行政院國家科學委員會專題研究計畫 成果報告
探討 haloperidol 抑制單核球與小神經膠細胞誘發基質金屬 蛋白酵素活化之作用機轉及其對大鼠蜘蛛網膜下腔出血後
之保護作用評估
計畫類別: 個別型計畫
計畫編號: NSC94-2320-B-038-050-
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 臺北醫學大學藥理學科
計畫主持人: 蕭哲志
計畫參與人員: 林彥妤 周泳臣
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 95 年 10 月 31 日
國科會精簡報告
計畫名稱:
探討 Haloperidol 抑制單核球與小神經膠細胞誘發基質金屬蛋白酵 素活化的作用機轉及其對大鼠蜘蛛網膜下腔出血後之保護作用
評估
計畫類別:個別型計畫
計畫編號: 94-2320-B-038-050 執行期間:94.08.01-95.07.31 計畫主持人:
蕭哲志 副教授
執行單位:
臺北醫學大學藥理學研究所
中 華 民 國 九十五 年 十 月 二十 日
Chinese abstract 中文摘要
基質金屬蛋白酵素(matrix metalloproteinases,MMPs)具有可以分解 及破壞細胞外基質(extracellular matrix,ECM)之能力。因此 MMPs 對於 組織的重組(remodeling)、修補(repairing)與破壞(destroy)上扮演著相當重 要之角色。而最近研究證據顯示出在一些神經退化性疾病的病理過程 中,例如:阿滋海默症(Alzheimer's disease)以及多發性硬化症(multiple sclerosis)等,皆與 MMPs 有關。另一方面,在癌症中,MMPs 也參與著 癌細胞生長及前進的許多過程且能使癌細胞在轉移(metastasis)入侵的進 程中穿透周圍組織。
在臨床上,用來治療精神疾病方面的藥物haloperidol 發現具有抗發 炎之效果。所以假設此藥物haloperidol 能抑制發炎時所誘發的不正常組 織重組(remodeling)。因此本研究使用可以誘發 MMP-9 大量表現之腫瘤 壞 死 因 子 (tumor necrosis factor-α , TNF-α) 與 細 菌 類 毒 素 (lipopolysaccharide,LPS)作為 THP-1 細胞之刺激劑。由電泳酵素分析法 (gelatin zymography)下,我們觀察到由 TNF-α刺激 MMP-9 酵素之活化及 表現量會隨著haloperidol 濃度增加皆可有效地被抑制。接著利用細胞存 活率測定(MTT assay)可發現 haloperidol 的抑制作用並非源自細胞之損 壞或死亡。以西方點墨法(Western blot)之實驗方法,發現 haloperidol 濃
度增加可有效地抑制由 TNF-α刺激 MMP-9 蛋白之表現量。此外在
enzyme-linked immunosorbent assay (ELISA)實驗中發現,haloperidol 於較
高濃度 20 μM 時能有意義地降低 TIMP-1 之產生。另外還發現,
haloperidol 可以隨著濃度增加而抑制 LPS 所誘發的 MMP-9 酵素活化與
haloperidol 能抑制 MMP-9 之 mRNA 表現。綜合上述實驗結果顯示 haloperidol 抑制效果較 valproic acid 強(haloperidol IC50:5.4 ± 0.3 μM;
valproic acid IC50:255.8 ± 73.8 μM)。
藉由Western blot 之實驗發現 haloperidol 能顯著地抑制由 TNF-α或 LPS 刺激所導致的 total inhibitor-κBα (IκBα)之降解作用。接著利用電泳 移動偏向分析法(electromobility shift assay,EMSA),分析細胞核內外 NF-κB (p65)之轉位與活化情形,得知 haloperidol 濃度為 10 μM 時,可 有意義地抑制TNF-α所誘發的 NF-κB 之轉位與活化。在 mitogen-activated protein kinases (MAPKs)方面,由目前結果觀察到 haliperidol 在較高濃度 (20 μM)時抑制 TNF-α所誘導的 c-Jun-NH2-terminal kinase (JNK)活化情 形較為明顯。另外更進一步發現 haloperidol 並不會影響 LPS 在 THP-1 cells 中所誘發之內生性 TNF-α表現量。
綜合目前之實驗結果,發現haloperidol 確實能選擇性地抑制人類單 核球細胞(THP-1 cells)中,TNF-α所誘發的 MMP-9 活性與表現,而此抑
制之機轉可能主要是影響NF-κB 之訊息傳遞路徑。期許未來能更加暸解
在活體實驗中對於抗發炎反應的功能與療效。
English Abstract
Matrix metalloproteinases (MMPs) play an important role in matrix remodeling, repairing and destroying. Recent evidence has indicated that MMPs are involved in the pathogenesis of neurodegenerative diseases as Alzheimer's disease and multiple sclerosis. On the other hand, in cancer, MMPs encourage tumor cells to penetrate the surrounding tissue during the invasive process of metastasis and can mediate many processes in tumor growth and progression.
Haloperidol have been used to treat psychotic diseases clinically. These two drugs have been shown to exert anti-inflammatory effects. Therefore, we hypothesized that haloperidol inhibited the inflammation-induced abnormal remodeling. We used tumor necrosis factor-α (TNF-α) and lipopolysaccharide (LPS) as a stimulus, that induced expression of MMP-9 in human monocytic THP-1 cells. According to gelatin zymography method, we found that the expression and activation of MMP-9 protein induced by TNF-α was inhibited by haloperidol in a concentration-dependent way. We also found that the inhibitory effect of haloperidol was not due to impairment of cellular viability measured by MTT assays. According to Western blot analysis, we observed that the inhibition on TNF-α-induced expression of MMP-9 protein by haloperidol and valproic acid is concentration-dependent.
Furthermore, we found at higher concentration of haloperidol at 20 μM could significantly reduced TIMP-1 proteins measured by enzyme-linked
immunosorbent assay (ELISA). Besides, we even more found that haloperidol suppressed MMP-9 activation and the expression of MMP-9 protein induced by LPS in a dose dependent way. In transcription level, haloperidol also suppressed the TNF-α-induced MMP-9 mRNA expression measured by RT-PCR. According to the previous experiments, we observed that haloperidol was more potent than valproic acid (haloperidol IC50:5.4 ± 0.3 μM;valproic acid IC50:255.8 ± 73.8 μM).
We found that haloperidol significantly inhibited the degradation of total inhibitor-κB-α (IκBα) induced by TNF-α or LPS in Western blot analysis.
In the nuclear aspect, we found that at 10 μM significantly inhibited TNF-α-induced NF-κB translocation and activation measured by EMSA. In MAPKs aspect, the concentration of haloperidol at 20 μM significantly suppressed JNK expression in TNF-α stimulation. Furthermore, we found haloperidol did not influence the LPS-induced production of TNF-α in the medium of THP-1 cells.
In summary, we found that haloperidol had inhibitory effect on MMP-9 expression and activation in THP-1 cells. Its main mechanism of action might be through NF-κB signal pathway on TNF-α and LPS stimulation. It will be interesting to do further studies and investigation on haloperidol as therapeutic targets on inflammatory research in vivo.
Introduction
基質金屬蛋白酵素matrix metalloproteinases
,
MMPs)可由多種細胞類 型(cell types)分泌,其結構中含有鋅(zinc)離子,屬於 endopeptidases 家 族,於生理上,具有分解細胞外基質(extracellular matrix, ECM)之大分子 能 力 , 與 組 織 的 型 態 改 變(morphogenesis) 、 修 補 (repair) 、 血 管 新 生 (angiogenesis)有關。而在病理角色方面,當 MMPs 過度分解 ECM 時,會引起一些疾病產生,例如:風濕性關節炎(rheumatoid arthritis)、骨關 節炎(osteoarthritis)、牙周炎(periodontitis)、皮膚自體免疫失調及腫瘤的 侵入與轉移等(Nagase et al., 1999; Shapiro et al., 1998; Westermarck et al., 1999)。根據 MMPs 之 domain 結構、對受質(substrate)的專一性,可分為 六大類(Table 1),且它們仍然保有部分共有的特徵:(1)能催化水解蛋白 質或ECM 中之主成份(proteoglycan);(2)結構中的 catalytic domain 上擁 有 鋅(zinc) 離 子 及 鈣 離 子 ; (3) 剛 分 泌 出 來 的 MMPs 不 具 有 活 性 (proMMPs) ; (4) 可 被 人 體 內 生 性 之 tissue inhibitor of matrix metalloproteinases (TIMPs)所抑制(Van Wart et al., 1990)。
MMPs 在 in vitro 未接受刺激的細胞培養下和 in vivo 完整的組織中通
常表現量都是很低的;在不同的生理環境下,當體內的 ECM 需要重新
塑造(remodeling)時,例如:胚胎形成(embryogenesis)、傷口修復(repair) 和 骨 骼 重 塑 (remidoling) , 或 受 到 許 多 種 類 的 cytokines ( 如 : TNF-α, IL-6)、growth fators (如:EGF, bFGF, IL-1, PDGF 與 TGF-β)、物 理 性 壓 力(stress) 、 chemical agents ( 如 : phorbol esters) 、 荷 爾 蒙 (hormones)、細胞與基質和細胞與細胞之間的交互作用刺激下,均會誘 導活化MMPs 大量表現(Westermarck and Kahari, 1999),所以 MMPs 的
表現必須被嚴密的調控著。而MMPs 家族是屬於可被高度調節的蛋白且 它們的調節至少包括三種不同方式:(1)MMPs 轉錄(transcription)之調 節;(2)proMMPs 之活化;(3)MMPs 之抑制(Curran et al., 2000; Stemlicht et al., 1999)。
有 許 多 不 同 的 訊 息 傳 遞 路 徑 可 以 透 過MMP 的 transcriptional activators,以影響MMPs蛋白的表現,例如:Mitogen-activated protein kinases (MAPKs)路徑。在不同的細胞類型中,可藉由這類路徑來刺激或 抑制MMPs的表現(Westermark et al., 1999; Simon et al., 1998; Johansson et al., 2000)。在特定MMPs (如:MMP-1、MMP-2、MMP-3、MMP-7、
