國立臺灣大學醫學院分子醫學所 碩士論文
Graduate Institute of Molecular Medicine College of Medicine
National Taiwan University Master Thesis
SIK2 對細胞自噬的調控
Regulation of Autophagy by SIK2
莊淨婷
Ching-Ting Chuang
指導教授:呂勝春 博士 Advisor: Sheng-Chung Lee, Ph.D.
中華民國 100 年 7 月
July, 2011
摘要
本文主要探討 SIK2 在細胞自噬中扮演的功能。SIK2 為 AMPK 家族的一員,
目前已知參與於脂肪細胞的分化和胰島素訊號傳導的調控。SIK2 的其他功能目前 尚 待 研 究 。 先 前 我 們 實 驗 室 發 現 SIK2 藉 由 和 p97/VCP 之 交 互 作 用 而 調 控 ER-associated protein degradation (ERAD),也發現 SIK2 可能參與調控蛋白 質聚集體的降解。在本篇研究,進一步證實 SIK2 可能參與調控細胞自噬。當使用 MG132 來誘導包涵體和聚集體的形成時,發現 SIK2 和 p62 或是 HDAC6 之間有交 互作用或複合體之存在。當細胞表現正常激酶活性的 SIK2 時,會導致泛素化蛋 白質和 LC3-II 減少,顯示 SIK2 極有可能參與處理蛋白質聚集體,且 SIK2 可能 透過和 p97/VCP 之交互作用並和 p62 、HDAC6 或是 Hsp90 複合體之形成達到蛋 白質聚集體的降解。另外,在蛋白質分泌的實驗中發現, SIK2 亦可能藉由調控 細胞胞器而影響了蛋白質的分泌。本論文之結果顯示: SIK2 不但在蛋白質聚集體 的降解扮演主要調控功能它也極可能在 ERAD 和細胞自噬之間扮演協調者之角色。
關鍵字: SIK2、p62、HDAC6、Hsp90、包涵體、蛋白質聚集體
Abstract
SIK2 (salt-inducible kinase 2) belongs to members of AMPK family. Other than the regulation of adipocyte differentiation and insulin signal transduction, the functions of SIK2 remain largely unknown. Previously, we showed that SIK2 can interact with p97/VCP to regulate ER-associated protein degradation (ERAD). SIK2 may also involve in inclusion body or aggresome processing. In this study, I investigate how SIK2 is involved in regulation of autophagy. SIK2 could be found in complex containing either p62/SQSTM1 or HDAC6 when the cells are treated with proteosome inhibitor MG132. Overexpression of WT-SIK2 resulted in decrease of ubiquitinated proteins and LC3-II levels. SIK2 may facilitate aggresome processing through p62, HDAC6 or Hsp90. In a ligand-induced protein secretion system using human growth hormone as reporter indicates that SIK2 plays crucial roles in autophagy-mediated protein secretion. My study demonstrated that SIK2 could serve as a positive regulator in autophagic-mediated protein degradation and secretion. Together these results suggest that SIK2 may function as a coordinator for the ER stress response and autophagy.
Key words: SIK2, p62, HDAC6, Hsp90, inclusion body, aggresomes
Contents
Master thesis ………..………. i
中文摘要 ………..……… ii
Abstract ………..………. iii
Contents .………...………. iv
Introduction ..……….……….. 1
Material and methods ..……… 5
Plasmids, Constructs and Antibodies ……….. 5
Cell cultures and transfections ………. 5
Chemicals treatment ……… 6
Preparation of Whole Cell Extracts ………. 6
Immunopreciptation assay ………...… 6
Flow cytometry ……… 7
Ligand-induced hGH secretion assay ………..……. 7
Results ..……… 8
SIK2 may form complex with p62 and HDAC6 ………..…….... 8
SIK2 facilitates the processing of inclusion bodies and aggresomes ……….. 8
Cooperation between SIK2 and HDAC6 facilitates autophagy …………..…. 9
Both kinase activity of SIK2 and chaperone activity of Hsp90 are important for processing of autophagosomes containing SIK2, p97/VCP and Hsp90 ..……… 11
SIK2-regulated vesicle dynamics or autophagy mediates protein secretion from a ligand-induced GH reporter system ………..……….…. 12
SIK2 is important for cell survival when the function of proteosome is impaired ………..………. 13
Discussion ..……….. 14
References ..………..……… 18 List of figures ..………. 23 Fig. 1 SIK2 may form complex with p62 and HDAC6 ………. 23 Fig. 2 p62 and SIK2-containing complex formation appeared
to be enhanced both in the soluble and insoluble
fractions when cells were treated with MG132. ……….. 25 Fig. 3 Complex formation between HDAC6 and SIK2 ……….. 27
Fig. 4 Both kinase activity of SIK2 and chaperone activity of Hsp90 are important for processing of autophagosomes
containing SIK2, p97/VCP and Hsp90 ……….. 31 Fig. 5 Autophagy is involved in regulation of ligand-induced
protein secretion by SIK2 ……… 33 Fig. 6 SIK2 is important for the cell survival under stress conditions ……… 35
Introduction
Salt-inducible kinase 2 (SIK2) was first identified by searching the genomic database for the isoforms of SIK1 [1]. SIK2 belongs to the AMP-activated protein kinase (AMPK) family, SIK subfamily. SIK2 is a serine/threonine kinase containing 926 amino acids with three distinct domains: a kinase domain, an UBA domain, and the polyGln-rich C-terminal region [2]. Studies about gene expression pattern showed that SIK2 mRNA was abundant in adipose tissue, and has little in the testis of mouse [1, 3].
