Polyak RYa. (1987) Experimental influenza infection: influence of stress. Acta virologica 31, 424–433.
Chiang J, Huang YW, Chen ML, Wang SY, Huang AC and Chen YJ. (2000) Comparison of anti-leukemic immunity against U937 cells in endurance athletes versus sedentary controls International journal of sports medicine. 21, 602-607.
Chisari FV. (1997) Cytotoxic T cells and viral hepatitis. The Journal of clinical
investigation. 99, 1472-1477.
Czaja AJ. (1979) Serologic markers of hepatitis A and B in acute and chronic liver disease.
Mayo Clinic proceedings. Mayo Clinic. 54, 721-732.
Davis JM, Murphy EA, Brown AS, Carmichael MD, Ghaffa, A and Maye EP. (2003) Effects of moderate exercise and oat β-glucan on innate immune function and susceptibility to respiratory infection. American journal of physiology. Regulatory,
integrative and comparative physiology. 286, R366–R372.
Geneva-Popova M and Murdjeva M. (1999) Study on proinflammatory cytokines (IL-1 beta, IL-6, TNF-alpha) and IL-2 in patients with acute hepatitis B. Folia Medica
(Plovdiv) 41, 478-481.
Haria M and Benfield P. (1995) Interferon-alpha-2a review of its pharmacological properties and therapeutic use in the management of viral hepatitis. Drugs 50, 873-896.
Heike Mcclary, Rick Koch, Francis V. Chisari and Luca G. Guidotti. (2000) Relative sensitivity of hepatitis B virus and other hepatotropic viruses to the antiviral effects of cytokines.. Journal of virology 74, 2255–2264.
Kane M. A. (1996) Global status of hepatitis B immunisation. Lancet 348, 696.
Kao, J. H., and Chen, D. S., (2002) Global control of hepatitis B virus infection. The Lancet Infectious Diseases 2, 395-403.
Knowles BB, Howe CC and Aden DP. (1980) Human hepatocellular carcinomacell lines secrete the major plasma proteins and hepatitis B surface antigen. Science 209, 497-501.
Kohut ML, Arntson BA, Lee W, Rozeboom K, Yoon KJ, Cunnick JE and McElhaney J. (2004) Moderate exercise improves antibody response to influenza immunization in older adults. Vaccine 22, 2298-2306.
Lee WM. (1997) Hepatitis B virus infection. The New England journal of medicine 337, 1733-1745.
Leibson PJ, Laszlo MH and Hayward AR. (1986) Inhibition of herpes simplex virus type 1 replication in fibroblast cultures by human blood mononuclear cells. Journal of
hepatology 57(3), 976-982.
Liao HF, Chiang LM, Yen CC, Chen YY, Zhuang RR, Lai LY, Chiang J and Chen YJ. (2006) Effect of a periodized exercise training and active recovery program on antitumor activity and development of dendritic cells. The Journal of sports medicine and
physical fitness 46,307-314.
Lin SJ, Shu PY, Chang C, Ng AK and Hu CP. (2003) IL-4 suppresses the expression and the replication of hepatitis B virus in the hepatocellular carcinoma cell line Hep3B.
Journal of immunology. 171(9),4708-16.
Lowder T, Padgett DA and Woods JA. (2005) Moderate exercise protects mice from death due to influenza virus. Brain, behavior, and immunity 19(5), 377-380.
Mahrle G and Schulze HJ. (1990) Recombinant interferon-gamma (rIFN-gamma) in dermatology. The Journal of investigative dermatology. 95, 132S-137S.
Moldoveanu AI, Shephard RJ and Shek PN. (2000) Exercise elevates plasma levels but not gene expression of IL-1beta, IL-6, and TNF-alpha in blood mononuclear cells.
Journal of applied physiology 89(4), 1499-1504.
Mosmann T. (1983) Rapid colorimetric assay for cellular growth and survival, Application to proliferation and cytotoxicity assays. Journal of immunological methods 69, 55-63.
Nieman, D.C., Johanssen, L.M., Lee, J.W., Cermak, J. and Arabatzis, K. (1990) Infectious episodes in runners before and after the Los Angeles Marathon. The Journal of
sports medicine and physical fitness 30, 316–328.
Nieman DC, Nehlsen-Cannarella SL and Markoff PA. (1990) The effects of moderate exercise training on natural killer cells and acute upper respiratory tract infections.
International journal of sports medicine 11, 467-473.
Perrillo RP, Schiff ER and Davis GL. (1990) A randomized, controlled trial of interferon alfa-2b alone and after prednisone withdrawal for the treatment of chronic hepatitis B: the hepatitis interventional therapy group. New England journal of
medicine 323(5), 295-301.
582–584.
Peters EM, Goetzche JM, Brobbelaar B and Noakes TD. (1993) Vitamin C supplementation reduces the incidence of postrace symptoms of upper respiratory infection in ultramarathon runners. The American journal of clinical nutrition 57, 170–174. Ren F., Hino K.,Yamaguchi Y., Funatsuki K., Hayashi A., Ishiko H., Furutani M., Yamasaki,
T., Korenaga K., Yamashita S., Konishi T. and Okita K. (2003) Cytokine-dependent anti-viral role of CD4-positive T cells in therapeutic vaccination against chronic hepatitis B viral infection. Journal of medical virology. 71, 376-384.
Reyes MP and Lerner AM. (1976) Interferon and neutralizing antibody in sera of exercised mice with coxsackievirus B-3 myocarditis. Proceedings of the Society for
Experimental Biology and Medicine. Society for Experimental Biology and Medicine 151, 333–338.