MMP-9、MMP-11、MMP-13、MMP-14和MMP-19)基因上的promotor regions中之binding sites調控片段已被定序出來(Figure 3)。藉由許多細胞 外 的 刺 激 可 使MMP基因啟動子(promoter)與其轉錄因子(transcription factors)結合而調控MMPs的表現。在惡質性的癌症(malignant tumors)中 AP-1(是由c-fos和c-jun 所組合而成的dimer)可以正向調控MMPs的基因 表現。而其中activator protein-1 (AP-1)為MMP-1, 3, 9基因的重要轉錄因 子 。 另 外 還 有 一 些 重 要 的 轉 錄 因 子 , 例 如 :ETS會 與promoter上的 polyomavirus enhancer-A binding-protein-3 (PEA3)之位置結合(Fini et al., 1998);而Nuclear factor of κB (NF-κB)會誘導MMP-1, 3, 9, 13, 14產生 (Bond et al., 1999; Han et al., 2001) ; T-cell factor 4 (TCF4) 和 CAS-associated zinc-finger protein (CIZ) 會 活 化 MMP-1, 3, 7 的 表 現 (Crawford et al., 2001; Nakamoto et al., 2000);p53可調控MMP-1, -2, -13 的轉錄等(Sun et al., 1999, 2000)。以上是屬於正向調控的轉錄因子,但也 有負向調控因子,其Negative regulatory elements如transforming growth factor-β inhibitory element (TIE)或AG-rich element (AGRE) (Kerr et al.,
1990; Benderdour et al., 2002)。
體內的 MMPs 活性必須被內生性抑制成份嚴密的調控著,而 tissue inhibitors of metalloproteinases (TIMPs)是一種存在於大部份組織與體液 中 之 內 生 性 蛋 白 質 且 因 不 同 組 織 而 分 佈 不 同 , 並 能 專 一 且 可 逆 的 (reversible)與 MMPs 形成 stoichiometric 方式(1:1)之複合物來抑制 MMPs 酵素活性(Brew et al., 2000),所以 TIMPs 也能參與 ECM 的組織重塑 (remodeling)。TIMPs 家族可分為四個成員:TIMP-1、-2、-3 和-4 (Brew et al., 2000) , 染 色 體 之 位 置 分 別 位 於 : XP11.23-11.4 (TIMP-1) ; 11q23-11q25 (TIMP-2);22q12.1-22q13.2 (TIMP-3);3q25 (TIMP-4),而 分子量約介於21-34 kDa,且各自擁有 12 個 cysteine residues 形成 6 個雙 硫鍵(disulfide bounds)並折疊成兩個 domain (N-, C-terminal domain) (Lambert et al., 2004)。每個會依照自己不同的能力而抑制不同的 MMPs (Edwards et al., 2001)。然而 TIMPs 也能優先地抑制不同種的 proMMPs 表現,例如:TIMP-1 可以與 proMMP-9 形成複合物(Wilhelm et al., 1989),
且 TIMP-2 和-4 可以降低 proMMP-2 被 MT1-MMP 活化(Gomez et al., 1997)。另外,TIMP-3 可同時與 proMMP-2 和 proMMP-9 結合(Butler et al., 1999),而 TIMP-4 也可以結合在 proMMP-2 的 C-terminal domain 上(Bigg et al., 1997)。此外,TIMP-1 可在許多的培養細胞中表現,如:纖維母細 胞(fibroblasts) 、 上 皮 (epithelial) 和 內 皮 (endothelial) 細 胞 、 成 骨 細 胞 (osteoblast) 、 軟 骨 細 胞 (chondrocytes) 、 平 滑 肌 細 胞 和 許 多 腫 瘤 細 胞 (Welgus et al., 1985; Bord et al., 1999; Cawston et al., 1986)皆可發現其蹤 跡。而特別的是,若病人有血液方面的異常疾病時,在其血漿中可發現 TIMP-1 數值明顯增高的現象,但在生理上的重要含義並未建立清楚 (Murate et al., 1997)。其它一些外在的刺激,如:生長因子(bFGF、PDGF
和EGF、TGF-β)、phorbol esters、serum 和 cytokines (IL-6、IL-1 和 IL-1β) 皆能誘發TIMP-1 之表現。此外,TIMP-1 具有其它功能性,例如:在正 常細胞中可以促進生長,尤其在一些病理疾病中,如:纖維硬皮病 (scleroderma fibroblasts)、肝癌(hepatoma)、乳癌(breast carcinomas)、骨癌 也能促進細胞生長(Kikuchi et al., 1997)。
神經性發炎疾病在臨床領域中,MMPs 被認為參與其中,例如:(1) 破 壞 腦 部 之 血 腦 障 壁(blood-brain-barrier) (Mun-Bryce and Rosenberg, 1998; Rosenberg et al., 1996a; Rosenberg et al., 1994; Rosenberg et al., 1995);(2)中樞與脊髓之髓鞘脫失(demyelination)引起多發性硬化症 (multiple sclerosis,MS)之中樞性發炎現象;(3)感染症 cytokines 產生所 誘發之發炎反應;(4)使 amyloid 蛋白沉澱造成阿茲海默症(Alzheimer’s disease,AD);(5)神經死亡;(6)腦中癌細胞之入侵、轉移與血管增生;
(7)不正常分解細胞外基質造成結構改變等(Yong et al., 1998)。
多發性硬化症(multiple sclerosis,MS)是一種發生於中樞神經系統(腦 部以及脊髓)的疾病。而神經纖維之外層則包裹著一層稱為“髓鞘"
(myelin)的物質,像電線的塑膠皮具有絕緣之功能,可讓不同的神經傳導 訊號不會相互干擾,同時還可以加速神經訊號之傳遞。若中樞之髓鞘被 破壞時,會引起多處塊狀髓鞘脫失現象,稱為“多發性"(mutiple),接
著組織試圖將其修復,引發局部性發炎反應,結果產生 plaque 而硬化
(sclerosis),進而阻斷神經傳導路徑。在 1992 年間,於罹患多發性硬化 症(multiple sclerosis,MS)之病患中,發現為 gelatinase B (MMP-9) (Gijbels et al., 1992)。另外,一些患有視神經炎(optoc neuritis)與其他神經發炎疾 病之患者的腦脊髓液(cerebrospinal fluid,CSF)中可檢測出 gelatinase B
(MMP-9)之存在。除了可在 MS 病患之 CSF 中發現 MMP-9 外,還可於 病患之血液中看到MMP-9 與 TIMP-1 之表現,另外在血液細胞中,也可 發現 MMP-9 與 TIMP-1 的 mRNA 表現量比一般正常人高(Özenci et al., 1999)。而 Waubant 等學者之文獻指出,利用 gadolinium-enhanced magnetic resonance imagine (MRI)此項技術,於 MS 病人中若偵測到較高濃度之 MMP-9 與較低濃度之 TIMP-1,將導致病情更加惡化(Waubant et al., 1999)。而臨床上已使用 interferon-β治療 MS 病患,其中治療機轉最近才 得知,主要是由於 interferon-β可壓抑一些發炎 cytokines,如:tumor necrosis facter-α (TNF-α)與 interleukin-1 (IL-1)所誘發之 gelatinase B 活性 表 現(Bartholomé et al., 2001);此外,於實驗性自體免疫腦脊髓炎 (Experimental Autoimmune Encephalomyelitis,EAE)之老鼠動物模式中,
模擬成人類 MS 之疾病,而其中也在老鼠的 CSF 中觀察到 gelatinase B (MMP-9)之表現量增加(Gijbels et al., 1993)。無論是在人類之 MS 或老鼠 之EAE 模式中,皆發現 gelatinase B (MMP-9)可分解人類與老鼠的 myelin basic protein (MBP) (Proost et al., 1993a; Gijbels et al., 1993),而導致中樞 神經發炎現象,由此可知 MMP-9 (gelatinase B)扮演著一個很重要的角 色。
阿茲海默症(Alzheimer’s disease,AD)視為一種神經退化性疾病 (neurodegenerative disease),臨床特徵為不可逆的認知退化,於神經病理 學上,發現神經內部有神經纖維糾結(neurofibrillary tangles,NFTs)以及 神經外部《腦皮層(cortex)與海馬迴(hippocampus)》有斑塊(plaques)產生 (Price et al., 1991),這些糾結與斑塊則會阻斷神經彼此溝通和傳遞訊息的 功能。目前並不清楚神經纖維糾結(NFTs)是如何形成,但是斑塊主要是 乙型-澱粉樣蛋白(amyloid-β,Aβ)沉澱所造成的。雖然 in vivo 並無直接
證據指出何種蛋白酶能將amyloid-β (Aβ)裂解,但是 in vitro 中,卻有許 多酵素,包括 MMPs (尤其是 MMP-9)則可將此蛋白裂解(Backstrom et al., 1996)。然而 MMPs 可在阿茲海默症患者死後的腦組織中發現其蹤跡,
尤其是 MMP-9 會出現於患者海馬迴(hippocampus)的神經元(neurons)中 (Backstrom et al., 1996)。此外,還可於阿茲海默症死後腦組織之斑塊 (plaques)與 Purkinje cells 中發現 TIMPs 的表現增加(Peress et al., 1995)。
而最近的文獻指出,於培養的microglia cells 中,發現 amyloid precursor protein (APP)之片斷會刺激 MMP-9 的表現(Gottschall and Deb, 1996)以及 促使人類單核球細胞株(THP-1 cells)釋放 MMP-9 (Chong et al., 2001),所
以可於阿茲海默症患者之血液中發現 MMP-9 表現。綜合以上之發現,
可以認為MMP-9 參與著阿茲海默症之病理過程的一項重要因素。
Haloperidol (4-[4-(p-chlorophenyl)-4-hydroxypiperidino]- 4'-fluorobutyrophenone) , 結 構 式 : C21H23ClFNO2 (Figure 1) , 是 屬 於 butyrophenone類之精神安定劑(tranquilizer),第一次使用是在1957年 (settle et al., 1993),由Dr Paul Janssen所合成出來的。之後廣泛地使用在 精神病學、產科與麻醉科上(Janssen, 1967; Kudo et al., 1999; Ichikawa et al., 1999)。而最常於臨床上用來治療精神失常(psychotic disorders)、神經 疾病(neurological diseases)與控制妥瑞症(Tourett’s syndrome,好發於兒童 的慢性神經精神異常疾病)之症狀(Kapur and Remington, 2001),一般治療 的血中濃度為5~15 ng/ml (Ulrich et al., 1998)。其藥理作用機制主要是阻 斷中樞神經系統內的多巴胺(dopamine)訊息傳遞路徑(Seeman and Van Tol, 1994),為多巴胺接受體拮抗劑(antagonist),它可以部份選擇性地作 用於腦中海馬迴(hippocampus)的dopamine D2受體上,使多巴胺系統不會 過度活化(Sears et al., 1997)。另外,有文獻指出,haloperidol可以直接阻
斷 許 多 種 類 的 離 子 通 道 , 包 括 :G-protein-activated inwardly rectify potassium channels (Kobayashi et al., 2000)、calcium-activated potassium channels (Akamine et al., 2002)、HERG 與HEAG potassium channels (Shuba et al., 2001; Gessner and Heinemann, 2003)、L-, N-,P-type calcium channels (Galizzi et al., 1986; Sah and Bean, 1994)。而臨床上若長期服用 haloperidol可能會造成副作用有:因為阻斷多巴胺受體而產生錐體外副 作用(extrapyramidal syndrom,EPS),主要症狀為為急性運動不能(acute dystonia)、假性帕金森氏症(psedo-parkinsonism)、靜坐不能(akathisia)、
遲發性運動不能(tardive dyskinesia);另外,靜脈注射持續點滴給予 haloperidol 可 能 會 造 成 多 發 性 心 室 心 率 不 整 (multiforme ventricular arrhythmias)之症狀(Maldonado, 2000)。此外,Moots et al學者們認為在in vitro 中 , haloperidol 可 以 抑 制 由 LPS 刺 激 所 誘 發 的 proinflammation
cytokines,如:TNF-α與IL-β (Moots et al., 1999)。另有許多文獻指出,
無 論 是 在 in vitro 或 in vivo 實 驗 中 , 於 人 類 及 嚙 齒 類 動 物 身 上 給 予 haloperidol皆可觀察到haloperidol對於神經細胞具有細胞毒性(Petzer et al.
2000; Eyles et al. 1996; Usuki et al. 1996; Castagnoli et al. 1999;
Subramanyam et al. 1991; Van der Schyf et al. 1994; Van der Schyf et al.