Previous reports showed that SIK2 expression is up-regulated during adipose differentiation. Human IRS-1 can be phosphorylated at Ser794 by SIK2 leading to attenuation of the insulin signal transduction. Both the activity and level of SIK2 are elevated in diabetic animals, leading to insulin resistance [3]. In addition, the co-activator of CREB, TORC, can be phosphorylated by SIK2 at Ser171. The phosphorylated TORC recruits 14-3-3 protein and be exported from the nucleus to the cytoplasm to prevent nuclear CREB from interacting with TORC [4]. When cells are treated with cAMP, PKA phosphorylates SIK2 at Ser587, and the phosphorylated SIK2 lost its repressive activity on CREB [5]. Dentin et al. indicated that insulin induces AKT-mediated SIK2 phosphorylation at Ser 358 and disrupt the activity of TORC2. The results indicate that SIK2 plays a role in regulation of hormonal signal transduction.
When SIK2 is overexpressed, it can repress the expression of some lipogenic genes, including acetyl CoA carboxylase2 (ACC2), FAS, stearoyl CoA desaturase1 (SCD1), and reduced triglyceride content in cells [6]. These findings indicate that SIK2 also play an important role in adipocyte lipogenesis and metabolism. However, the role of SIK2 in other physiological processes remains poorly understood.
Recently, SIK2 has been found to be required for the ERAD process (unpublished data). It is proposed that SIK2 facilitates ERAD via directly phosphorylating p97/VCP to enhance its ATPase activity, which is important for the retro-translocation step of unfolded/misfolded proteins in the ER lumen. SIK2 physically interacts with p97, and colocalizes with p97 in the ER membrane. Preliminary studies suggest SIK2 plays the important role for the step of p97-mediated retro-translocation of misfolded proteins in ERAD. Published results demonstrate that p97/VCP is required for the formation and clearance of aggresomes [7-9]. Aggresomes are structures near the nucleus at the proximity of the microtubule-organizing center and its formation could decrease the toxicity of scattered small aggregates (inclusion bodies). In our lab, we found that SIK2 and p97 co-localized to the MG132 or TDP-43-induced aggresomes. Knockdown of SIK2 resulted in increase while overexpression of SIK2 caused decrease of inclusion bodies/aggresomes. Our results suggest that in addition to its functions in ERAD, SIK2 may also play important roles in autophagic-mediated protein degradation.
Autophagy is one of the major degradation mechanisms, dealing with the recycling and turnover of damaged or excess organelles and protein aggregates. The mTORC1 kinase complex is one of the important negative regulators of autophagy. In contrast, AMPK is a positive regulator that induces autophagy in response to energy depletion [10]. AMPK is a conserved cellular energy manager controlling energy homeostasis.
The ULK1 (mammalian homolog of yeast Atg1) kinase complex plays a major role in autophagy induction. Previous studies suggest that mTOR and AMPK was involved in autophagy inhibition and induction through phosphorylation of ULK1 [11].
Alternatively, it has been proposed that AMPK triggers autophagy through an indirect mechanism, inhibiting mTORC1 activity by phosphorylating TSC2 and Raptor [12, 13].
Sequestosome 1/p62 (also called SQSTM1) is a polyubiquitin-binding protein. p62 can link autophagy machinery to the polyubiquitinated proteins by its LC3 interacting domain to interact with LC3 and UBA domain and which is required for the formation of protein aggregates [14-16]. p62 has been found to co-localize with tau, huntingtin and α-synuclein in neurodegenerative disease [17, 18]. These studies indicate that p62-LC3 interaction is important in autophagy mediated clearance of inclusion bodies.
LC3 is a mammalian homolog of yeast Atg8. Atg8 can be cleaved at its c-terminal to generate form-I from pro-Atg8. Atg8 form-I can be converted to form- II by conjugating with phosphatidylethanolamine (PE). The form- II Atg8 can be recruited to the membrane of autophagosome. Atg8 is important in regulating autophagosome formation [19]. Recently, many studies have shown that LC3 is required for regulating autophagic activity and recruiting p62 to the autophagosome. Furthermore, p62 could interact with proteasome. Reduction of the level of p62 results in the inhibition of proteasome-mediated degradation and the accumulation of ubiquitinated protein aggregates. p62 participates both in autophagy and proteosome degradation machinery through its UBA domain linking to polyubiquitinated proteins [20-22].
Recent studies suggest that histone deacetylase HDAC also has functions other than transcriptional regulation. For example, HDAC6 are also involved in aggresome formation, and this function is depend on both its deacetylase activity and ubiquitin-binding domain [23]. Not long after this observation, HDAC6 was also reported to involve in autophagic clearance of mutant huntingtin aggregates [24]. It suggests a role for HDAC6 in processing of misfolded proteins. Furthermore, studies also showed that HDAC6-interacting chaperone, p97/VCP, can dissociate the HDAC6–ubiquitin complexes and decrease HDAC6 promoting the polyubiquitinated
proteins accumulation [25]. It proposed that p97/VCP could promote protein degradation, but HDAC6 would promote the formation of ubiquitinated protein aggregates.
In recent studies, researchers indicated that during Ras-induced senescence, there is a cellular compartment at the trans Golgi apparatus (GA), the TOR-autophagy spatial coupling compartment (TASCC). Autolysosomes and mTOR accumulated at TASCC and disruption of mTOR localization to the TASCC suppressed interleukin-6/8 synthesis.
The spatial integration of rough ER-GA, autolysosomes, and mTOR could augment their functions, leading to the efficient synthesis of secretory and lysosomal proteins.
