行政院國家科學委員會專題研究計畫 期末報告
細胞核基質蛋白 Matrin 3 (MATR3)在人類 D 型肝炎病毒 RNA 的合成過程中的重要性
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 101-2320-B-040-006-
執 行 期 間 : 101 年 08 月 01 日至 102 年 07 月 31 日 執 行 單 位 : 中山醫學大學醫學研究所
計 畫 主 持 人 : 許國堂
計畫參與人員: 博士班研究生-兼任助理人員:丘翎燕
公 開 資 訊 : 本計畫涉及專利或其他智慧財產權,2 年後可公開查詢
中 華 民 國 102 年 10 月 17 日
中 文 摘 要 : 人類的 D 型肝炎病毒(HDV)具有一個環狀單股的核酸(RNA)做 為它的基因體。因此,使用其 RNA 為鑄模來做為合成新基因 體之用。到目前為止,HDV 的基因體中,只有發現一個基 因。這個基因的產物就是 D 型肝炎病毒的抗原(HDAg)。細胞 核酸聚合酶的 pol II 可能是主要的核酸聚合酶用來轉錄 HDAg-mRNA,可是 pol II 只能使用 DNA 為鑄模,如何讓它扭 轉成以 RNA 為鑄模的專一性轉錄,還未有任何合理的解釋。
假如 pol II 可以辨識 HDV 的 RNA 並做為鑄模進行轉錄工作,
則我們可以合理推測其中必須要有中間者才可以把 pol II 的 工作羣和 HDV 的 RNA 銜接起來。這樣或許是 pol II 可以把 HDV 的 RNA 拿來進行轉錄工作的一種步驟。所以我們在此計 畫中推論 MATR3 應該是經過 D 型肝炎病毒的抗原來參與轉錄 HDAg-mRNA 的重要因素。為了找出細胞內可能參與 HDAg 基因 轉錄的蛋白質,我們已經建立了會表現 HDAg 的 Huh7 細胞株 來研究會被 HDAg 向上調控的蛋白質,並且經過蛋白質體學之 二維電泳分析和液相層析串聯質譜的分析之後,找到了在細 胞核液中的 Matrin3。
Matrin3 是細高度保留的細胞核內基質蛋白,和細胞膜內膜 可以形成細胞核內骨架來參與染色質編排、DNA 修復、轉錄 和 RNA 修飾。Matrin3 具有核定位訊息序列(NLS),兩個鋅指 DNA 結合區(Zinc finger)預計會和 DNA 結合。同時它也有兩 組 RNA 辨識模體(RRM)預計會和 RNA 結合。因為 Matrin3 是 pol II 的工作羣而且會和細胞核內的修飾過的 RNA 結合,所 以在此部份之後續計畫,我們假設 pol II 的工作羣中的 Matrin3 是經由 HDAg 才能用 HDV 的 RNA 拿來進行轉錄工作。
我們並且從微矩陣基因晶片技術的分析也看到 MATR3 和 p54nrb/NonO 的表達量還是有增加的。也觀察到在表現 HDAg 的 Huh7 細胞株中 Matrin3 和它的伙伴 p54nrb/NonO 的表達量 是有增加的。我們將使用基因高度表達和 RNA 干擾抑制方法 對 Matrin3 在複製 HDV 時的角色進行研究。我們已經把 MATR3 和 p54nrb/NonO 的表達載體建立起來。因為我們已經 建立了可以完全不用 DNA,可以很接近 HDV 在細胞內複製的 狀態的模式進行實驗,所以我們可以專一性地辨識細胞的 HDV 核酸聚合酶(RdRP)的工作羣。是以我們將有機會瞭解到 pol II 工作羣如何和 HDV RNA 進行作用,這對於醫學和生 物學上都可以有相當的影響性。
中文關鍵詞: D 型肝炎病毒; D 型肝炎病毒抗原; 蛋白質體學; Matrin 3; RNA 為鑄模的核酸聚合酶; 微矩陣基因晶片
英 文 摘 要 : Human hepatitis delta virus (HDV) has a single-
stranded circular RNA genome that replicates by RNA- directed RNA synthesis. The virus encodes only a single protein product, the hepatitis delta antigen (HDAg). It has been proposed that transcription of HDAg-mRNA is mediated by RNA polymerase II, a
cellular DNA-directed RNA polymerase. Due to the lack of natural infection for in vivo model and a reliable in vitro transcription assay system, the hunt for the human cellular HDV RNA-directed RNA polymerase (RdRP) is hampered. If genomic of HDV RNA can be recognized by the DNA-dependent pol II, there must be a switch exist that connects the pol II transcriptional
machinery and HDV RNA to synthesize mRNA of HDAg. To identify cellular proteins that involved in HDAg transcription, we have analyzed the nuclear lysate of Huh-7 stable clones that expressing HDAg by
proteomics on two-dimensional polyacrylamide gel electrophoresis to detect the HDAg-upregulated
proteins. From these experiments, Matrin3 was further identified by LC-MS/MS analysis.