1996; Avent et al. 1997; Lockhart et al. 1995; DeCoster et al. 1995),而於這 篇文獻中則指出,haloperidol對於神經細胞具有毒性可能是經由p38及 JNK路徑,以誘發神經凋亡(Noh et al., 2000)。
Nuclear factor-κB (NF-κB) 是 屬 於 人 體 內 一 個 重 要 轉 錄 因 子 (transcription factor)。而NF-κB這條路徑之活化與免疫作用(immunity) 、 細胞增生(proliferation)、凋亡(apoptosis)與發炎過程均相關。當NF-κB訊 息 受 到 影 響 , 可 能 會 導 致 某 些 疾 病 , 如 : 多 發 性 硬 化 症(multiple
sclerosis) 、 風 濕 性 關 節 炎 (rheumatoid arthritis) 、 發 炎 性 腸 道 疾 病 (inflammatory bowel disease)與氣喘(asthma)等息息相關。目前發現哺乳動 物之NF-κB家族共有五個成員:RelA (p65)、RelB、c-Rel、p105/p50 (NF-κB1)、p100/p52 (NF-κB2) (Verma et al., 1995; Ghosh et al., 1998),在 細胞未受到刺激時,是以homo-或heterodimers型式與細胞質之內生性抑 制蛋白inhibitors of NF-κB (IκBs)家族結合在一起,形成不活化狀態之 NF-κB 。 而 NF-κB 最 常 見 且 典 型 的 活 化 態 是 由 p50 與 p65 組 成 的 heterodimer (Li et al., 2002);其中,NF-κB家族皆含有一段由300個胺基 酸組成的Rel homology domain (RHD)區域,此區域可與IκBs、DNA結合 而發生反應。而IκB家族成員,主要包括IκBα、IκBβ、IκBε、BCL-3,
其中以IκBα為最主要的抑制蛋白;而平時細胞未受刺激時,NF-κB-IκB complex可以來回穿梭於細胞核與細胞質之間,如果大多數細胞處於休止 期時,則NF-κB-IκB complex大部分還是留在細胞質內(Ghosh et al., 2002);若細胞受到病原體(pathogens)《如:lipopolysaccharide (LPS)、
peptidylglycans 、 lipoproteins 、 unmethylated bacterial DNA 與 double-stranded RNA》(Imler et al., 2000)、stress signal或pro-inflammatory cytokines《如:tumor-necrosis factor (TNF)與interleukin-1 (IL-1)》刺激時,
IκB protein會被IκB kinase (IKK,由兩個catalytic subunits IKKα與IKKβ 再 加 上 一 個regulatory subunit IKKγ 所 組 成 ) 磷 酸 化 , 緊 接 著 被 ubiquitylated,隨後再被proteasome水解,使得NF-κB protein translocate 至細胞核內,與DNA上的κB sequences binding,調節一大群基因之轉錄 (Baldwin, Jr, 1996; Karin, 1999) , 包 括 : antimicrobial peptides 、 stress-response proteins 、 anti-apoptotic proteins 、 pro-inflammatory cytokines、chemokines、adhesion molecules、matrix metalloproteinases
(MMPs) 、 cyclooxygenase 2 (COX-2) 與 inducible nitric oxide synthase (iNOS)等(Baeuerle et al., 1997; Tak et al., 2001)。
Mitogen-activated protein kinases (MAPKs) 屬 於 serine/threonine kinases 酵素,會受到生長因素或環境因素等影響。主要的mammalian MAPKs為(a) extracellular signal-regulated kinases (ERK1與ERK2);(b) c-Jun N-terminal kinase (JNK1, JNK2 與 JNK3) ; (c) p38-MAPK (p38-MAPKα、p38-MAPKβ、p38-MAPKγ和p38-MAPKδ) (Figure 13)。而 其中ERK pathway主要會受到生長因子的活化且與細胞的生長、增生與 分化有關(Robinson and Cobb, 1997),另外JNK與p38 MAPK也已知為 stress-activated protein kinases (SAPK),因此它們容易受到壓力的刺激 (如:osmotic shock)、UV光的照射與一些細胞激素(如:TNF-α)的影響而 使其活化,與細胞的存活、凋亡、transformation、進化、遷移與免疫之 活化有關(Tibble and Woodgett, 1999;Ivanov and Ronai, 2000)。以上種種 的刺激因子易造成一種或多種MAPK kinase (MKK)刺激Thr與Tyr的區域 磷酸化而產生活性,進而translocate至細胞核,促使轉錄因子(AP-1為主) 磷酸化,進而啟動活化與基因表現(其中包括調節MMP-1, 3, 9等基因表 現) (Roger et al., 1994; Baud and Karin, 2001; Yang et al., 2003)。而 Activator protein-1簡稱AP-1,也是屬於轉錄因子(transcription factors)其 中ㄧ個重要成員,其組成包括ATF (ATF2、LRF1/ATF3、B-ATF、JDP1 與JDP2)、Fos (c-Fos、FosB、Fra-1與Fra-2)、Jun (c-Jun、JunB與JunD) 與Maf (c-maf、MafA、MafB、MafG/F/K與Nrl),這些蛋白會調控細胞之 增生與凋亡(Peng, 2004)。在轉錄(transcription)之過程中,AP-1的活化會 受到MAPKs調控,尤其是JNK與p38都會使c-Jun磷酸化而產生活性,造 成許多特定基因表現,如MMP-9與細胞激素的產生(Yang et al., 2003)。
無論是在中樞神經發炎疾病或是白血病發展進程上,MMPs (尤其是 MMP-9)於這些病程中都扮演著舉足輕重的角色。如 cytokines (如:
TNF-α)與 endotoxin (如:LPS)皆會刺激或誘導 MMPs 產生,而造成細胞
外基質之分解作用,但過度的MMPs 產生時,便會造成各種不同的生理
與病理傷害。故本篇論文則是以人類單核球(THP-1)為實驗細胞,藉以了 解一些中樞性藥物haloperidol 對 TNF-α與 LPS 刺激所誘發的 MMP-9 活 性及表現之影響性,並探討其中可能所牽涉的藥理作用機轉。且更進一 步地研究haloperidol 在細胞訊息傳遞路徑中,如:NF-κB 或 MAPKs 等 機制之影響性。藉以評估將來haloperidol 在癌症及發炎疾病上是否能成 為老藥新用的明日之星。
Results
一、探討 haloperidol 對 TNF-α 人類單核球細胞(THP-1 cells) 產生 MMP-9 酵素活性的作用
由之前實驗室已建立刺激物誘發MMP-9 酵素活性表現與電泳酵素分
析法(gelatin zymography)的實驗標準條件,在細胞濃度為 1 × 106 cells/ml 之條件下,投與發炎性細胞激素TNF-α (10 ng/ml)處理 THP-1 細胞 24 小 時後,利用電泳酵素分析法(gelatin zymography)可發現能誘發單核球細 胞(THP-1 cells)大量表現 MMP-9 的活性,而非 MMP-2 的活性。接著,
便以此條件為標準進行以下之實驗。
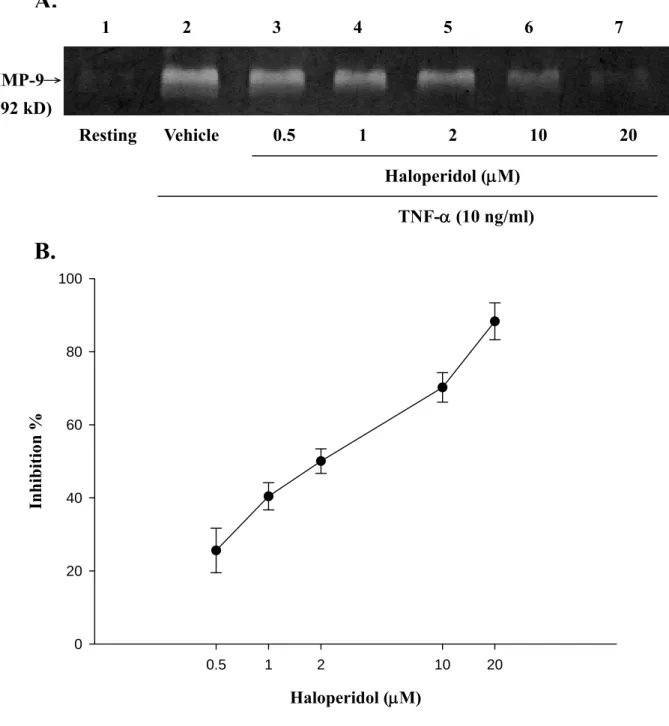
1. Haloperidol 能抑制 TNF-α刺激 THP-1 細胞產生 MMP-9 之酵素活性
本實驗首先是要探討以不同濃度的 haloperidol (0.5-20 μM)來觀察 THP-1 細胞對於利用 TNF-α 刺激而產生 MMP-9 活性之影響程度。由電 泳酵素分析法之實驗結果發現,隨著haloperidol 濃度的增加(0.5 μM、1 μM、2 μM、10 μM、20 μM ),MMP-9 的活性表現呈現逐漸減少且有意 義地被抑制之現象(Figure 2 A)。其中 haloperidol 對 TNF-α 刺激作用的抑 制百分率(Inhibition %)分別為 25.6 ± 6.1 % (0.5 μM);40.4 ± 3.7 % (1 μM);50.0 ± 3.4 % (2 μM);70.2 ± 4.0 % (10 μM)及 88.3 ± 5.1%
(20 μM),其抑制 50 %反應濃度(IC50)為 5.4 ± 0.3 μM (n = 4-7,Figure 2 B)。由此結果可知,在 THP-1 細胞中利用 TNF-α 所誘發之 MMP-9 皆能 被haloperidol 隨著濃度的上升而呈現活性減少之反應機制,並且呈現濃 度抑制效應 (concentration-dependent inhibition)。
為了證明haloperidol 藥物對於 THP-1 細胞中抑制 MMP-9 活性的表
現,並不是經由細胞毒性引起之細胞減少或死亡而導致MMP-9 的表現
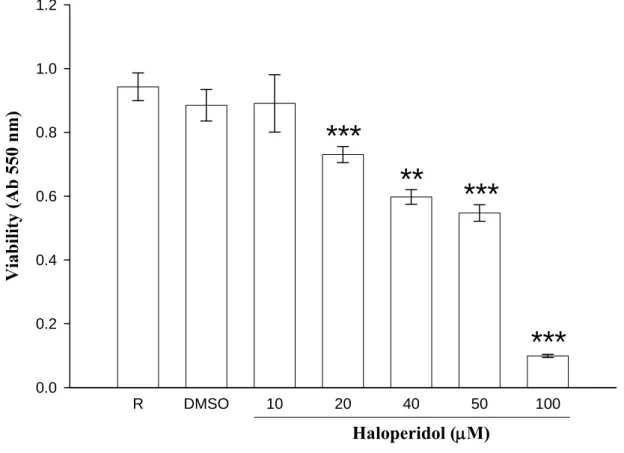
與活性減少,本實驗利用MMT assay 來測其細胞之存活率。將 THP-1 細胞(1×106 cells/ml)種植於 24 well 的培養盤中,分別投與 haloperidol (10 μM、20 μM、40 μM、50 μM 及 100 μM)處理細胞 22 小時,隨後再加入 MTT 試劑處理 2 小時,利用存活細胞的粒線體內酵素將 MTT 試劑還原 成formazan 紫色結晶,再加入 DMSO 溶解紫色結晶後,以 550 nm 吸光 值測之,此值即為存活細胞的含量。由實驗結果發現不加藥及刺激劑 時,resting 的吸光值為 0.94 ± 0.04 nm,當處理不同濃度的 haloperidol 對於細胞的數量發現有稍微的影響(10 μM,0.89 ± 0.05 nm 及 20 μM,0.73
± 0.03 nm),但整體而言 haloperidol 濃度 20 μM 以下(data not shown)並不 會明顯影響細胞的存活(n = 3~8,Figure 3),隨著 haloperidol 濃度增加,
其導致細胞死亡的程度越明顯(40 μM,0.60 ± 0.02 nm;50 μM,0.55 ± 0.03 nm 及 100 μM,0.10 ± 0.01 nm)。因此,本實驗所使用之 haloperidol 藥