Blocking autophagy, and thus caused loss of mTORC1 from the TASCC, would affect protein secretion. TASCC was also formed during macrophage differentiation and in glomerular podocytes, both had increased protein secretion. Since the TASCC can be found in diverse conditions, it may represent a general mechanism for rapid protein turnover [26]. In addition, recent results also highlight a novel role for autophagy which is not only involved in lysosomal degradative pathway but also participates in unconventional protein secretion which the secreted proteins neither have a signal sequence nor transport by the ER-Golgi pathway [27-29].
In this study, I examined the involvements of SIK2 in autophagy. I found that regulation of autophagy by SIK2 is dependent on its interaction with p97/VCP, p62 or HDAC6. In addition, SIK2 can regulate autophagy-dependent, ligand-induced protein secretion. My results suggest that SIK2 may play crucial roles in autophagic-mediated protein degradation and secretion. The present results also suggest that SIK2 could be a coordinator for ER stress response and autophagy.
Material and Methods
Plasmids, Constructs and Antibodies
The pBluescript II vector encoding SIK2 sequence (KIAA0781) was from HUGE.
The DNA fragment encoding SIK2 was excised using Sal I and Xho I sites in the pBluescript-SIK2 vector and in turn sub-cloned into pCMV-Flag mammalian expression vector (Stratagene). Flag-SIK2 KD (kinase dead, K49M) , Flag-His-p97 S770A was generated by using site-directed mutagenesis kit. pC4S1-FM4-FCS-hGH plasmid was kindly provided by ARIAD Pharmaceuticals, Inc. pLKO.1-shRNA: shLuc, and shSIK2 were obtained from National RNAi Core Facility (Institute of Molecular Biology / Genomic Research Center, Academia Sinica). The following antibodies were used in this study: monoclonal antibodies against ubiquitin (Millipore), rabbit antibodies against PARP1, LC3, ULK1, ULK1/S555P, and ULK1/S757P (cell signalling), polyclonal antibodies against growth hormone (GeneTex). Monoclonal antibodies to SIK2 (clone 15G10), anti-p97(clone 4G9), anti-tubulin-a(clone 10D8), and rabbit anti-SIK2/T175p, anti-Hsp27, anti-Hsp70, anti-AMPK/T172p, anti-AMPK, and anti-Flag antibodies were generated in our Lab.
Cell cultures and transfections
293T cells were cultured in Dulbecco’s modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 100 units/ml penicillin-streptomycin (GIBCO) at 37℃ in a humidified incubator containing 5% CO2. U2OS cells were maintained in McCoy’s 5A supplemented with 10% FBS. As the cell density reached 80%, the cells were rinsed with PBS and then trypsinized. The cells were dispersed by repeated pipetting over the surface and diluted to appropriate seeding
concentration.
293T cells were transfected by the calcium phosphate precipitation method. 12-16 hrs after transfection, the cultured medium was refreshed for recovery about 8 hrs and treated with different drugs. U2OS were transfected using turbofect according to manufacturer’s instruction. 4hrs after transfection, the cultured medium was refreshed for recovery, then for different drugs treatment.
Chemicals treatment
Different drug dosages and treatment time were used: 1 μM 17DMAG (InvivoGen) for 16hr, 10 μM compound C (Merck) for 12-16hr, 1mM AICAR (Biomol) for 12-16 hr, 5μM MG132 (Biomol) for 12-16hr, 0.2μM Bafilomycin (Biomol) for 12-16hr, 4μM AP21998 (ARIAD) for 1 hr.
Preparation of Whole Cell Extracts
The cells were washed with phosphate-buffered saline (PBS, PH 7.4) and pelleted by spinning for 3 min at 3000 rpm. Cell pellet was lysed directly with cold whole cell extraction (WCE) buffer (0.15M NaCl, 20mM Tris〔PH 7.6〕, 0.2% Triton X-100, 1mM PMSF, 1μg/ml leupeptin, and 1μg/ml pepstatin A, 0.5M NaF, Van, sodium butyrate, 1mM DTT, 25mM NEM). Lysis was performed with pipetting. The cell lysates were kept on ice for 20min, and then centrifuged for 15 min in 14,000 rpm at 4℃.. The pellet was dissolved in 1X sample buffer and used for further analysis.
Immunopreciptation assay
Whole cell extracts were incubated with anti-FLAG M2 beads (Sigma). After gently rotating for 1.5 hours at 4 ℃, the beads were washed with WCE buffer three times.
The precititated FLAG-tagged proteins were eluted by 1X SDS-PAGE sample buffer (8M urea, 300mM Tris-HCl(pH6.8), 20% β-mercaptoethanol, 9.2% SDS, and 0.2%
bromophenol Blue).
Flow cytometry
293T cells were washed twice with cold PBS and then resuspended in annexin V binding buffer (10 mM Hepes, pH 7.4, 140 mM NaCl, 2.5 mM CaCl2) at a concentration of 1x106 cells/ml. 100 μl of cell suspension were transfered to 5 ml test tube and add 5 μl of FITC Annexin V. The cells were gently vortexed and incubated for 15 min at 25 °C in the dark. After adding 400 μl of annexin V binding buffer to each tube, the cells were analyzed by flow cytometry.
Ligand-induced hGH secretion assay
hGH secretion assays were performed as described previously [30]. 293T cells were transfected with 4xFKBP-hGH and other plasmids. After 16 hours, the medium was refreshed. After 24 hours, the medium was then changed to serum-free medium containing secretion inducer, AP21998 (final concentration, 4μM). AP21998 was incubated with the cells for 1 h before the supernatant was collected. The supernatant was then concentrated by TCA precipitation and dissolved in 6M urea. Measuring protein concentration, equal amounts of protein was loaded and the protein level of growth hormone was measured by western blot assay.