Matrin3 is a highly conserved, inner nuclear matrix protein of 125 kDa. Nuclear matrix proteins bound to the inner nuclear membrane form a skeletal nuclear framework with roles in chromatin organization, DNA repair, transcription, and RNA processing. Matrin3 contains a bipartite nuclear localization signal (NLS), two zinc finger domains predicted to bind DNA, and two RNA recognition motifs (RRM). We further postulated that Matrin3 may connect the DNA-dependent pol II machinery via HDAg to synthesize the HDAg- mRNA. We also applied DNA microarray technology to analyze gene expression altered by HDAg and found both Matrin3 and p54nrb/NonO, were upregulated. The expressions of Matrin3 and its partner, p54nrb/NonO, were upregulated in small-HDAg expressing Huh7 cells that examined by immunoblotting.
To characterize the role of Matrin3 plays in HDV RNA synthesis, we went on to study the essential role of Matrin3 in HDV replication by gene overexpression and RNA interference. We have cloned and constructed the expression vectors of Matrin3 and p54nrb/NonO. Since we already established a cDNA-free HDV replication
system in cellular model, we are able to specifically analyze the cellular RdRP machinery from human cell that responsible for HDV RNA synthesis. How to switch the pol II machinery from DNA-dependence to recognize HDV RNA as the template for RNA synthesis is an
important issue in the fields of biology and medicine.
英文關鍵詞: Hepatitis delta virus; Hepatitis delta antigen;
Proteomics; Matrin 3; RNA-directed RNA polymerase;
DNA microarray
□期中進度報告 行政院國家科學委員會補助專題研究計
畫 V 期末報告
(計畫名稱)
細胞核基質蛋白 Matrin 3 (MATR3)在人類 D 型肝炎病毒 RNA 的合成過程中的重要 性
The role of an inner nuclear matrix protein Matrin 3 (MATR3) in human hepatitis delta virus RNA synthesis
計畫類別:v 個別型計畫 □整合型計畫 計畫編號:NSC 101-2320-B-040-006-
執行期間: 101 年 08 月 01 日至 102 年 07 月 31 日 執行機構及系所:中山醫學大學醫學研究所
計畫主持人: 許國堂 共同主持人:
計畫參與人員: 兼任助理人員:丘翎燕
本計畫除繳交成果報告外,另含下列出國報告,共 ___ 份:
□移地研究心得報告
□出席國際學術會議心得報告
□國際合作研究計畫國外研究報告
處理方式:除列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年 v 二年後可公開查詢 中 華 民 國 102 年 10 月 30 日
I 附件一
中文摘要及關鍵詞 (keywords):
D 型肝炎病毒; D 型肝炎病毒抗原; 蛋白質體學; Matrin 3; RNA 為鑄模的核酸聚合酶; 微矩陣基 因晶片
人類的 D 型肝炎病毒(HDV)具有一個環狀單股的核酸(RNA)做為它的基因體。因此,使用 其 RNA 為鑄模來做為合成新基因體之用。到目前為止,HDV 的基因體中,只有發現一個基因。
這個基因的產物就是 D 型肝炎病毒的抗原(HDAg)。細胞核酸聚合酶的 pol II 可能是主要的核酸 聚合酶用來轉錄 HDAg-mRNA,可是 pol II 只能使用 DNA 為鑄模,如何讓它扭轉成以 RNA 為 鑄模的專一性轉錄,還未有任何合理的解釋。假如 pol II 可以辨識 HDV 的 RNA 並做為鑄模進 行轉錄工作,則我們可以合理推測其中必須要有中間者才可以把 pol II 的工作羣和 HDV 的 RNA 銜接起來。這樣或許是 pol II 可以把 HDV 的 RNA 拿來進行轉錄工作的一種步驟。所以我們在 此計畫中推論 MATR3 應該是經過 D 型肝炎病毒的抗原來參與轉錄 HDAg-mRNA 的重要因素。
為了找出細胞內可能參與 HDAg 基因轉錄的蛋白質,我們已經建立了會表現 HDAg 的 Huh7 細 胞株來研究會被 HDAg 向上調控的蛋白質,並且經過蛋白質體學之二維電泳分析和液相層析串 聯質譜的分析之後,找到了在細胞核液中的 Matrin3。