Romero R and Lavine JE. (1996) Cytokine inhibition of the hepatitis B virus core promoter. Hepatology 23, 17-23.
Schindler C and Brutsaert S. (1999) Interferons as a paradigm for cytokine signal transduction. Cellular and molecular life sciences 55, 1509-1522.
Schultz U and Chisari FV. (1999) Recombinant duck interferon gamma inhibits duck hepatitis B virus replication in primary hepatocytes. Journal of virology. 73, 3162-3168.
Sheron, N., Lau, J., Daniels, H., Goka, J., Eddleston, A., Alexander, G.J. and Williams, R (1991) Increased production of tumour necrosis factor alpha in chronic hepatitis B virus infection. Journal of hepatology 12, 241-245.
Shinkai, S., Kohno, H., Kimura, K., Komura, T., Asai, H., Inai, R., Oka, K., Kurokawa, Y. and Shephard, R. (1995) Physical activity and immune senescence in men.
Medicine and science in sports and exercise. 27(11), 1516-1526.
Solomon GF, Merigan TC and Levine S. (1966) Variation in adrenal cortical hormones within physiologic ranges, stress and interferon production in mice. Rivista di
patologia nervosa e mentale. 126, 74–79.
Suri, D., Schilling, R., Lopes, A.R., Mullerova, I., Colucci, G., Williams, R. and Naoumov, N.V. (2001) Non-cytolytic inhibition of hepatitis B virus replication in human hepatocytes. Journal of hepatology 35, 790-797.
Twist EM, Clark HF, Aden DP, Knowles BB and Plotkin SA. (1981) Integration pattern of hepatitis B virus DNA sequences in human hepatoma celllines. Journal of
virology 37, 239-241.
Wang, S.Y., Hsu, M.L., Hsu, H.C., Tzeng, C.H., Lee, S.S., Shiao, M.S. and Ho, C.K. (1997) The anti-tumor effect of Ganoderma lucidum is mediated by cytokines released from activated macrophages Tlymphocytes. International journal of cancer 70, 699–705.
Yang, H.I., Lu, S.N., Liaw, Y.F., You, S.L., Sun, C.A., Wang, L.Y., Hsiao, C.K., Chen, P.J., Chen, D.S. and Chen, C.J., (2002) Taiwan community-based cancer screening project group Hepatitis B e antigen and the risk of hepatocellular carcinoma. The
New England journal of medicine 347, 168-174.
Van Zonneveld M, Honkoop P and Hansen BE. (2004) Long-term followup of alpha-interferon treatment of patients with chronic hepatitis B. Hepatology 39(3),
804-810.
Figure 1. The relative HBsAg expression in Hep3B treated with PHA-MNC-CM from the CMT and SC groups. a. SC-PHA-MNC-CM, conditioned media prepared from sedentary control blood mononuclear cells that were stimulated by phytohemagglutinin and b. CMT-PHA-MNC-CM, conditioned media prepared from cyclist blood mononuclear cells that were stimulated by phytohemagglutinin. The concentrations of PHA (μg/ml) were the final concentrations during incubation of blood mononuclear cells.* Significant difference (p < 0.01).
Figure 2. Levels of secreted IFN-γ in PHA-MNC-CM. Conditioned media prepared from blood mononuclear cells that were stimulated by phytohemagglutinin (PHA-MNC-CM). CMT, (cyclist group); SC, (sedentary control).The concentrations of phytohemagglutinin (PHA, μg/ml) were the final concentrations when incubated with blood mononuclear cells. *Significant difference (p < 0.01).
Figure 3. Level of secreted TNF-α in the PHA-MNC-CM. The conditioned media prepared from blood mononuclear cells that were stimulated by phytohemagglutinin (PHA-MNC-CM). CMT, (cyclist group); SC, (sedentary control). The concentrations of phytohemagglutinin (PHA, μg/ml) were the final concentrations when incubated with MNC. *Significant difference (p < 0.01).
Figure 4. Levels of secreted IFN-α in PHA-MNC-CM. Conditioned media prepared from blood mononuclear cells that were stimulated by phytohemagglutinin (PHA-MNC-CM). CMT, (cyclist group); SC, (sedentary control). The concentrations of PHA (μg/ml) were the final concentrations when incubated with MNC. *Significant difference (p < 0.01).
Figure 5. Levels of secreted IL-1β in PHA-MNC-CM. Conditioned media prepared from blood mononuclear cells that were stimulated by phytohemagglutinin (PHA-MNC-CM). CMT, (cyclist group); SC, (sedentary control). The concentrations of PHA (μg/ml) were the final concentrations when incubated with MNC.* Significant difference (p < 0.01).
Table 1. Basic physiologic characteristics and cardiopulmonary fitness of subject
The cyclist athletes with moderate training group were designated as (CMT) and the sedentary control group as (SC). Results are expressed as mean ± SEM.
SC CMT
Subjectnumber 14 12
Age (year) 23.2 ± 2.9 22.9 ± 2.3
Height (cm) 172 ± 3 173 ± 3
Body mass (kg) 64.6 ± 3.6 65.9 ± 4.1
Resting heart rate
(beats/min) 71.3 ± 4.2 50.2 ± 2.6
Max. heart rate
(beats/min) 187.5 ± 3.2 182.4 ± 3.0
V
‧O2max(ml/[kg x min]) 38.7 ± 4.5 59.3 ±2.6 Albumin (g/dl)
Key points
• Competitive cyclists with moderate training possessed greater immunomodulatory response against surface antigen expression of human hepatitis B virus than did sedentary controls.
• The cytokines such as IFN-γ, TNF-α, IFN-α and IL-1β in the MNC-CM from the
cyclists were higher than those in the control group.