Results SIK2 may form complex with p62 and HDAC6
To study the possible functions of SIK2 in autophagy, I tested whether SIK2 could interact with p62 and HDAC6, two proteins known to be important regulators for autophagy. I performed co-transfection of FLAG-p62 or FLAG-HDAC6 and His-SIK2 WT or His-SIK2 KD co-immunoprecipitation with M2 beads. Overexpressed His-SIK2 WT and His-SIK2 KD and endogenous SIK2 could be co-immunoprecipitated with FLAG- p62 or FLAG-HDAC6 (fig. 1, fig. 2A and fig. 3A). It appeared that SIK2 KD existed in either p62 or HDAC6 complex in larger amounts than SIK2 WT (fig. 1A).
Since p62 and HDAC6 both bind to polyubiquitinated proteins, I also want to know whether the interaction between SIK2 and p62 or HDAC6 will be affected by NEM treatment. NEM has been used as an inhibitor of deubiquitinases. The data showed the interaction between p62 or HDAC6 and SIK2 in the absence of NEM was decreased (fig. 1A). These results suggest that SIK2 may form complex with p62 or HDAC6, and this complex is likely composed of sequestered polyubiquitinated proteins.
SIK2 facilitates the processing of inclusion bodies and aggresomes
MG132 could block proteosome activity and induce aggresome formation. To examine whether the complex formation between SIK2 and p62 was affected by treatment with MG132, I performed co-transfection of FLAG-p62 and His-SIK2 followed by immunoprecipitation with M2 beads. I probed the levels of p62, SIK2 and the ubiquitinated proteins. After treating with MG132, both p62 and SIK2 levels in the immunocomplex were increased (fig. 2A, left and right panels). The results showed that when p62 co-transfected with SIK2-WT, the level of ubiquitinated proteins was significantly reduced as compared with that of p62 alone or cotransfected with
SIK2-KD (fig. 2A). We also examined the levels of p62, SIK2 and ubiquitinated protein in insoluble fraction. SIK2 as well as ubiquitinated proteins were highly enriched in the insoluble fraction, especially in samples prepared from cells treated with MG132.
Significantly, SIK2-KD was highly accumulated as compared with the endogenous SIK2 (i.e., p62 transfected cells) or SIK2-WT untreated or treated with MG132 (Fig. 2B, middle panel). The ubiquination level was increased after treating with MG132, but was decreased when SIK2-WT was overexpressed (Fig. 2B, top panel). Matured form of LC3 (LC3-II) was increased in insoluble fraction after treated with MG132, however, when SIK2-WT was overexpressed, LC3-II was less accumulated than that of SIK2-KD (fig. 2B, bottom panel). These data further suggest that MG132 treatment results in aggresome formation (and decreases small inclusion bodies). Together, these results suggest that SIK2-WT facilitates processing of inclusion bodies/aggresomes by autophagy.
Cooperation between SIK2 and HDAC6 facilitates autophagy
Some researchers clearly indicated the necessary role of HDAC6 catalytic activity for aggresome formation and autophagy promotion [31], but the data demonstrated by others suggest that HDAC6-dependent tubulin deacetylation should decrease the interaction of molecular motors with microtubules and interfere with these processes [32]. To address the functional cooperation between SIK2 and HDAC6 in inclusion bodies/aggresomes processing by autophagy, I performed co-transfection of SIK2 (WT and KD) and HDAC6 (WT and CD) and probed the levels of SIK2, HDAC6, ubiquitinated proteins and LC3 in the M2 immunoprecipitates (from supernatants) and pellets, respectively. Co-transfection of HDAC6-WT and SIK2-WT or SIK2-KD resulted in accumulation of supershifted species (presumably polyubiquitinated SIK2)
in the MG132 treated samples, especially when SIK2-KD was overexpressed (Fig. 3A top panel, left half). When HDAC6-CD was co-transfected with SIK2-WT or SIK2-KD, the level of SIK2 immunoprecipitated with overexpressed HDAC6-CD seemed higher but the supershifted species appeared weaker than the HDAC6-WT co-transfected samples (compare the right half to the left half of Fig. 3A, top panel).
The levels of FLAG-HDAC6-WT and FLAG-HDAC6-CD seemed the same (Fig. 3A, lower panel) while the ubiquitinated signal seemed stronger in the HDAC6-WT than the HDAC6-CD overexpressed immunoprecipitates (Fig. 3A, middle panel). The SIK2-KD was dramatically accumulated in the insoluble fraction regardless of HDAC6-WT or HDAC6-CD was co-expressed (Fig. 3B, top panel). When co-expressed with SIK2-WT, the level of HDAC6-WT appeared to be efficiently reduced as compared to the SIK2-KD or vector control untreated or treated with MG132 (Fig. 3B, lower panel).
Similarly, HDAC6-CD also appeared efficiently removed by SIK2-WT untreated with MG132 but not so when treated with MG132 (Fig. 3B lower panel). The levels of HDAC6 in the insoluble fraction seemed to correlate with the supershifted signals (presumably ubiquitinated HDAC6). The accumulation of LC3-II correlated with MG132 treatment (Fig. 3B, middle panel). Finally, the level of ubiquitinated signals in the insoluble fraction strictly correlated with the MG132 treatment (Fig. 3B). The levels of endogenous or overexpressed SIK2 in the supernatant fraction (before immunoprecipitation) were shown in Fig. 3C (top panel). The endogenous SIK2 appeared to be reduced when treated with MG132 whether overexpressed of HDAC6-WT or HDAC6-CD. The anti-FLAG antibody probed supershifted signals are especially prominent in overexpressed HDAC6-WT or HDAC6-CD treated with MG132 (Fig. 3C, middle panel). These results and the reduced endogenous SIK2 levels and the accumulation of LC3-II treated with MG132 correlate very well. Significantly,
the supershifted anti-FLAG antibody-probed signals were undetectable in the soluble fraction of co-expressed HDAC6-WT and SIK2. These results together with the results of Fig. 3B (top panel) suggest that both HDAC6-WT and HDAC-CD can efficiently sequester endogenous SIK2 to the insoluble fraction when treated with MG132. Taken together, these data indicate that cooperation between SIK2 and HDAC6 facilitates autophagy. Both the kinase activity of SIK2 and the catalytic activity of HDAC6 appear to be important!