Matrin3 是細高度保留的細胞核內基質蛋白,和細胞膜內膜可以形成細胞核內骨架來參與染 色質編排、DNA 修復、轉錄和 RNA 修飾。Matrin3 具有核定位訊息序列(NLS),兩個鋅指 DNA 結合區(Zinc finger)預計會和 DNA 結合。同時它也有兩組 RNA 辨識模體(RRM)預計會和 RNA 結合。因為 Matrin3 是 pol II 的工作羣而且會和細胞核內的修飾過的 RNA 結合,所以在此部份 之後續計畫,我們假設 pol II 的工作羣中的 Matrin3 是經由 HDAg 才能用 HDV 的 RNA 拿來進 行轉錄工作。
我們並且從微矩陣基因晶片技術的分析也看到 MATR3 和 p54nrb/NonO 的表達量還是有增加
的。也觀察到在表現 HDAg 的 Huh7 細胞株中 Matrin3 和它的伙伴 p54nrb/NonO 的表達量是有增 加的。我們將使用基因高度表達和 RNA 干擾抑制方法對 Matrin3 在複製 HDV 時的角色進行研
究。我們已經把 MATR3 和 p54nrb/NonO 的表達載體建立起來。因為我們已經建立了可以完全不
用 DNA,可以很接近 HDV 在細胞內複製的狀態的模式進行實驗,所以我們可以專一性地辨識 細胞的 HDV 核酸聚合酶(RdRP)的工作羣。是以我們將有機會瞭解到 pol II 工作羣如何和 HDV RNA 進行作用,這對於醫學和生物學上都可以有相當的影響性。
II
英文摘要 and Key words: Hepatitis delta virus; Hepatitis delta antigen; Proteomics; Matrin 3;
RNA-directed RNA polymerase; DNA microarray
Human hepatitis delta virus (HDV) has a single-stranded circular RNA genome that replicates by RNA-directed RNA synthesis. The virus encodes only a single protein product, the hepatitis delta antigen (HDAg). It has been proposed that transcription of HDAg-mRNA is mediated by RNA polymerase II, a cellular DNA-directed RNA polymerase. Due to the lack of natural infection for in vivo model and a reliable in vitro transcription assay system, the hunt for the human cellular HDV RNA-directed RNA polymerase (RdRP) is hampered. If genomic of HDV RNA can be recognized by the DNA-dependent pol II, there must be a switch exist that connects the pol II transcriptional machinery and HDV RNA to synthesize mRNA of HDAg. To identify cellular proteins that involved in HDAg transcription, we have analyzed the nuclear lysate of Huh-7 stable clones that expressing HDAg by proteomics on two-dimensional polyacrylamide gel electrophoresis to detect the HDAg-upregulated proteins. From these experiments, Matrin3 was further identified by LC-MS/MS analysis.
Matrin3 is a highly conserved, inner nuclear matrix protein of 125 kDa. Nuclear matrix proteins bound to the inner nuclear membrane form a skeletal nuclear framework with roles in chromatin organization, DNA repair, transcription, and RNA processing. Matrin3 contains a bipartite nuclear localization signal (NLS), two zinc finger domains predicted to bind DNA, and two RNA recognition motifs (RRM). We further postulated that Matrin3 may connect the DNA-dependent pol II machinery via HDAg to synthesize the HDAg-mRNA. We also applied DNA microarray technology to analyze gene expression altered by HDAg and found both Matrin3 and p54nrb/NonO, were upregulated. The expressions of Matrin3 and its partner, p54nrb/NonO, were upregulated in small-HDAg expressing Huh7 cells that examined by immunoblotting.