物濃度範圍內對於THP-1 細胞皆不具明顯嚴重的細胞毒性。
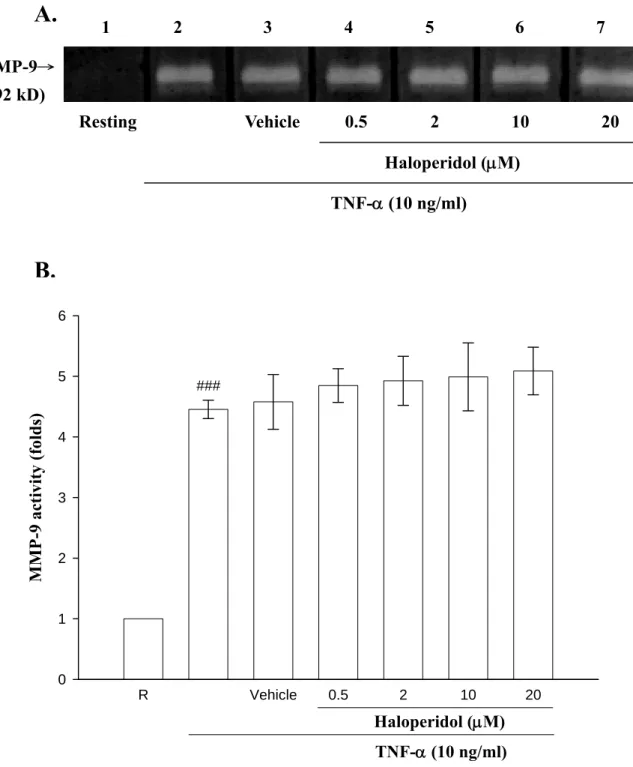
三、探討藥物 haloperidol 對於 MMP-9 酵素活性的影響
由之前的實驗結果得知haloperidol (0.5~20 μM)會呈現藥物濃度的 抑制MMP-9 活性的表現,因此,我們再利用 zymography 方法來瞭解 haloperidol 是否會直接影響 MMP-9 本身的酵素活性,由實驗結果得知 (Figure 4),初步看起來 haloperidol 對於 TNF-α (lane 2:4.45 ± 0.15 fold) 刺激後所誘發的MMP-9 酵素活性並沒有影響(lane 4,0.5 μM:4.85 ± 0.28 fold;lane 5,2 μM:4.93 ± 0.40 fold;lane 6,10 μM:4.99 ± 0.56 fold;
lane 7,20 μM:5.09 ± 0.39 fold)。
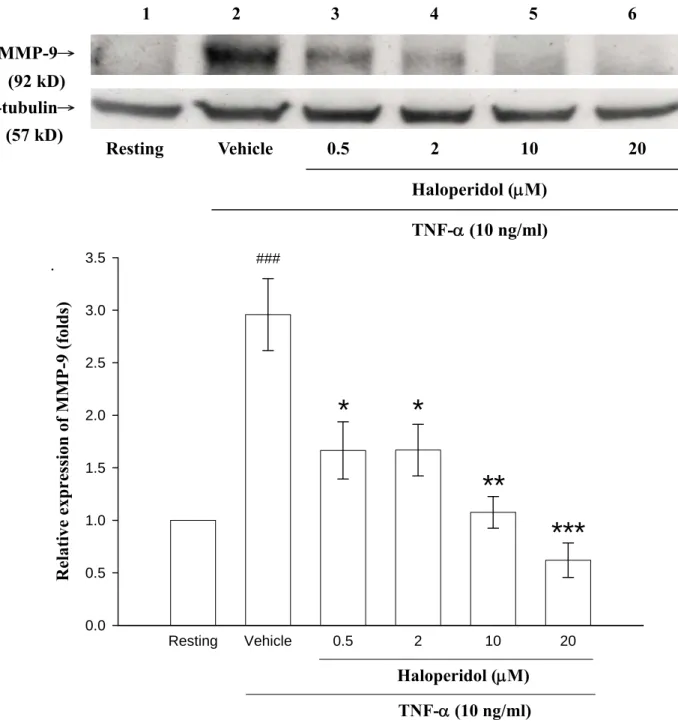
四、探討 haloperidol 對 THP-1 細胞中由 TNF-α所誘發釋放的 MMP-9 蛋白表現
由之前的實驗結果已證實TNF-α確實能誘導 THP-1 細胞中 MMP-9 的 活性增加,而haloperidol 則可抑制此作用。另外,由 MTT assay 的結果 亦可證實 haloperidol 能壓抑 MMP-9 的活性並非是由於藥物本身所引發 的 細 胞 毒 性 作 用 而 導 致 MMP-9 的 含 量 減 少 。 因 此 , 為 了 更 暸 解 haloperidol 及 valproic acid 對於 MMP-9 的抑制作用是否會影響其 MMP-9 的 蛋 白 質 含 量 表 現 , 故 我 們 利 用 西 方 點 墨 法(Western blot) 來 分 析 haloperidol 對於 MMP-9 的蛋白質表現量影響為何。
1. Haloperidol 能抑制 TNF-α刺激 THP-1 細胞產生 MMP-9 蛋白表現量
在 Figure 5 中,將 THP-1 細胞處理並培養 24 小時後,取其細胞之 萃取物(cell lysate)進行實驗,我們發現在未加藥及刺激劑的情形下 (resting,land 1),其細胞萃取物只偵測到非常微量的 92 kD 之 MMP-9
蛋白表現量。而以 TNF-α刺激 24 小時後,發現其細胞萃取物則含有大
量的MMP-9 蛋白質表現(Figure 5,land 2:TNF-α,2.96 ± 0.34 fold),當 給予不同濃度的haloperidol (0.5 μM~20 μM)處理後,可發現到 TNF-α所 誘發的 MMP-9 蛋白質表現量會隨著藥物(haloperidol)濃度的增加而呈現 有意義地逐漸減少之情形(land 3:0.5 μM,1.66 ± 0.27 fold;land 4:2 μM,
1.67 ± 0.25 fold:land 5,1.08 ± 0.15:land 6,0.62 ± 0.16)。
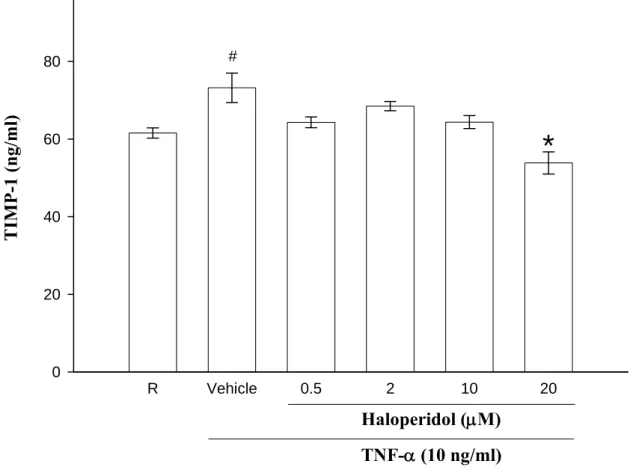
五、探討 haloperidol 對 THP-1 細胞中由 TNF-α所誘導釋放的 TIMP-1 蛋白表現量
有許多文獻指出,發炎性細胞激素如:TNF-α會影響 MMP-9 的活化
作用。在細胞中,MMPs 的含量與其活性表現均受到許多內生性蛋白嚴 密的調節與控制,其中,MMPs 的活化與否,也與內生性組織抑制劑 (tissue inhibitor of metalloproteinases,TIMPs)的生成與結合有關。目前已 知的 TIMPs 有四種,每種 TIMPs 對於 MMPs 的親和力與抑制力都不盡 相同。而TIMP-1 則是 MMP-9 的主要內生性組織抑制劑(Brew, 2000)。
因此,為了暸解 MMP-9 蛋白表現量的減少是否與其內生性的抑制劑
TIMP-1 有關,所以我們將 zymography 實驗所取得的上清液(supernatant)
進行 TIMP-1 蛋白表現的實驗。我們以酵素免疫分析測定法(Human
TIMP-1 ELISA system)來觀察,以每 106的細胞數為單位。在 Figure 6 中,
發現 THP-1 細胞在沒有外界的刺激之下(resting),便可偵測到些許程度 的TIMP-1 蛋白表現(land 1:61.55 ± 1.32 ng/ml )。經 TNF-α刺激後,則 可見到有意義的上升情形(land 2:73.19 ± 3.80 ng/ml)。之後投予不同濃 度的haloperidol 0.5 μM (land 3)、2 μM (land 4)、10 μM (land 5)和 20 μM (land 6)處理後,發現 TIMP-1 蛋白表現並無明顯的影響且無統計上之意 義(land 3;64.31 ± 1.40 ng/ml;land 4:68.48 ± 1.18 ng/ml;land 5:64.37
± 1.68 ng/ml )。但在投予濃度為 20 μM 之 haloperidol 時(land 6:53.84 ± 2.87 ng/ml),則呈現有意義的抑制情形。
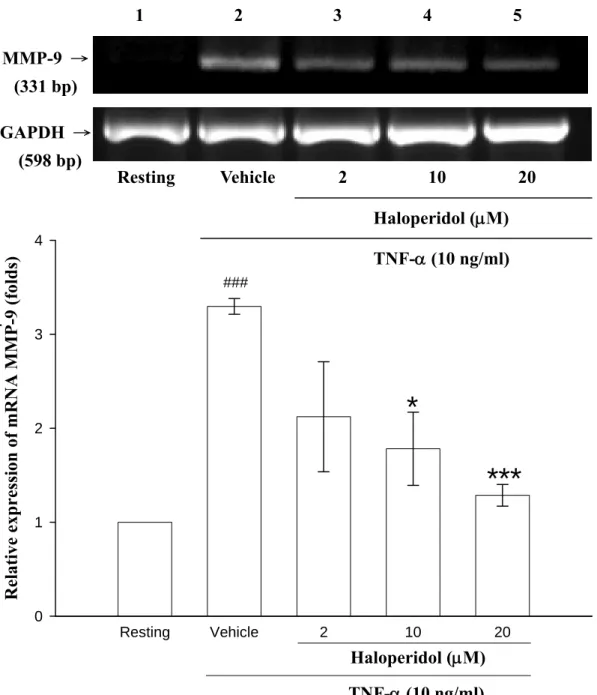
六、探討 haloperidol 對於 THP-1 細胞中以 TNF-α所誘發的 MMP-9 mRNA 之影響
由之前的實驗結果發現haloperidol 可以明顯的抑制由 TNF-α或 LPS
所誘發的MMP-9 酵素表現量,而其細胞內的蛋白質表現量也有明顯的
抑制作用。因此,我們接著觀察MMP-9 基因的轉錄過程中是否受到
haloperidol 的抑制作用。首先,先萃取 THP-1 細胞內全部的 RNA 來進
行reverse transcriptase reaction (RT-PCR)實驗,利用反轉錄酵素(reverse transcriptase)將所萃取的 total RNA 轉換成 cDNA,再於核酸循環加熱器 中進行聚合酵素連鎖反應(簡稱 PCR)。使用 MMP-9 與 GAPDH 的專一性 引子(primers)來進行放大 MMP-9 與 GAPDH 之 cDNA 產物,並以 1.5 % 的洋菜膠(agarose gel)進行電泳分析,House-keeping gene,GAPDH 在此 為 internal control。由 Figure 7 的結果顯示,以 TNF-α (10 ng/ml)處理 6 小時後,MMP-9 mRNA 的表現很明顯地被誘發出來(land 2:TNF-α,3.30
± 0.08 fold),而給予藥物 haloperidol (lane 3:2 μM,2.12 ± 0.59 fold;lane 4:10 μM,1.78 ± 0.39 fold;lane 5:20 μM,1.29 ± 0.12 fold)之後,我 們可以觀察到TNF-α所誘發 MMP-9 mRNA 的表現會隨著 haloperidol 濃 度的增高,而呈現抑制的效果。
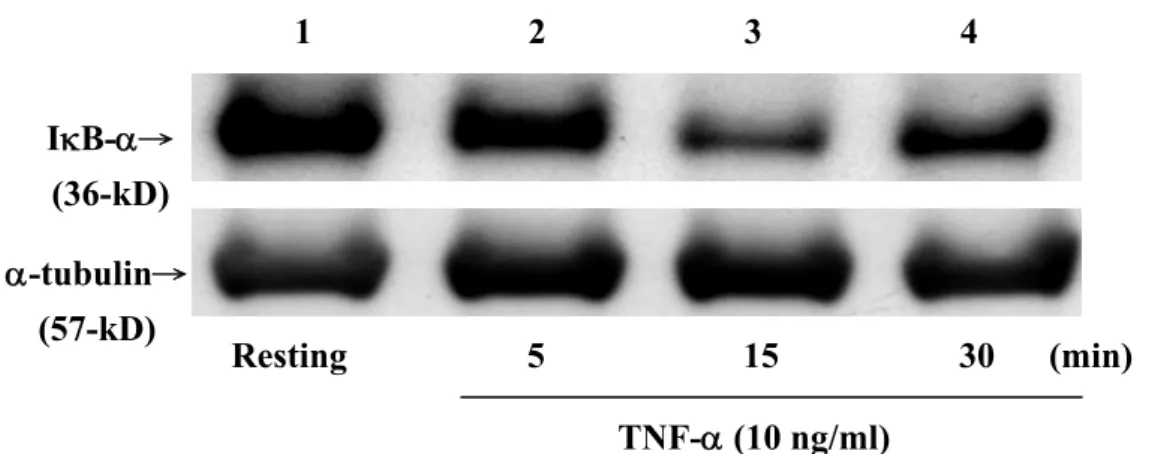
七、探討 haloperidol 對 TNF-α刺激 THP-1 細胞引發 IκB-α之 降解作用
轉錄因子(如:NF-κB/Rel family)可以調控許多發炎反應、細胞增生以 及細胞凋亡等等之基因表現。因此當細胞(如:單核球細胞)受到外來刺 激時(如:細菌內毒素 LPS),會釋放出部分的內生性調節因子,並間接 造成轉錄因子(NF-κB/Rel)的活化作用(Baldwin, 1996; O’Connell, 1998;
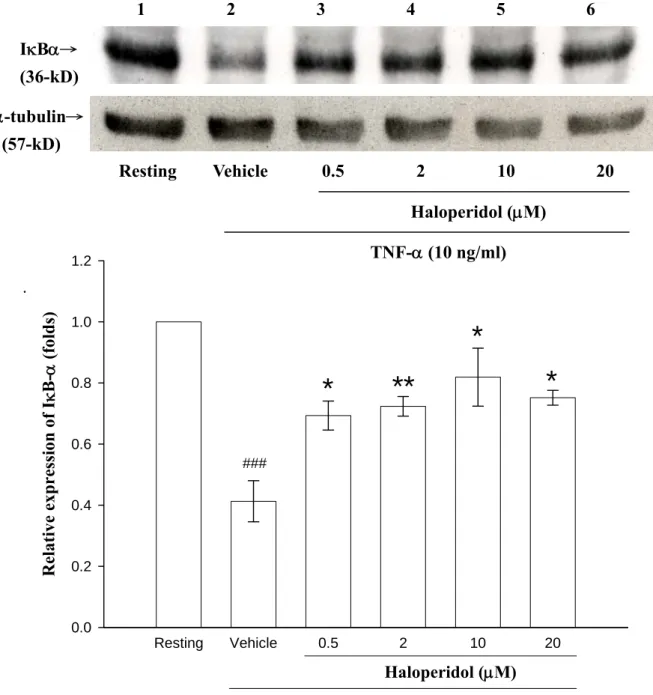
Martin, 2000)。其中,內生性調節因子最主要的就是細胞激素(如:TNF-α 和IL-1β等)。轉錄因子最主要的活化途徑是藉由 IκB-α的 degradation 使 NF-κB 進入核中使基因進行轉錄作用。因此,首先利用 Western blotting 來觀察NF-κB 的抑制因子(IκB-α)蛋白表現情形,以瞭解 haloperidol 在 IκB-α磷酸化的分解過程,並進而影響 NF-κB 的活化上所扮演的角色。
首先以TNF-α刺激 THP-1 細胞,觀察不同時間點,其 total IκB-α之蛋白
表現。由Figure 8 的結果發現,以培養 30 min 的 Resting 當 1,接著以 TNF-α (10 ng/ml)刺激 5 分鐘(land 2:0.743 fold)、15 分鐘(land 3:0.453 fold) 及30 分鐘(land 4:0.610 fold)後、發現在 THP-1 cell 以 TNF-α刺激 15 分 鐘時,total IκB-α的降解作用最為明顯,造成 IκB-α蛋白的表現明顯被減
少。因此根據這些結果,將細胞以TNF-α刺激 15 分鐘來進行之後的實
驗。
在 Figure 9 中,當未投予 TNF-α時,因無訊息之傳遞,total IκB-α則 無法進行降解作用,故 IκB-α蛋白的表現為最高(lane 1:1 ± 0.0),以 TNF-α 刺 激 15 分 鐘 (land 2 : 0.41 ± 0.07 fold) , 投 予 不 同 濃 度 之 haloperidol,可發現 IκB-α含量隨著 haloperidol 濃度的增高(land 3:0.5 μM,0.69 ± 0.05 fold;lane 4:2 μM,0.72 ± 0.03 fold;land 5:10 μM,
0.82 ± 0.09 fold)而逐漸上升。但是當藥物提高到 20 μM 時,卻發現 haloperidol 對於 IκB-α含量回升作用具有稍微降低現象(lane 6:20 mM,
0.75 ± 0.24 fold)。由以上所觀察到的結果,我們可以推論 haloperidol 可 能會藉由抑制IκB-α 蛋白的降解作用,來阻止 NF-κB 進入細胞核內與特 定序列之DNA 結合的作用而達到抑制 TNF-α刺激所誘發的 MMP-9 活性 表現。
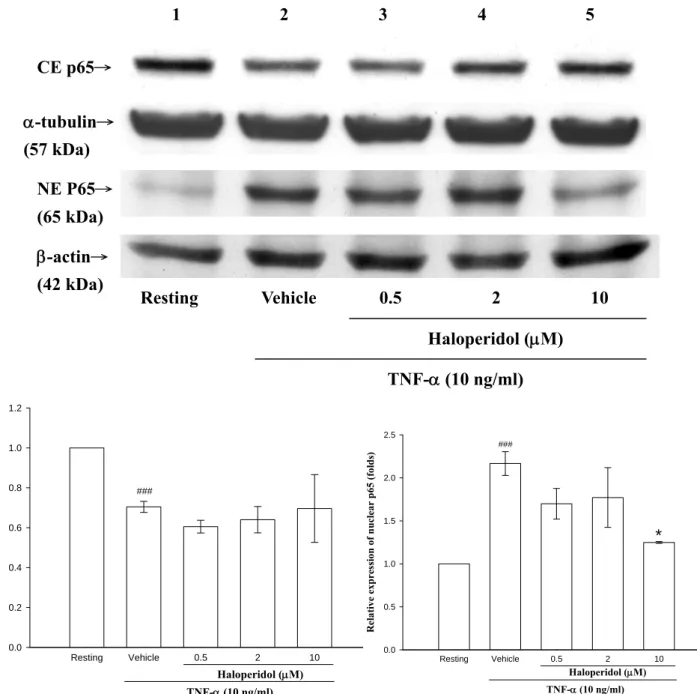
八、探討 haloperidol 對 TNF-α刺激 THP-1 細胞核內與核外 p65 蛋白質之轉位(translocate)作用
根據上述的實驗結果知道 haloperidol (0.5, 2, 10 μM)可以減少核內 NF-κB 與 DNA binding 之表現量,推測是否是因為減少 NF-κB (p65)轉 錄因子進入核內所造成的,故接下來將探討haloperidol 對於 TNF-α誘發 之 NF-κB (p65)進入細胞核內之影響。首先利用 NE-PERTM nuclear and
cytoplasmic extraction reagents 萃取細胞核內及細胞質的蛋白質,再利用 Western blot 觀察 THP-1 細胞核內外之 p65 表現情形。根據之前本實驗 室所做的結果,發現在THP-1 cell 以 TNF-α刺激 30 分鐘時,p65 的表現 最為明顯(data not shown),根據這些結果,將細胞以 TNF-α刺激 30 分鐘 來進行以後的實驗。在 Figure 10 中可以觀察到 TNF-α刺激 30 分鐘之後,