Both kinase activity of SIK2 and chaperone activity of Hsp90 are important for processing of autophagosomes containing SIK2, p97/VCP and Hsp90
It is known that p97/VCP, Hsp90 and HSF1 or HDAC6 and Hsp90 formed mutually exclusive polyubiquitinated protein-containing complex. When p97/VCP is competed with HDAC6, HSF1 is released from the complex and Hsp70 and Hsp27 are induced. We have determined that Hsp90 interacts with SIK2 and only the SIK2-WT, not the SIK2-KD or SIK2/T175A mutant protein level is significantly decreased when Hsp90 is inhibited by 17DMAG (Fig.4 ). To test whether SIK2 is part of the p97/VCP, Hsp90 and HSF1 complex, we performed cotransfection of p97/VCP (both wild-type and ATPase dead mutant) and SIK2 (WT and KD). The level of SIK2-WT, not SIK2-KD was decreased in the soluble fraction when cotransfected with wild-type p97/VCP and treated with 17DMAG. Cotransfection of p97/VCP-AA with SIK2-WT and SIK2-KD seemed to result in the decrease the levels of both SIK2-WT and
SIK2-KD (Fig.4A , upper panel). When examined the levels of SIK2-KD and p97/VCP or p97/VCP-AA in the insoluble fraction, SIK2-KD and p97/VCP were further
accumulated when treated with 17DMAG (Fig.4B , upper panel). When cotransfected with p97/VCP-AA, SIK2-KD was not accumulated to the same level as compared to
cotransfected p97/VCP. Hsp70 and Hsp27 levels in the soluble or insoluble fraction were increased when treated with 17DMAG (Fig.4A and 4B, lower panel). These
results suggest that p97/VCP and SIK2-KD are sequestered in the insoluble aggresomes.
When Hsp90 is inhibited, the aggresome formation is further enhanced. Taken together, these and the results from the previous sections suggest that SIK2 kinase activity is likely required for the resolution of aggresomes by autophagy.
SIK2-regulated vesicle dynamics or autophagy mediates protein secretion from a ligand-induced growth hormone reporter system
Previous studies have shown that autophagy could participate in unconventional protein secretion [27], and AMPK could regulate autophagy. We treated 293 T cells with bafilomycin and compound C. Bafilomycin can inhibit vacuole H+-ATPase and vesicle dynamics/autophagy and compound C is an AMPK inhibitor. The secreted growth hormone (GH) was decreased when treated with bafilomycin or compound C (fig. 5A).
The levels of LC3-II accumulated in bafilomycin or compound C treated samples correlate well with the secreted GH level (fig. 5B). To address whether SIK2 plays any role in ligand-induced GH secretion, I performed knockdown of endogenous SIK2 or overexpressed SIK2-KD in 293T cells, the GH secretion was significantly impaired when the endogenous SIK2 was reduced or when the SIK2-KD was overexpressed (fig.
5C and D). The secreted GH level of overexpressed SIK2-WT was drastically reduced by bafilomycin or compound c treatment (fig. 5E). The LC3-II level was higher in SIK2-KD than SIK2-WT overexpressed cells. When treated with compound C, the level of SIK2 was significantly decreased. This result in conjunction with the inhibition of AMPK by compound C suggests that AMPK-activated and SIK2-regulated vesicle dynamics or autophagy is indeed responsible for the ligand-induced secretion of GH.
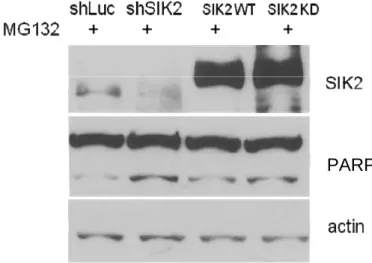
SIK2 is important for cell survival when the function of proteosome is impaired To address the importance of SIK2 for cellular homeostasis, I performed cell viability experiments under ER stress and disruption of SIK2. 293T cells transfected with pSuper-shLUC-dsRed or pSuper-shSIK2-dsRed were treated with MG132, stained with FITC-annexin-V and analyzed by flow cytometry. FITC-positive cells were scored among 10,000 cells counted (fig. 6A). I arbitrarily separated these FITC-positive cells into weak dsRed (dsRed+) and strong dsRed (dsRed++) groups. In the absence of MG132, the number of FITC-positive cells in SIK2 knockdown remained essentially no difference from control knockdown cells (fig. 6A, 6B). However, after treating with MG132, the ratio of FITC-positive cells in dsRed++ and dsRed+ of shSIK2 knockdown cells is 10.4. Contrary, the ratio of FITC-positive cells in dsRed++ and dsRed+ of shLUC control knockdown cells is 4. These results suggest that when the level of endogenous SIK2 is reduced, the cells are much more susceptible to MG132 treatment (fig. 6B). To further examine the apoptosis of SIK2 knockdown cells treated with MG132, I performed Western blot assay by probing the 89 kDa PARP-1. After SIK2 knockdown or SIK2-KD overexpression and MG132 treatment, the cleaved PARP-1 form was increased compared with shLUC or wild type SIK2 (fig. 6C, 6D).