To characterize the role of Matrin3 plays in HDV RNA synthesis, we went on to study the essential role of Matrin3 in HDV replication by gene overexpression and RNA interference. We have cloned and constructed the expression vectors of Matrin3 and p54nrb/NonO. Since we already established a cDNA-free HDV replication system in cellular model, we are able to specifically analyze the cellular RdRP machinery from human cell that responsible for HDV RNA synthesis. How to switch the pol II machinery from DNA-dependence to recognize HDV RNA as the template for RNA synthesis is an important issue in the fields of biology and medicine.
III
前言:
Background of Hepatitis delta virus (HDV) HDV life
HDV has a circular, single-stranded RNA genome of 1,700 nucleotides, with about 70%
self-complementarily (Kos et al., 1986; Kuo et al., 1988a; Makino et al., 1987; Wang et al., 1986).
This genome is thought to be replicated by the host RNA polymerase II through a double- rolling-circle mechanism (Chen et al., 1986; Fu & Taylor, 1993; Lee et al., 1993). Replication of HDV genome is independent to HBV infection (Chen et al., 1986). Once enter the cell, HDV can produce linear multimers of both genomic and antigenomic RNAs, both of which posses a self-cleaving activity that processes the RNAs to unit length (Kuo et al., 1988b; Sharmeen et al., 1988; Wu et al., 1989). There are two detectable HDV antigens (HDAg) encoded by a single open reading frame on the antigenomic strand (Bergmann & Gerin, 1986; Bonino et al., 1986; Pohl et al., 1987; Weiner et al., 1988). The large (L-HDAg) is a result of RNA editing which extends the protein coding sequence of the small antigen (S-HDAg) by 19 residues (Luo et al., 1990).
HDV antigens
The S-HDAg is essential for viral replication and is proposed to facilitate the interactions between the cellular transcription machinery and the viral template (Kuo et al., 1989; Lin et al., 1990).
HDAg inhibits cellular RNA polymerase II but not the pol I and pol III in vivo with a CAT reporter assay and in vitro with the purified recombinant HDAg have been shown (Lo et al., 1998), suggesting that mass amount of HDAg capable interact with pol II specific transcriptional component. The central domain of S-HDAg, which is involved in binding to the viral genome, contains two arginine-rich motifs bracketing a leucine zipper (Lee et al., 1993). The nuclear import of HDV capsid protein may be the first biological function of S-HDAg that leads to the replication of HDV RNA in the nuclei of infected cells (Chou et al., 1998). L-HDAg appears later in infection, suppresses replication and initiates viral packaging (Chang et al., 1991; Chao et al., 1990). We have reported that HDV replication was associated with epigenetic regulation resulted in histone hyperacetylation and clusterin induction. Thus, both isoforms of the HDAg may act as co-activators of cellular gene expression (Liao et al., 2009).
HDV-RNAs
Three major stable RNA species are thought to be involved in the replication of the HDV RNA genome. (a) The genomic 1,679-nucleotide circular single-stranded RNA. (b) Its exact complement, the antigenome, and (c) an 800-nucleotide polyadenylated RNA that is least abundant for the mRNA of HDAg (Gudima et al., 1999; Perrotta & Been, 1990; Taylor, 1990). It has been proposed that the 800-nt mRNA is continuing synthesized throughout HDV replication that is not suppressed by the translated HDAg as previous reported (Modahl & Lai, 1998).
HDV RNA editing
During HDV genome replication in the nucleus, some of the new antigenomic RNAs get edited by an enzyme of the class known as adenosine deaminase acting on RNA (ADAR). The small form of ADAR1, is considered to be the specific enzyme that acts on HDV RNA (Wong & Lazinski, 2002). It changes adenosine to inosine. Editing occurs specifically on adenosine 1012 (the amber/W site) of the
antigenome (Casey & Gerin, 1995) and changes the UAG (amber) stop codon of S-HDAg to a UGG (Trp) codon, which allows for the C-terminal addition of the 19 to 20 amino acids unique to L-HDAg . This edited change is processing by subsequent rounds of RNA-directed RNA synthesis, as adenosine to guanosine. Typically, 20-50% of antigenomic RNA was edited in vitro and in vivo (Polson et al., 1996) and is suppressed by S-HDAg (Polson et al., 1998).