細胞質中的p65 表現量會降低(lane 2:0.70 ± 0.03 fold;n=5),細胞核內 p65 所表現的量會明顯的增加(land 2:2.17 ± 0.14 fold;n=4)。當投與不 同濃度的haloperidol (0.5, 2, 10 μM)處理後,在細胞質發現濃度為 0.5 與 2 μM 之 p65 減少情形比只投與刺激劑來得明顯(lane 3,0.5 μM:0.61 ± 0.03 fold;land 4,2 μM:0.64 ± 0.07 fold;n=4),不過濃度為 10 μM 即 開始具有p65 反轉情形(land 5,10 μM:0.7 ± 0.17 fold;n=2),但不具統 計上的意義。在細胞核中,觀察到haloperidol 在濃度為 0.5 與 2 μM 時,
無明顯變化(lane 3,0.5 μM:1.70 ± 0.18 fold;land 4,2 μM:1.77 ± 0.35 fold;n=4),但濃度為 10 μM 時則可有意義地抑制 p65 translocate 至細胞 核的情形(land 5,10 μM:1.25 ± 0.01 fold;n=2;P <0.05)。顯示 haloperidol 能抑制由TNF-α所誘發的 p65 蛋白表現。
九、探討 haloperidol 對 TNF-α刺激 THP-1 細胞核內 NF-κB 蛋 白質轉位與活化之作用
根據之前的實驗結果推測 haloperidol 較明顯可以抑制 IκB-α 蛋白的 降解作用,故我們投予haloperidol 濃度為 0.5, 2, 10 μM,利用 EMSA 之 方法來探究藥物是否確實會影響其核內NF-κB 與 DNA binding 之能力。
我們分別以 TNF-α (10 ng/ml)刺激細胞15分鐘及 30 分鐘,發現在 30 分 鐘之時間點,細胞核內NF-κB 與 DNA binding 之表現量較為明顯(data not
shown),也因此之後實驗皆以刺激 30 分鐘為時間點。我們可以發現 (Figure 11),當未給予 TNF-α (10 ng/ml)刺激時,細胞核內 NF-κB 與 DNA binding 幾乎沒有表現(lane 1)。以 TNF-α刺激 30 分鐘後,發現 DNA 明 顯產生 shift 現象(lane 2,n=5)。當投與不同濃度之 haloperidol (0.5~10 μM),可發現 shift 現象有減少的情形,而濃度為 0.5 μM 時即可有意義 地抑制 THP-1 細胞核內 TNF-α所誘發的 NF-κB 活化情形(land 3,
0.5 μM:0.74 ± 0.06 fold;n=5;P <0.01),但濃度為 2 μM 時此抑制效果 卻減少(land 4,2 μM:0.85 ± 0.07 fold;n=5),不過較高濃度 10 μM 發 現可明顯抑制NF-κB 活化(land 5,10 μM:0.32 ± 0.01;n=2;P <0.001),
顯示haloperidol 可以減低核內 NF-κB 與 DNA binding 的表現量。由以上 所觀察到的結果可以推論,haloperidol 可以減少核內 NF-κB 與 DNA binding 之表現量。
十、探討 haloperidol 對 TNF-α誘發 THP-1 細胞內 MAPKs 路 徑中 JNK 之活化作用
基因表現的訊息調控路徑,除了藉由上述 NF-κB 路徑之活化作用
外,也可以經由Mitogen-activated protein kinases (MAPKs)的訊息傳遞路 徑。由於主要的MAPKs 為 extracellular signal-regulated kinase (ERK 44/42 kD)、c-Jun N-terminal kinase (JNK 54/46 kD)和 p38 kinase。而在細胞訊 息傳遞之過程中,須自身被磷酸化才具有活化之酵素型態,進而轉移進 入細胞核,再與轉錄因子作用以進行基因表現。由於它們易受發炎性細 胞激素(cytokines)的影響而被活化,所以利用 Western blot 瞭解 TNF-α是 否經由 MAPKs 的路徑來誘發 MMP-9 的表現,並觀察 haloperidol 在其 中所扮演的角色。根據之前本實驗室所做的結果,發現在THP-1 cell 以
TNF-α刺激 15 分鐘時,JNK (46 kD)的表現為較為明顯,所以以此為實 驗條件(data not shown)。在 Figure 12 中,可以觀察到在 TNF-α刺激 15 分鐘之後,phospho-JNK (46 kD)表現有明顯的增加(lane 2,1.56 ± 0.13 fold),分別投與不同濃度的 haloperidol 之後,抑制由 TNF-α刺激 phospho-JNK (46 kD)蛋白表現的詳細數值如下:haloperidol 2 μM (land 3) 為1.39 ± 0.16 fold、10 μM (land 4)為 1.45 ± 0.13 fold,而 20 μM (land 5) 則為 1.10 ± 0.05 fold。發現當 haloperidol 濃度為 20 μM 時,抑制 phospho-JNK (46 kD)蛋白表現量較為明顯,約抑制了 80 %;另外 haloperidol 則不會影響由 TNF-α刺激之 total-JNK (46 kD)蛋白表現。因 此,TNF-α所誘導產生的 MMP-9 也可能經由 JNK 之訊息傳遞路徑。據 此實驗結果推論haloperidol 也可能會經由抑制 JNK 蛋白的表現,再進一 步的抑制轉錄因子的活化以及影響TNF-α所誘發的 MMP-9 表現量。
十一、探討 haloperidol 對於 LPS 刺激人類單核球細胞(THP-1 cells)產生 MMP-9 酵素活性的作用
由之前實驗室已建立刺激物誘發 MMP-9 酵素活性表現與電泳酵素
分析法(gelatin zymography)的實驗標準條件,在細胞濃度為 1 × 106 cells/ml 之條件下,投與細菌之內毒素 LPS (50 ng/ml)處理 THP-1 細胞 24 小時後,利用電泳酵素分析法(gelatin zymography)可發現能誘發單核球 細胞(THP-1 cells)大量表現 MMP-9 的活性,而非 MMP-2 的活性。接著,
便以此條件為標準進行以下之實驗。
■ Haloperidol 能抑制 LPS 刺激 THP-1 細胞產生 MMP-9 之酵素活性 本實驗首先是要探討以不同濃度的haloperidol (0.5~20 μM)來觀察
THP-1 細胞對於利用 LPS 刺激而產生 MMP-9 活性之影響程度。由電泳 酵素分析法之實驗結果發現,隨著haloperidol 濃度的增加(0.5 μM、2 μM、10 μM、20 μM ),MMP-9 的活性表現呈現逐漸減少且有意義地被 抑制之現象(Figure 13 A)。其中 haloperidol 對 LPS 刺激作用的抑制百分 率(Inhibition %)分別為 38.5 ± 0.9 % (0.5 μM);47.5 ± 4.2 % (2 μM);76.6
± 1.4 % (10 μM);91.6 ± 3.1 % (20 μM),其抑制 50 %反應濃度(IC50)為 3.1
± 0.8 μM (n = 3,Figure 13 B)。由此結果可知,在 THP-1 細胞中經由 LPS 所誘發之MMP-9 皆能被 haloperidol 隨著濃度的上升而呈現活性減少之 反應機制,並且呈現濃度抑制效應 (concentration-dependent inhibition)。
十二、探討 haloperidol 對 THP-1 細胞中由 LPS 所誘發釋放的 MMP-9 蛋白表現
由之前的實驗結果(Figure 30)已證實 LPS 確實能誘導 THP-1 細胞中 MMP-9 的活性增加,而 haloperidol 則可抑制此作用。因此,為了更暸 解haloperidol 對於 MMP-9 的抑制作用是否會影響其 MMP-9 的蛋白質含 量表現,故我們利用西方點墨法(Western blot)來分析 haloperidol 對於 MMP-9 的蛋白質表現量影響為何。
■ Haloperidol 能抑制 LPS 刺激 THP-1 細胞產生 MMP-9 蛋白表現量
在Figure 14 中,將 THP-1 細胞處理並培養 24 小時後,取其細胞之 萃取物(cell lysate)進行實驗,我們發現在未加藥及刺激劑的情形下 (resting,land 1),其細胞萃取物只偵測到非常微量的 92 kD 之 MMP-9
蛋白表現量。而以 LPS 刺激 24 小時後,發現其細胞萃取物則含有大量
的MMP-9 蛋白質表現(Figure 31,land 2:LPS,2.81 ± 0.22 fold),當給
予不同濃度的haloperidol (2 μM~20 μM)處理後,可發現到 LPS 所誘發的 MMP-9 蛋白質表現量會隨著藥物(haloperidol)濃度的增加而呈現有意義 地逐漸減少之情形(land 3:2 μM,1.41 ± 0.13 fold;land 4:10 μM,1.04
± 0.13 fold;land 5:20 μM,0.54 ± 0.10 fold)。
十三、探討 haloperidol 對 LPS 刺激 THP-1 細胞引發 IκB-α之 降解作用
轉錄因子(如:NF-κB/Rel family)可以調控許多發炎反應、細胞增生以 及細胞凋亡等等之基因表現。因此當細胞(如:單核球細胞)受到外來刺 激時(如:細菌內毒素 LPS),會釋放出部分的內生性調節因子,並間接 造成轉錄因子(NF-κB/Rel)的活化作用(Baldwin, 1996; O’Connell, 1998;
Martin, 2000)。其中,內生性調節因子最主要的就是細胞激素(如:TNF-α 和IL-1β等)。轉錄因子最主要的活化途徑是藉由 IκB-α的 degradation 使 NF-κB 進入核中使基因進行轉錄作用。因此,首先利用 Western blotting 來觀察NF-κB 的抑制因子(IκB-α)蛋白表現情形,來瞭解 haloperidol 在 IκB-α磷酸化的分解過程,並進而影響 NF-κB 的活化上所扮演的角色。
首先以LPS 刺激 THP-1 細胞,觀察不同時間點,其 total IκB-α之蛋白表 現。由Figure 15 的結果發現,以培養 120 min 的 Resting 當 1,接著以 LPS (50 ng/ml)刺激 30 分鐘(land 2:1.062 fold)、60 分鐘(land 3:0.918 fold)、90 分鐘(land 4:0.769 fold)及 120 分鐘(land 5:1.302 fold)後,則 以LPS 刺激 90 分鐘時,其 total IkB-α的降解作用最為明顯。因此根據 這些結果,將細胞以LPS 刺激 90 分鐘來進行之後的實驗。
■ Haloperidol 能抑制 LPS 刺激 THP-1 細胞所誘發產生 total IκB-α之降 解作用