Discussion
In this study, the function of SIK2 in inclusion body/aggresome processing was demonstrated. I found that there are at least two other proteins, HDAC6 and p62, could function with SIK2 in aggresome processing. Importantly, the kinase activity of SIK2 was required for autophagy. When SIK2-KD-transfected or endogenous SIK2 knockdown cells were treated with MG132, the ubiquitinated proteins and LC3-II were elevated. Results from our lab showed that p97 was targeted and its ATPase activity stimulated by SIK2. When p97-Ser770 was mutated to p97-SerAla (p97S770A), SIK2-induced inclusion body/aggresome processing was compromised. Co-transfection of SIK2-KD and p97-S770A into 293T cells results in the elevated accumulation of polyubiquitinated proteins in the insoluble fraction. These results suggest that SIK2 plays important functions in autophagy. My results clearly showed that both p62- and HDAC6-containing protein complexes are involved in SIK2-regulated vesicle dynamics/autophagy. The exact underlying mechanisms remain to be answered.
Moreover, the complex formed between SIK2 and p62 or HDAC6 was affected by ubiquitination. In the absence of NEM, the level of complex containing either p62/SIK2 or HDAC6/SIK2 was decreased. Whether the interaction between p62 or HDAC6 and SIK2 is direct or indirect remains to be investigated. Results from our lab have already shown that Sik2 is unlikely to play roles in the formation of aggresomes.
Ubiquitin–proteasome system and macroautophagy are two major pathways for protein degradation. Recent studies indicate that inhibition of the ubiquitin–proteasome system by proteasome inhibitors could induce macroautophagy through multiple pathways, including blocking mTOR complex 1 signaling, leading to accumulation of ubiquitinated proteins and activation of HDAC6 and reduced proteasomal degradation
of LC3. Treating cells with MG132 may activate ER stress response and autophagy.
Combining results from our Lab and my present results clearly establish SIK2 as important regulator for both processes. SIK2 may play roles for coordinating these events!
Disturbance of cellular energy homeostasis, such as starvation and nutrient depletion, results in AMPK activation, mTORC1 inactivation and ULK1 activation.
This complex interplay of signal transduction pathways explains, at least in part, the activation of autophagy during energy deprivation. Whether SIK2 could play a redundant role of AMPK remains a possibility. Studies indicate a connection between energy sensing and autophagy, but there are many further questions that remain to be answered. For example, autophagy can not be persistently activated or it will lead to cell death. Thus, if the cells are persistent in glucose starvation condition, how does autophagy be down regulated? Does another factor inactivate AMPK? What might be the role of SIK2 in this machinery? Understanding the mechanisms of autophagy attenuation is as crucial as understanding its induction.
Ubiquitin-proteasome system (UPS) and autophagy are the major routes for intracellular protein degradation. These pathways do communicate under certain circumstances. For example, mutated proteins such as polyglutamine-containing huntingtin (Htt106Q) or a1-antitrypsin often form aggregates that interfere with the proteasomal degradation and may rely on autophagy for their disposal. In fact, ER stress caused by tunicamycin or thapsigargin or proteasome inhibition by MG132 or lactacystin leads to the activation of autophagy. The pathways responsible for the induction of autophagy after ER stress are not entirely clear. One of the possible
pathways coordinating cross-talk between proteasome inhibition and autophagy activation may be mediated by JNK targeted phosphorylation of Bcl-2. Phosphorylated Bcl-2 dissociates from Beclin 1, thus allowing its initiation of autophagy. Other than the involvement of JNK, very little is known on the factors or signaling pathways that may coordinate the cross-talk of UPS and autophagy systems. In this paper, we uncovered the crucial roles of SIK2 in both UPS and autophagy. Our results indicate that the kinase activity of SIK2 is dispensable for its interaction with p97/VCP. These interactions are central for inclusion body or aggresome formation. However, the kinase activity of SIK2 is important for the ERAD process and autophagy. Several proteins including HDAC6, p62 and HSP90 are involved in the SIK2 and p97/VCP complex formation and dictating the autophagic degradation. In the HDAC6 and p62 complex, SIK2 kinase activity is critical for the resolution of autophagosomes. SIK2 kinase activity is also playing critical roles for the resolution of p97/VCP and HSP90 formed autophagosomes.
When HSP90 is inhibited by 17AAG, SIK2-WT is degraded along with reduced autophagosome formation while SIK2-KD remained in the insoluble fraction and failed to be degraded.
Another important finding of my research is that SIK2 is involved in regulating protein secretion through mediating autophagy. Recent publication showed that the TOR-autophagy spatial coupling compartment (TASCC) can regulate secretory phenotype (26). The spatial integration of rER-GA, autolysosomes, and mTOR could augment their functions, leading to the efficient synthesis of secretory and lysosomal proteins. Blocking autophagy would affect protein secretion. TASCC can be found in diverse contexts, it may represent a general mechanism for rapid protein turnover [26].
In this way, we observed the growth hormone secretion in growing cells, 293T. After
SIK2 knockdown or SIK2 KD overexpression, the growth hormone secretion was decreased. We also found that LC3- II was increased when we overexpressed SIK2 KD.
This suggests that SIK2 may influence growth hormone secretion mediated through autophagy regulation.
References
1. Katoh, Y., et al., Salt-inducible kinase (SIK) isoforms: their involvement in steroidogenesis and adipogenesis. Mol Cell Endocrinol, 2004. 217(1-2): p.
109-12.
2. Okamoto, M., H. Takemori, and Y. Katoh, Salt-inducible kinase in
steroidogenesis and adipogenesis. Trends Endocrinol Metab, 2004. 15(1): p.
21-6.
3. Horike, N., et al., Adipose-specific expression, phosphorylation of Ser794 in insulin receptor substrate-1, and activation in diabetic animals of salt-inducible kinase-2. J Biol Chem, 2003. 278(20): p. 18440-7.