Systems have been used to replicate HDV in cell lines
HDV replication products have been obtained by several transfection approaches that bypass the processes of virus attachment and penetration: (A) Transfection with a cDNA plasmid containing three tandem head-to-tail inserts of the HDV genome under the control of SV-40 promoter (Kuo et al., 1989) (B) Transfection of in vitro synthesized genomic RNA into S-HDAg stable expressed fibroblasts (Glenn et al., 1990). (C) Co-transfection of in vitro synthesized HDV RNA and mRNA of S-HDAg into Huh-7 cells (Modahl & Lai, 1998). (D) Transfection of HDV RNA and recombinant S-HDAg ribonucleoprotein complex (RNP) into Huh-7 cells (Dingle et al., 1998; Sheu & Lai, 2000). All of the four approaches require the presence of the S-HDAg for HDV replication. We have established the system of (B) that replicates in vitro synthesized genomic RNA in S-HDAg stable expressing Huh-7 cells (Huh-sm). This RNA transfection system is considered as the most reliable system to study the HDV replication (Liao et al., 2012).
Enzymology of RNA-directed RNA replication of HDV
Replication of HDV genomic, antigenomic and message RNAs apparently are mediated by distinct cellular RNA polymerases. The mRNA transcription is inhibited by a low concentration of α-amanitin and could berescued partially by an α-amanitin-resistant mutant pol II. Therefore, pol II is supposed to synthesize mRNA of HDAg from the HDV RNA genomic template (Modahl et al., 2000).
How pol II transcription machineries are converted from DNA-directed synthesis to RNA-directed synthesis is not clear. Our proposal in here is focus at this issue. The synthesis of the anti-genomic HDV RNA was not affected by α-amanitin, therefore, pol II may not responsible for the antigenomic RNA synthesis (Modahl et al., 2000). Moreover, the synthesis of genomic RNA was inhibited by L-HDAg, whereas, anti-genomic RNA synthesis was not affected (Modahl & Lai, 2000). We have shown that only genomic RNA associated with S-HDAg could be used as template for replication, anti-genomic RNA and S-HDAg RNP could not initiate replication (Sheu & Lai, 2000). All these evidences point to that replication and transcription of HDV are mediated by distinct cellular polymerases. Recently, it has been reported that human RNA polymerases II (Greco-Stewart et al., 2007), I and III (Greco-Stewart et al., 2009) interact with the terminal stem-loop region. The RNA pol II forms a preinitiation complex on this region (Abrahem & Pelchat, 2008). However, whether the initiation site of 1.7-kb antigenomic or genomic HDV RNA synthesis is identical to that of 0.8-kb mRNA transcription is not known. We demonstrated the nucleotide positions 1582 to 1683 (TR-P1) that located at the terminal stem-loop region are substantial for the maximal promoter activity.
Mutations of TR-P1 to P1-m5 completely block the HDV RNA synthesis in Huh-sm cells (Liao et al., 2012). TR-P1 region most likely is the authentic HDV RNA promoter for mRNA transcription.
Study the HDAg regulated proteins by proteomic and immunoprecipitation methods
To identify the changes in the expression profiles of host proteins caused by the presence of the different virus components, Huh7 cells have been transiently transfected with plasmids coding
exclusively for the S-HDAg, L-HDAg, genomic RNA, and antigenomic gRNA, respectively. Total protein extracts were separated by 2-DE and differentially expressed spots were identified by MALDI-TOF followed by database searching (Mota et al., 2008). Although a total of 32 proteins were identified using this approach, whether or not they interact with HDAg and/or affect HDV metabolism was not addressed. Similar approach was applied to Huh7-D12 cells that stably expressing HDV RNPs (Mota et al., 2009) and 23 differentially expressed proteins were identified, of which 15 were down regulated and 8 up regulated in Huh7-D12 cells. A combined proteomic-RNAi screen for host proteins interacting with S-HDAg also has been reported on HEK293 cells that stably express a functional Flag-S-HDAg (Cao et al., 2009). By anti-Flag immunopurification and mass spectrometry analysis, many proteins were identified. Total of 65 proteins were selected to look for whether they would also affect the accumulation of HDV RNA following siRNA knockdown. The results showed most of the proteins tested could affect RNA accumulation.