在 Figure 16 中,當未投予 LPS 時,因無訊息之傳遞,total IκB-α則無 法進行降解作用,故IκB-α蛋白的表現為最高(lane 1:1 ± 0.0),以 LPS 刺激90 分鐘(land 2:0.68 ± 0.04 fold),投予不同濃度之 haloperidol,可 發現IκB-α含量隨著 haloperidol 濃度的增高(land 3:2 μM,0.80 ± 0.01 fold;land 4:10 μM,0.78 ± 0.02 fold;land 5:20 μM,0.82 ± 0.01 fold) 而逐漸上升,根據以上所觀察到之結果,可以推論haloperidol 可能會經 由抑制IκB-α蛋白質的降解作用而更進一步的減少 NF-κB translocate 至
細胞核內部的作用,造成LPS 所誘導出來的 MMP-9 因此被抑制而無法
表現。
十四、探討 haloperidol 在 THP-1 細胞中對於 LPS 所誘發的內 生性 TNF-α表現作用
過去的研究曾指出,在單核球與巨噬細胞中,LPS 對於 TNF-α之生 成是一種強效的誘發劑。而在THP-1 與 PBMCs 中,LPS 皆會誘導非 常顯著的TNF-α生成(Kumar Mangalam, 2002)。若給予 TNF-α的抗體 則會抑制MMP-9 的生成還有 TNF-α mRNA 之表現與 proMMP-9 蛋白 的釋放(Leber, 1998)。而其他學者指出,在 AML 患者中,能偵測到明 顯的TNF-α表現量(Cimino, 1991)。另外於 Ismair et al 實驗中發現,在 acute promyelocytic leukemia (APL) cell line 裡,內生性的 TNF-α能選 擇性的調節 MMP-9 之表現(Ismair, 1998)。由以上的研究顯示 TNF-α 對於MMP-9 蛋白表現之重要性,我們取得實驗之上清液(supernatant) 並使用Human TNF-α ELISA system 來測其 TNF-α之變化。首先,根 據本實驗室所做的結果,在Figure 17 中,未給予 LPS 刺激時(resting),
以5x105 cells 為單位,便可偵測到些許程度之 TNF-α蛋白表現量(3.72
± 0.30 pg/5x105 cells),之後分別以 LPS 濃度為 10 ng/ml、50 ng/ml 與 100 ng/ml 所誘發的 TNF-α表現量分別為 9.56 ± 0.88 pg/5x105 cells (10 ng/ml)、14.26 ± 0.79 pg/5x105 cells (50 ng/ml)與 14.46 ± 0.84 pg/5x105 cells (100 ng/ml),由上述結果顯示,發現以 LPS 濃度為 50 ng/ml 所誘
發出來的 TNF-α表現量已達最高值,故之後之實驗均以此為刺激濃
度。由Figure 18 中可觀察到,同樣無 LPS 刺激下(resting),以 5x105 cells 為單位,便可偵測到些許程度之 TNF-α蛋白表現量(3.72 ± 0.30 pg/5x105 cells),隨後以 LPS 50 ng/ml 誘發 TNF-α表現 14.26 ± 0.79 pg/5x105 cells,再以不同濃度之 haloperidol (0.5, 2, 10, 20 μM)處理後,
發現均有意義地使TNF-α之蛋白表現量上升,分別為 0.5 μM,20.04 ± 0.59 pg/5x105 cells;2 μM,21.34 ± 0.81 pg/5x105 cells;10 μM,25.05 ± 0.47 pg/5x105 cells;20 μM,22.49 ± 1.35 pg/5x105 cells。根據以上結果 顯示,haloperidol 的藥物作用並不是因為影響內生性 TNF-α的表現而 對MMP-9 蛋白產生抑制。
Discussion
無 論 在 癌 症 或 中 樞 發 炎 疾 病 中 , 基 質 金 屬 蛋 白 酵 素(matrix metalloproteinases,MMPs)扮演著極為重要的角色。人類癌症的進程中,
不 僅 癌 細 胞 本 身 可 釋 出MMPs 之 外 , 腫 瘤 周 圍 的 基 質 細 胞 (stromal cells),包括內皮細胞(endothelial cells)、纖維母細胞(fibroblasts)、肌纖維 母細胞(myofibroblasts)、發炎細胞(inflammatory cells)也會釋放MMPs (Klein et al., 2004)。在中樞神經系統方面(CNS),所有的中樞神經細胞,
如:神經元(neurons)與神經膠質細胞(glia)皆可產生MMPs (Yong et al.,
1998),而異常之活化MMPs可分解基底層(basal lamina)的主成份,導致 血腦障壁(blood-brain barrier,BBB)的破壞,促使神經發炎產生許多中樞 神經性發炎疾病(如:多發性硬化症、阿茲海默症與惡性神經膠質瘤等) (Rosenberg et al., 1992; Anthony et al., 1998; Asahi et al., 2001)。而其中 MMP-9與神經退化的過程息息相關,原因是因為會導致神經的髓鞘脫落 (demyelination) (Gijbels et al., 1993)引起多發性硬化症之疾病。另外,在 急性白血病患者之病理發展過程中發現與MMP-9的關係最為密切。所以 MMP-9無論在神經發炎性疾病或白血病當中所扮演之角色需要被更加 重視。
MMP-9(屬於type IV collagenase,又稱為gelatin B,分子量為92 kD) 可 在 許 多 種 類 的 單 核 球 細 胞 中 被 發 現 , 包 括 : 周 邊 血 液 的 單 核 球 (peripheral blood monocytes)、組織中的巨噬細胞(microphages)、Kupffer cells與蝕骨細胞(osteoclasts)等皆可發現MMP-9的表現(Welgus et al., 1990; Masure et al., 1993; Winwood et al., 1995; Swallow et al., 1996)。另外 在一些leukemic cell lines中,包括:HL-60、NB4、U-937及THP-1也可釋 放MMP-9酵素(Ries et al., 1994; Ismair et al., 1998; Saarialho Kere et al., 1993; Van et al., 1991)。在一般正常生理中,成熟白血球(leukocytes)本身 就可以分泌MMP-9,利用MMP-9分解基質的能力,使白血球能從血液離 開而滲入周邊組織發炎處來發揮它們的免疫功能(Doherty et al., 1994;
Welgus et al., 1990; Leppert et al., 1995; Weiss et al., 1986)。另外,單核球 細胞若受到一些細胞激素(如:TNF-α、IL-1β、CSFs或IL-3)或細菌內毒 素(LPS)刺激時,便會使MMPs表現。有文獻指出,單核球(monocytes)若 是受到細胞激素TNF-α或IL-1β的刺激,便會誘發MMP-9的表現而不是 MMP-1 (Saren et al., 1996)。在單核球的細胞株裡,MMP-9的分泌與基因
轉錄的程度有關且可被一些細胞激素(如:TNF-α透過TNFR1)來促進調節 製造(Ries and Petrides, 1995; Ismair et al., 1998)。另外,LPS則會誘發顯 著的MMP-1與MMP-9表現(Lai et al., 2003)及許多有關發炎的細胞激素 (cytokines) (Yong et al., 1998)。而在一些神經退化性疾病中(如:阿茲海 默症與帕金森氏症)發現有些細胞激素的表現比較活躍,其中包括:
TNF-α、IL-1及TGF-β (Akiyama et al., 2000; Nagatsu et al., 2000),而其 中,TNF-α被認為是誘發發炎的強效細胞激素(Munoz-Fernandez et al., 1998)。
而本實驗所用的實驗細胞為THP-1細胞,是從白血病(leukemia)患者 所取得的單核球(monocytes)加以分離培養而得,有些特性則與單核吞噬 細胞(mononuclear phagocytes)很相似(Combs et al., 1999; Hsu et al., 1996;
Giulian et al., 1990; Prieto et al., 1994; Tsuchiya et al., 1982; Yates et al., 2000),且可對於一些細胞激素(如:IL-α、IL-1β、IL-6、IL-8、IL-12與 TNF-α)產生反應(Murthy et al., 2000; Szczepanik et al., 2001; Yates et al., 2000)。而許多文獻指出,在所有的神經退化性疾病中(如:阿茲海默症 及帕金森氏症等),與microglia細胞的活化有關(Chen., 2003; Liu et al., 2003; Nakamura, 2002; Akiyama et al., 2000),另外根據此篇學者所做出的 實 驗 中 發 現 , THP-1 細 胞 可 模 擬 中 樞 神 經 內 的 microglia 細 胞 (microglia-like human THP-1 cells) (Klegeris et al., 2005),所以本實驗使用 THP-1 cell lines來當作我們的實驗細胞,加入不同濃度的中樞神經用藥 haloperidol (0.5~20 μM)且加入刺激劑TNF-α (10 ng/ml)誘發MMP-9表 現,觀察藥物是否對MMP-9的活性產生影響,進而探究其中的藥理機制。
由電泳酵素分析法(gelatin zymography)之實驗結果證實,在THP-1細 胞中,haloperidol能抑制以TNF-α所誘發的MMP-9的活性表現且達到統
計上的意義,其抑制50%反應之濃度(IC50)分別為5.4 ± 0.3 μM;由此實驗 結 果 得 知 , 在THP-1 細 胞 中 TNF-α 所 誘 發 的 MMP-9 活 性 皆 能 隨 著 haloperidol的藥物濃度升高而使得MMP-9活性表現量減少,呈現濃度效 應(concentration-dependent)之抑制作用。接下來,為了證明haloperidol 在THP-1細胞中抑制MMP-9的活性表現,並不是因為引起細胞毒性而使 細胞減少所產生的結果,所以我們利用了MTT assay得知haloperidol在濃 度40 μM以上存活率就已低於60%。故本實驗所使用的haloperidol藥物濃 度範圍內(0.5~20 μM),細胞存活率不會有明顯減少之情形(Figure 18),
表示本實驗所使用之藥物濃度範圍並不會明顯造成細胞毒性。另外,為 了瞭解haloperidol是否會直接影響MMP-9本身的酵素活性表現,由實驗 結果觀察到,haloperidol對於MMP-9的酵素活性不會產生影響。