4. Screaton, R.A., et al., The CREB coactivator TORC2 functions as a calcium- and cAMP-sensitive coincidence detector. Cell, 2004. 119(1): p. 61-74.
5. Katoh, Y., et al., Salt-inducible kinase-1 represses cAMP response
element-binding protein activity both in the nucleus and in the cytoplasm. Eur J Biochem, 2004. 271(21): p. 4307-19.
6. Du, J., et al., SIK2 can be activated by deprivation of nutrition and it inhibits expression of lipogenic genes in adipocytes. Obesity (Silver Spring), 2008. 16(3):
p. 531-8.
7. Kobayashi, T., A. Manno, and A. Kakizuka, Involvement of valosin-containing protein (VCP)/p97 in the formation and clearance of abnormal protein
aggregates. Genes Cells, 2007. 12(7): p. 889-901.
8. Kitami, M.I., et al., Dominant-negative effect of mutant valosin-containing protein in aggresome formation. FEBS Lett, 2006. 580(2): p. 474-8.
9. Ju, J.S., et al., Impaired protein aggregate handling and clearance underlie the pathogenesis of p97/VCP-associated disease. J Biol Chem, 2008. 283(44): p.
30289-99.
10. Kim, J., et al., AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat Cell Biol, 2011. 13(2): p. 132-41.
11. Egan, D.F., et al., Phosphorylation of ULK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy. Science, 2011. 331(6016): p.
456-61.
12. Inoki, K., T. Zhu, and K.L. Guan, TSC2 mediates cellular energy response to control cell growth and survival. Cell, 2003. 115(5): p. 577-90.
13. Gwinn, D.M., et al., AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol Cell, 2008. 30(2): p. 214-26.
14. Ichimura, Y., et al., Structural basis for sorting mechanism of p62 in selective autophagy. J Biol Chem, 2008. 283(33): p. 22847-57.
15. Pankiv, S., et al., p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J Biol Chem, 2007. 282(33): p. 24131-45.
16. Bjorkoy, G., et al., p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J Cell Biol, 2005. 171(4): p. 603-14.
17. Kuusisto, E., A. Salminen, and I. Alafuzoff, Ubiquitin-binding protein p62 is present in neuronal and glial inclusions in human tauopathies and
synucleinopathies. Neuroreport, 2001. 12(10): p. 2085-90.
18. Nagaoka, U., et al., Increased expression of p62 in expanded
polyglutamine-expressing cells and its association with polyglutamine inclusions. J Neurochem, 2004. 91(1): p. 57-68.
19. Kabeya, Y., et al., LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J, 2000. 19(21): p. 5720-8.
20. Seibenhener, M.L., et al., Sequestosome 1/p62 is a polyubiquitin chain binding protein involved in ubiquitin proteasome degradation. Mol Cell Biol, 2004.
24(18): p. 8055-68.
21. Geetha, T. and M.W. Wooten, Structure and functional properties of the
ubiquitin binding protein p62. FEBS Lett, 2002. 512(1-3): p. 19-24.
22. Ciani, B., et al., Structure of the ubiquitin-associated domain of p62 (SQSTM1) and implications for mutations that cause Paget's disease of bone. J Biol Chem, 2003. 278(39): p. 37409-12.
23. Kawaguchi, Y., et al., The deacetylase HDAC6 regulates aggresome formation and cell viability in response to misfolded protein stress. Cell, 2003. 115(6): p.
727-38.
24. Iwata, A., et al., HDAC6 and microtubules are required for autophagic
degradation of aggregated huntingtin. J Biol Chem, 2005. 280(48): p. 40282-92.
25. Boyault, C., et al., HDAC6-p97/VCP controlled polyubiquitin chain turnover.
EMBO J, 2006. 25(14): p. 3357-66.
26. Narita, M., et al., Spatial coupling of mTOR and autophagy augments secretory phenotypes. Science, 2011. 332(6032): p. 966-70.
27. Duran, J.M., et al., Unconventional secretion of Acb1 is mediated by autophagosomes. J Cell Biol, 2010. 188(4): p. 527-36.
28. Nickel, W. and C. Rabouille, Mechanisms of regulated unconventional protein secretion. Nat Rev Mol Cell Biol, 2009. 10(2): p. 148-55.
29. Nickel, W., Pathways of unconventional protein secretion. Curr Opin
Biotechnol, 2010. 21(5): p. 621-6.
30. Rivera, V.M., et al., Regulation of protein secretion through controlled
aggregation in the endoplasmic reticulum. Science, 2000. 287(5454): p. 826-30.
31. Reed, N.A., et al., Microtubule acetylation promotes kinesin-1 binding and transport. Curr Biol, 2006. 16(21): p. 2166-72.
32. Dompierre, J.P., et al., Histone deacetylase 6 inhibition compensates for the transport deficit in Huntington's disease by increasing tubulin acetylation. J Neurosci, 2007. 27(13): p. 3571-83.
33. Oliver, F.J., et al., Importance of poly(ADP-ribose) polymerase and its cleavage in apoptosis. Lesson from an uncleavable mutant. J Biol Chem, 1998. 273(50): p.
33533-9.
34. Tewari, M., et al., Yama/CPP32 beta, a mammalian homolog of CED-3, is a CrmA-inhibitable protease that cleaves the death substrate poly(ADP-ribose) polymerase. Cell, 1995. 81(5): p. 801-9.
List of figures
A.