An alternative approach to search for HDV host factors employed mass spectrometry analysis following UV crosslinking of HeLa nuclear proteins with an RNA corresponding to the right terminal stem–loop domain of HDV genomic RNA (Greco-Stewart et al., 2006). This identified the polypyrimidine tract-binding protein-associated splicing factor (PSF) as a host factor bound to HDV RNA, although PSF functional involvement during physiological HDV replication remains unclear.
Additional protein interactions have also been reported (Sikora et al., 2009). The proteins of eEF1A1, p54nrb/NonO, HnRNP-L, GAPDH and ASF/SF2 were co-immunoprecipitated with the right terminal stem-loop domain of HDV genomic RNA in vitro. Those data indicate that HDV RNA associates with RNA-processing pathways and translation machinery during its replication provides new insights into HDV biology. There have been additional reports of host proteins that bind to either HDAg or HDV RNA. PML (Li et al., 2006), nucleolin (Lee et al., 1998), B23 (Huang et al., 2001), and transcription factor YY1 (Huang et al., 2008) have been shown to interact with HDAg and were suggested to affect HDV replication. The Pol I specific factor SL1 (Li et al., 2006) and the negative transcription elongation factor DSIF (Yamaguchi et al., 2001) have also been demonstrated to interact with HDAg, but their effect on HDV replication in vivo is not clear. A double-stranded RNA-activated kinase, PKR, has been shown to bind to S-HDAg (Chen et al., 2002) and HDV RNA (Robertson et al., 1996) (Circle et al., 1997), and affect HDV replication (Chen et al., 2002). Finally, GAPDH was reported to bind to HDV RNA (Lin et al., 2000) and ADAR1 was found to edit HDV antigenomic RNA (Wong &
Lazinski, 2002).
Background of MATR3, PSF and p54nrb/NonO Matrin3
Nuclear matrix is a major component of the cell nucleus (Berezney et al., 1995) and is composed of over 200 proteins (Fey & Penman, 1988). The major proteins of the nuclear matrix include lamins A, B and C, which can form a meshwork within the nucleoplasm (Inagaki et al., 1996): lamin B1, which associates with chromatin at the matrix attachment regions (Inagaki et al., 1996); the nucleolar protein B-23; core heterogeneous nuclear RNPs, and the nuclear matrins (Berezney et al., 1995). The nuclear matrins are components of the internal nuclear matrix and include matrins 1, 3, 4, D, E, F, G, 12 and 13 (Berezney et al., 1995). Of these, matrins D, E, F, G, and 4 have been identified as DNA-binding proteins (Hakes & Berezney, 1991). Matrin3 is a highly conserved, inner nuclear matrix protein of 125
kDa. The primary structure deduced for Matrin3 predicts a bipartite nuclear localization signal (NLS), two DNA binding domains with C2H2-type zinc finger domain and two tandem RNA recognition motifs. It has been implicated in the processing of RNAs and retention of hyper-edited RNA within the nucleus (Zhang & Carmichael, 2001). Moreover, Matrin3 has nuclear localization signals and several phosphorylation sites for tyrosine or serine/threonine kinases (Beausoleil et al., 2004). Though its activity and mode of action are still unclear, it appears that Matrin3 serves as a scaffolding molecule that interacts with a number of unique nuclear localized proteins (Malyavantham et al., 2008; Zeitz et al., 2009). A majority of these proteins are involved in RNA metabolism and chromatin remodeling while others function in protein translation, DNA replication/repair and apoptosis.
Polypyrimidine tract-binding protein (PTB)-associated splicing factor (PSF) and p54nrb/NonO PSF, a 100 kDa polypeptide, was identified and characterized in a complex with PTB (Patton et al., 1993). p54nrb (human) and NonO (mouse) are highly homologous to the C-terminus of PSF (Dong et al., 1993; Yang et al., 1993). Proteomics have identified PSF and p54nrb/NonO in the nucleolus (Andersen et al., 2002) and in association with the nuclear membrane (Dreger et al., 2001). Matrin3 together with the proteins PSF and p54nrb/NonO were implicated in nuclear retention of hyper-edited RNA, which prevents the translation of such RNA (Zhang & Carmichael, 2001). This process occurs in nuclear bodies called paraspeckles that contain PSF and p54nrb/NonO proteins (Bond & Fox, 2009).