由Western blot實驗方法得知,在THP-1細胞中,haloperidol可明顯且 有 意 義 地 抑 制 以TNF-α 所 誘 發 產 生 的 MMP-9 蛋 白 表 現 , 且 隨 著 haloperidol之藥物濃度增加而逐漸減少MMP-9蛋白的表現,並且皆呈現 濃度效應(concentration-dependent)的抑制作用。發現藥物在較低濃度(0.5, 2 μM)時就可以有效地壓制MMP-9的蛋白表現,且於濃度20 μM壓制情形 更顯著。另一方面,於正常生理情況下,體內MMPs活化需獲得平衡調 節,故體內含有內生性抑制成份TIMP-1 (28.5 kD glycoprotein)。此蛋白 成分可被許多種類的細胞(包括:纖維母細胞、內皮細胞、單核球細胞或 巨噬細胞)所分泌,且可與MMP-9蛋白以1:1的方式形成複合物進而抑 制MMP-9的蛋白表現(Goldberg et al., 1992)。但在某些疾病上,特別是擁 有血液異常之患者或患有多發性硬化症之病人的腦脊髓液中皆可發現 TIMP-1之數值明顯增高(Murate et al., 1997; Őzenci et al., 1999),而本實 驗所用的細胞株(THP-1 cells)為罹患白血症之病人身上分離而得的單核
球 細 胞 , 利 用TNF-α 刺 激 劑 可 誘 發 TIMP-1 蛋 白 表 現 (Figure 25) , haloperidol於較低濃度(0.5 μM)時就可抑制由TNF-α所引發的TIMP-1蛋 白表現,當haloperidol到較高濃度時(20 μM)即可有意義地壓抑TIMP-1的 蛋白表現,顯示haloperidol對於不正常生理情況下所產生的調節成分 (如:MMP-9與TIMP-1)可能可以壓制其表現,對於某些疾病似乎可以提 供一個不錯的治療方針。接下來,我們推測haloperidol可能抑制的機轉 在MMPs蛋白質轉譯(translation)之上游部分(upstream)。
一般而言,基因訊息DNA會經由特定轉錄(transcription)解碼而形成 mRNA,之後mRNA再經由轉譯(translation)製造出特定蛋白質。故我們 利用RT-PCR之實驗方法觀察haloperidol是否可以抑制MMP-9 mRNA的 產生。由實驗結果發現haloperidol確實隨著濃度升高而可以抑制MMP-9 的mRNA表現(Figure 24)且藥物濃度於10 μM以上則可以有意義地壓制 MMP-9之mRNA表現。然而,對於藥物是否會影響已轉錄後之mRNA穩 定性(mRNA stability),仍需進一步研究探討。因此再更深一層地探討 MMP-9基因轉錄過程的上游所受到相關之影響。
在不同細胞株裡,MMP-9的表現似乎可被許多不同的訊息傳遞路徑 或轉錄因子所調節 ,如:Mitogen-activated protein kinases (MAPKs) pathway、Nuclear factor-κB (NF-κB) pathway (Moon et al., 2004; Bond et al., 1998)、轉錄調節因子(NF-κB與AP-1) (Sato and Seiki, 1993; Gum et al., 1996)。而TNF-α可以刺激單核球和巨噬細胞產生及釋放大量的MMPs,
其主要的路徑就是活化NF-κB (Sarén et al., 1996; Ismair et al, 1998; Li and Verma, 2002)。在神經系統內的神經膠原細胞中,NF-κB可被許多種 物質所活化,其中包括:neurotrophic factors、cytokines與neurotransmitters 而大量表現(Bhakar et al., 2002; O’Neill and Kaltschmidt, 1997; Yalcin et
al., 2003; Carter et al., 1996; Guerrini et al., 1995; Hamanoue et al., 1999;
Kaltschmidt et al., 1995)。另外,若活化NF-κB則可能造成神經死亡(Pizzi et al., 2002; Shou et al., 2002),導致神經退化失調與發炎現象產生(Bhakar et al., 2002; Blondeau et al., 2001; Fridmacher et al., 2003; Mattson et al., 2000)。在許多文獻顯示,轉錄因子NF-κB與癌症之形成有緊密的相互關 係(Sun et al., 2003; Suh and Rabson, 2004)。而NF-κB的活化主要機轉是將 NF-κB 的 抑 制 型 蛋 白 IκB-α 磷 酸 化 後 分 解 掉 , 促 使 NF-κB 由 核 外 translocate至核內活化,啟動各種基因表現,進而造成許多發炎性相關因 子產生(包括:MMPs與cytokines) (Li and Verma, 2002; Panwalkar et al., 2004)。故本實驗便針對NF-κB之路徑進行探討。首先,利用Western blot 之實驗方法觀察NF-κB的抑制型蛋白IκB-α之表現情形,探討haloperidol 是否經由抑制IκB-α的降解作用而降低MMP-9蛋白之表現,並藉此瞭解 haloperidol在IκB-α磷酸化的分解作用與在NF-κB的訊息傳遞路徑上所扮 演的角色。由Western blot分析結果顯示,TNF-α作用15分鐘後可明顯地 引發THP-1細胞內IκB-α之降解反應,隨著時間的增加則會漸漸地回復到 基準值。當投予不同濃度之藥物haloperidol後,在Figure 26中可發現隨著 haloperidol的濃度增高(0.5~10 μM),以TNF-α刺激的IκB-α蛋白降解表現 量會逐漸回升,在較低濃度時(0.5, 2 μM)抑制IκB-α降解情況差不多,但 在haloperidol濃度為10 μM時抑制IκB-α降解程度較濃度為20 μM的效果 好,不過濃度20 μM時還是可以減少其降解程度,顯示haloperidol可以抑 制由TNF-α刺激所導致的IκB-α之降解作用。因此,我們推測haloperidol 可能主要是經由抑制IκB-α的降解作用,而減少由TNF-α刺激所引發的游 離態NF-κB進入細胞核與MMP-9之promoter序列結合,使得MMP-9的基 因表現減少。有文獻指出,必需磷酸化抑制型蛋白IκB-α才能使得NF-κB
活化,其中扮演著磷酸化IκB-α之上游酵素為IKK complex (Yamaoka et al., 1998; Zandi et al., 1997),其直接作用(磷酸化)在IκB-α之Ser 32與Ser 36位置上(DiDonato et al., 1997; Lee et al., 1997; Mercurio et al., 1997;
Zandi et al., 1997; Zandi and Karin, 1999)。然而haloperidol是否作用於 IκB-α之上游酵素(如:IKK)則仍需進一步探討。
根據上述的實驗結果推論 haloperidol 可能是經由抑制 IκB-α蛋白之降 解 作 用 而 達 到 抑 制 TNF-α刺激所誘發的 MMP-9 表現。為了確認 haloperidol 對 THP-1 細胞以 TNF-α誘發釋放 MMP-9 蛋白表現的過程
中,其 NF-κB 進入細胞核內的表現情形。因此,我們萃取細胞核內的蛋
白質並利用Western blot 觀察 NF-κB (p65)在細胞核內外的表現。TNF-α 刺激 30 分鐘之後,細胞質中的 p65 表現量會降低,細胞核內 p65 所表 現的量會明顯的增加,表示 p65 由細胞質 translocate 至細胞核內。當投 與不同濃度的haloperidol 藥物處理後(0.5, 2, 10 μM),在細胞質發現藥物 濃度於10 μM 即開始具有 p65 反轉情形,但不具統計上的意義。在細胞 核中,觀察到 haloperidol 在濃度於 10 μM 時則可有意義地抑制 p65 translocate 至細胞核。顯示 haloperidol 似乎在藥物濃度為 10 μM 時便能
抑制由 TNF-α所誘發的 p65 蛋白表現。另外,為了更進一步地確認在
THP-1 細胞核內,haloperidol 是否亦會影響 NF-κB 與 DNA binding 的表 現,而使得 NF-κB 轉錄轉譯出的 MMP-9 蛋白表現量減少,故本實驗利 用電泳移動偏向分析法(electrophoretic mobility shift assay, EMSA)分析 當投予不同濃度的haloperidol 時,再加入刺激劑 TNF-α,觀察可否影響 NF-κB 與 DNA binding 的程度。由 Figure 28 中,加入 TNF-α刺激 30 分 鐘後,發現 DNA 明顯產生 shift 現象,當投與不同濃度之 haloperidol (0.5~10 μM),可發現 shift 現象有減少的情形,而濃度為 0.5 μM 時即可
有意義地抑制 THP-1 細胞核內 TNF-α所誘發的 NF-κB 活化情形,但濃 度為2 μM 時此抑制效果卻減少,不過較高濃度 10 μM 發現可明顯抑制 NF-κB 活化,此結果與 p65 的 translocation 結果相符,顯示 haloperidol 可以減低核內NF-κB 與 DNA binding 的程度。綜合以上實驗結果,我們 更加可以確立,haloperidol 可能是經由 NF-κB 的訊息傳遞路徑來抑制 MMP-9 的表現。
許 多 常 見 的inducible MMPs (MMP-1, -3, -9 與 -13) 含 有 activator protein-1 (AP-1)轉錄因子結合區域(binding site),在它們近側的promoter 上需要細胞激素的作用(如:TNF-α)而被誘發。AP-1轉錄因子是由c-fos 與 c-jun 蛋 白 所 組 成 的 heterodimers , 這 些 蛋 白 會 經 由 mitogen-actived protein kinases (MAPKs)的訊息傳遞路徑而被合成與活化(Firestein and Manning, 1999)。近來主要被研究的MAPKs有三種,包括:extracellular signal-regulated kinases (ERKs)、c-Jun N-terminal kinase (JNK)與p38 MAPK等。而MAPKs在訊息傳遞過程中,必需由上游之不同MAPK/ERK kinase 分別將其磷酸化,轉為活化態之蛋白酵素,才能進一步地調控轉 錄因子來進行基因的表現(Davis, 1994; English and Cobb, 2002)。之前有 文獻提到在哺乳動物腦中會導致神經退化的可能路徑主要是經由JNKs 而活化c-Jun/AP-1 (Waetzig and Herdegen, 2004),而JNKs (包括三個 isoforms:JNK1, JNK2, JNK3)需被更上游的MAPK kinases (MAP2K,如:
MKK4與MKK7)與MAPK kinase kinases (MAP3K,如:MLKs或ASK)活 化(Herdegen and Waetzig, 2000; Rincon et al., 2001; Waetzig et al., 2004)。