B. input
Fig. 1 SIK2 may form complex with p62 and HDAC6
(A) 293T cells were co-transfected with Flag-p62 (F-p62) or Flag-HDAC6 (F-HDAC6) and His-SIK2 (H-SIK2). 40 hrs after transfection, cells were harvested. The cell lysates were immunoprecipitated with M2 beads in the presence or absence of NEM (25mM), and probed with anti-SIK2 and anti-flag antibody. (B) The protein level of SIK2, flag-p62 and flag-HDAC6 in the cell lysates were probed with mAb to SIK2 (clone 15G10) and anti-flag antibody.
A.
B. C.
Fig. 2 p62 and SIK2-containing complex formation appeared to be enhanced both in the soluble and insoluble fractions when cells were treated with MG132.
Flag-p62 and His-SIK2 were co-transfected into 293T cells and treated with EtOH or Mg132(5μM) for 16 hrs. The harvested cells were separated to soluble (A) and insoluble fractions (B). (A) The soluble fraction was used for immunoprecipitation with M2 beads in the presence of NEM (25mM) and probed with anti-ubiquitin, Flag and SIK2 mAb (15G10). (B)(C)The protein level of Flag-p62, ubiquitin, LC3 and SIK2 in insoluble or soluble fraction was assessed by western blot.
A.
B.
C.
Fig. 3 Complex formation between HDAC6 and SIK2
WT flag-HDAC6 (W) or catalytic dead flag-HDAC6 (CD) and His-SIK2 or pcDNA3 were co-transfected into 293T cells. 24 hrs after transfection, cells were treated with EtOH or MG132 (5μM) for 16 hrs. The harvested cells were separated to soluble (A)(C) and insoluble fractions (B). (A) The soluble fraction was immunoprecipitated with M2 beads in the presence of NEM (25mM) and probed with anti-ubiquitin and SIK2 (15G10) antibody. (B)(C) The protein level of Flag-p62, ubiquitin and SIK2 in insoluble or soluble fraction was assessed by western blot.
SIK2 (long exposure A.
B.
SIK2 (long exposure Sup
Pellet
Fig. 4 Both kinase activity of SIK2 and chaperone activity of Hsp90 is important for processing of autophagosomes containing SIK2, p97/VCP and Hsp90
WT flag-His-p97/VCP(F-H-p97W) or flag-His-p97/VCP S770A(F-H-p97A) and Flag-SIK2 or pCMV2B were co-transfected into 293T cells. 24 hrs after transfection, cells were treated with 17DMAG (1μM) for 16 hrs. The harvested cells were separated to soluble (A) and insoluble fractions (B). (A) The soluble fraction was probed with anti-SIK2 (15G10), actin, Hsp27 and Hsp70 antibody. (B) The insoluble fraction was probed with anti-SIK2 (15G10), p97(4G9), Hsp27 and Hsp70 antibody.
A. B.
C. D.
E. F.
LC3 LC3
Fig. 5 Autophagy is involved in regulation of ligand-induced protein secretion by SIK2
(A ) pC4S1-FM4-FCS-hGH (hGH) was transfected into 293T cells. 24 hrs after transfection, the cells were treated with DMSO, bafilomycin (BFA) (0.2 μM) or compound C (10μM) for 15hrs, changed to serum-free medium and treated with DMSO or AP21998 (4μM) for 1 hr. The serum-free culture medium was collected and subjected to TCA precipitation. The level of growth hormone (GH) in the precipitates was determined by immunoblotting with rabbit anti-GH. (B) The cell-free extracts were used for probing SIK2, LC3 and actin by western blot analysis. (C) pC4S1-FM4-FCS-hGH (hGH) and shLUC, shSIK2, Flag-SIK2-WT (F-SIK2-WT), Flag-SIK2-KD (F-SIK2-KD) or pCMV2B vector were cotransfected into 293T cells. 40 hrs after transfection, the cells were changed to serum-free medium and treated with AP21998 (4μM) for 1 hr. The culture medium was subjected to TCA precipitation and the level of growth hormone was determined by immunoblotting. (D) The harvested cells from (C) were used for western blot analysis. Primary antibodies against SIK2 and β-actin were used. (E) pC4S1-FM4-FCS-hGH (hGH) and Flag-SIK2-WT (F-SIK2-WT), Flag-SIK2-KD (F-SIK2-KD) were cotransfected into 293T cells. 24 hrs after transfection, the cells were treated with DMSO, BFA (0.2 μM), or compound c(10μM) for 15hrs, changed to serum-free medium and treated with AP21998 (4μM) for 1 hr. The culture medium was subjected to TCA precipitation and the level of growth hormone was determined by immunoblotting. (F) The harvested cells from (E) were used for western blot analysis. Primary antibodies against SIK2, LC3 and β-actin were used.
A.
B.
shLUC shSIK2 RATIO
EtOH 47.2 / 9.87 = 4.8 45.2 / 8.6 = 5.25 5.25 / 4.8 = 1.1 MG132 48.5 / 12 = 4 75.1 / 7.2 = 10.4 10.4 / 4 = 2.6
C.
D.
siLUC siSIK2 SIK2 WT SIK2 KD PARP-1 fragment 337.07590 2940.555 1245.3063 2156.037
siSIK2/siLUC= 8.723719 KD/WT= 1.731331
Fig. 6 SIK2 is important for the cell survival under stress conditions
(A) 24 hrs after transfection shLUC or shSIK2, 293T cells were treated with EtOH or MG132(10μM) for 16 hrs. Cells were then isolated and binding with annexin V-FITC and analyzed by flow cytometry. GFP and annexin V-positive cells were counted.
Quantification data was shown in (B). (C) 24 hrs after transfection with shLUC, shSIK2, flag-SIK2 WT,or flag-SIK2 KD, 293T cells were treated with EtOH or MG132 (10μM) for 16 hrs. The harvested cells were probed with SIK2, PARP-1, and β-actin antibodies, respectively. Quantification data was shown (D).
PARP-1