而大部份慢性神經退化疾病(如:阿滋海默症與帕金森氏症等)或實驗動 物 模 式 中 , 經 常 與p38 MAPK這條路徑的活化很有關係(Koistinaho, 2002)。另外p38 kinase廣泛地分佈於哺乳動物之腦組織中,並且認為p38
的活化與神經內的壓力反應有關,但對於p38在神經系統中的角色尚未 非常明確(Mielke and Herdegen, 2000)。所以我們假設haloperidol有可能透 過MAPKs/AP-1路徑來影響MMP-9蛋白之產生及表現。因此在本實驗的 THP-1細胞中,我們主要針對JNK signal進行探討。在Figure 29中,加入 TNF-α刺激劑可造成磷酸化JNK蛋白質表現量有增加之情形,分別處理 不同濃度haloperidol (2, 10, 20 μM)後,發現磷酸化JNK的表現在濃度為2 與10 μM時,會些微被抑制但不具有統計上之意義,而較高濃度(20 μM) 時抑制情形較為明顯,因此我們推測haloperidol濃度為20 μM時能顯著地 抑制MMP-9蛋白表現,有可能部份是藉由抑制JNK的表現,相對地也可 能影響轉錄因子AP-1,進而降低MMP-9蛋白的表現量。
有許多文獻指出,MMPs (尤其是MMP-9)於細菌性腦膜炎中參與著 生理與病理過程,利用可以分解基質的能力進而破壞血腦障壁,使其通 透性變大引起更多細菌入侵(Rosenberg et al., 1995)。所以我們的實驗 中,使用白血球中的單核細胞THP-1 cells觀察若接觸到外在細菌性內毒 素LPS之刺激,探討THP-1細胞所產生的MMP-9蛋白表現與有給予藥物 haloperidol之下,藥物對於MMP-9的蛋白表現所產生的抑制作用。由 zymography 實 驗 方 法 可 觀 察 到 , haloperidol 能 抑 制 人 類 單 核 球 細 胞 (THP-1)中,LPS所誘發的MMP-9酵素活性表現(Figure 30 A)且呈現 concentration-dependent的抑制作用。在LPS (50 ng/ml)的刺激之下,
haloperidol對於MMP-9酵素抑制50%的反應濃度(IC50)為3.1 ± 0.8 μM (Figure 30 B)。接著利用Western blot分析haloperidol能抑制LPS在人類單 核 球 細 胞(THP-1) 中 所 誘 發 的 MMP-9 蛋 白 表 現 量 且 呈 現 顯 著 的 concentration-dependent抑制作用。
Nuclear factor-κB (NF-κB)這條訊息傳遞路徑控制了許多誘發性的發
炎基因與MMPs的表現,且文獻中提到LPS在單核球細胞中,可藉由活化 NF-κB/Rel轉錄因子而產生許多種類之基因表現,包括:TNF-α與IL-1等 (Sweet and Hume, 1996)也可活化許多第二傳遞因子與訊息傳遞路徑 (Sweet and Hume, 1996; Weinstein et al., 1992; Han et al., 1994; Hambleton et al., 1996)。我們利用Western blot實驗方法觀察NF-κB之抑制性因子 IκB-α蛋白表現量,藉以瞭解藥物haloperidol對於IκB-α磷酸化的分解作 用與在NF-κB訊息傳遞路徑所扮演的角色。發現LPS刺激90分鐘時所取 得的細胞lysate,IκB-α蛋白表現呈現明顯的降解作用。接下來以90分鐘 為LPS刺激之基準,分別投與不同濃度的haloperidol之後,可觀察到 haloperidol能抑制LPS在THP-1細胞中所引發的IκB-α降解作用,使得 NF-κB無法translocate至細胞核中活化MMP-9基因表現,造成MMP-9的 酵素與蛋白表現量減少。有文獻所做的實驗結果顯示,利用LPS刺激人 類單核球細胞與細胞株(THP-1 cells) (O'Connell et al., 1998; Guha and Mackman, 2001),使得轉錄因子NF-κB translocate至核內,可能藉由活 化IKKβ蛋白而導致的結果(O'Connell et al., 1998)。可提供本實驗更進一 步探討。而我們的實驗當中發現,在THP-1細胞中利用TNF-α與LPS刺激 時所誘發的IκB-α蛋白降解之時間有明顯的差異(TNF-α:15分鐘;LPS:
90分鐘) (Figure 25與32),可能原因為此兩種刺激劑活化IKKs時,牽涉到 不同之路徑所導致的(O’Connell et al., 1998; Hawiger et al., 1999; Fischer et al., 1999)。而另一篇文獻則顯示LPS刺激人類單核球細胞可經由許多 訊息傳遞路徑,其中包括IKK-NF-κB路徑與MAPKs路徑(ERK1/2; JNK;
p38)且牽涉到轉錄因子NF-κB (p50/p65)與AP-1 (c-Fos/c-Jun)。Lai等學者 則提出在人類周邊血液分離而得的單核球細胞,利用LPS刺激所誘發的 MMP-9蛋白或mRNA表現主要是受MAPKs其中之ERK1/2路徑所調控的
(Lai et al., 2003)。另外最新文獻則提到,也是利用LPS刺激人類周邊血液 之單核球細胞,發現可經由phosphatidylinositol-3/Akt/IKKα/NF-κB路徑 誘發MMP-9產生(Lu and Wahl, 2005)。而本實驗結果顯示haloperidol可明 顯地抑制由LPS所誘發的MMP-9酵素與蛋白的表現(Figure 30與31),但卻
無法明顯地抑制由LPS所誘發的IκB-α降解作用(無法顯著地回到基準
點),顯示藥物haloperidol可能對於LPS所誘發的MMP-9蛋白表現並非單
純壓制NF-κB這條路徑,針對此點需再做更進一步之探討。也証實
haloperidol的藥物作用並不是影響內生性TNF-α的表現而抑制MMP-9蛋 白的產生。顯示haloperidol可以選擇性抑制過量之TNF-α所引起的病理現 象。在未來的展望與研究方面,我們期許能藉由進ㄧ步的活體實驗,將 haloperidol視為老藥新用的治療目標,瞭解藥物如何調控in vivo中MMP-9 的表現與重要性。並更進一步地評估藥物在預防或治療中樞神經系統發 炎疾病。
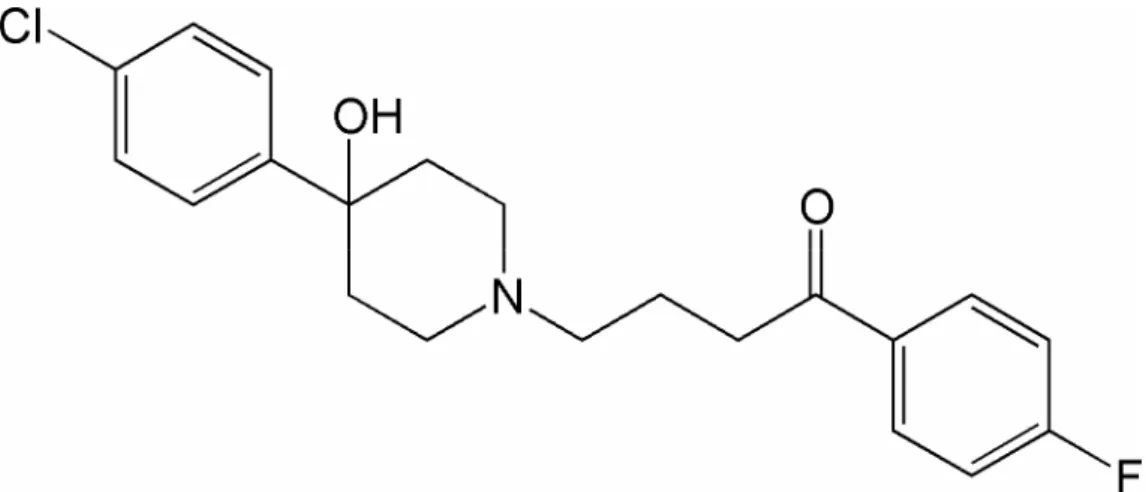
Figure 1. Chemical structure of haloperidol.
Figure 2. Effects of haloperidol on TNF-α-induced enzymatic activity of matrix metalloproteinase-9 (MMP-9) in THP-1 cells. THP-1 cells (1×106 cells/ml) were dispensed on 24-well plates and treated with TNF-α (10 ng/ml) for 24 hours as indicated. Cells were treated with the indicated concentrations of haloperidol (lane 3, 0.5 μM; land 4, 1 μM; lane 5, 2 μM;
land 6, 10 μM; land 7, 20 μM) or vehicle (land 2) for 15 minutes before treatment with TNF-α. Cell-free supernatants were then assayed for MMP-9 activity by gelatin zymography, as detailed in “Methods” (land 1, control). Percent inhibition is presented as mean ± S.E.M. of four to seven independent experiments.
Haloperidol (μM)
0.5 1 2 10 20
Inhibition %
0 20 40 60 80 100
MMP-9→
(92 kD)
1 2 3 4 5 6 7
Resting Vehicle 0.5 1 2 10 20 Haloperidol (μM)
TNF-α (10 ng/ml)
A.
B.
R DMSO 10 20 40 50 100
Viability (Ab 550 nm)
0.0 0.2 0.4 0.6 0.8 1.0 1.2
Haloperidol (μM)
***
** ***
***
Figure 3. Cytotoxicity of haloperidol on THP-1 cells. THP-1 cells were treated with different concentration of haloperidol (10-100 μM) and incubated for 24 hrs. Cell viability was measured by a colorimetric assay at 550nm based on the ability of mitochondria to reduced the tetrazolium dye 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) in viable cells. Percentage of viability is presented as mean ± S.E.M. of three to eight independent experiments. * P < 0.05; ** P < 0.01; *** P < 0.001 as compared with the resting.
R Vehicle 0.5 2 10 20
MMP-9 activity (folds)
0 1 2 3 4 5 6
Haloperidol (μM) TNF-α (10 ng/ml)
###
1 2 3 4 5 6 7
Resting Vehicle 0.5 2 10 20 Haloperidol (μM)
TNF-α (10 ng/ml) MMP-9→
(92 kD)
Figure 4. Evaluation of the direct effect of haloperidol on MMP-9 enzymatic activity by conditioned medium from TNF-α-treated cells. THP-1 cells (1×106 cells/ml) were dispensed on 24-well plates and treated with TNF-α (10 ng/ml) for 24 hours as indicated. Haloperidol (lane 4, 0.5 μM; lane 5, 2 μM; lane 6, 10 μM; lane 7, 20 μM) were added to incubation buffer for 17 hrs. The data are representative example of four experiments. ### P < 0.001 as compared with the resting.