PSF and p54nrb/NonO form a heterodimer and are involved in various aspects of RNA and DNA metabolism (Shav-Tal & Zipori, 2002) such as transcription (Buxade et al., 2008; Dong et al., 2007;
Hata et al., 2008), pre-mRNA 3' processing (Rosonina et al., 2005), transcription termination (Kaneko et al., 2007) and mRNA splicing (Ito et al., 2008; Kameoka et al., 2004). Both PSF and p54nrb/NonO were identified in a biochemical screen for DNA end-rejoining proteins (Bladen et al., 2005).
Furthermore, they were shown to facilitate in vitro the DNA binding capability of the KU70/KU80 heterodimer, a component of DNA-PK holoenzyme; and PSF was shown to bind RAD51 and to modulate RAD51-mediated homologous pairing and strand exchange.
方法、結果與討論 (含結論與建議)
This research plan: Investigation of the essential role of Matrin3 in HDV replication by gene overexpression and RNA interference.
Methods:
(1) Proteomics analysis of HDAgs upregulated proteins in Huh7 cells stably expressing S-HDAg or L-HDAg
Results: Matrin3 was further identified by LC-MS/MS analysis.
Fig.1 Proteomic analysis of HDAg-regulated cellular proteins by 2D-PAGE and LC-MS/MS. The Huh7 cells that (A) stably expressing L-HDAg (Huh-LAg) and (B) stably expressing S-HDAg (Huh-SAg) were harvested and the nuclear lysates (2 mg) were obtained for proteomic analysis. The upregulated proteins that observed in gels were sliced for LC-MS/MS. Numbers of proteins that were identified are 1: Matrin3; 2: Heat shock protein 27; 3: Glyceraldehydes-3-phosphate dehydrogenase (GAPDH); 4: HnRNP-L. The 1’and 4’indicate similar spots that were not used for analysis and presumed to be the identified one.
Methods:
(2) Analysis of gene expression altered by L-HDAg or S-HDAg with DNA microarray technology.
Results: We found transcription of Matrin3 and p54nrb/NonO was upregulated.
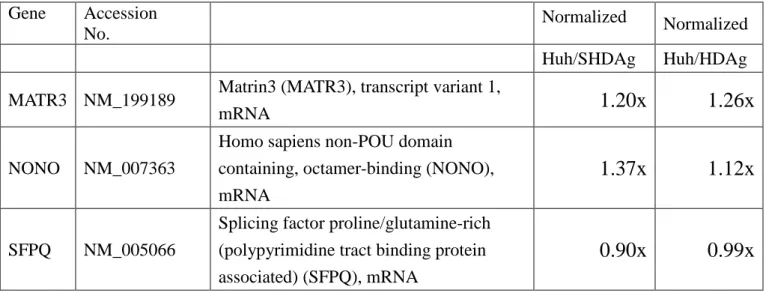
Table 1: The comparison of MATR3, p54nrb/NonO and SFPQ transcription levels in Huh7 cells expressing S-HDAg or L-HDAg.
Gene Accession No.
Normalized Normalized Huh/SHDAg Huh/HDAg MATR3 NM_199189 Matrin3 (MATR3), transcript variant 1,
mRNA
1.20x 1.26x
NONO NM_007363
Homo sapiens non-POU domain containing, octamer-binding (NONO), mRNA
1.37x 1.12x
SFPQ NM_005066
Splicing factor proline/glutamine-rich (polypyrimidine tract binding protein associated) (SFPQ), mRNA
0.90x 0.99x
Methods:
(3) Examination of the Matrin3 and p54nrb/NonO expressions in Huh7 cells expressing S-HDAg.
Results: The expressions of Matrin3 and p54nrb/NonO, were upregulated in S-HDAg expressing Huh7 cells
Figure 2. The expression of MAtrin3 and p54nrb/NonO were upregulated in Huh7 cell expressing S-HDAg (Huh7-SHDAg) cells that examined by immunoblotting.
Methods:
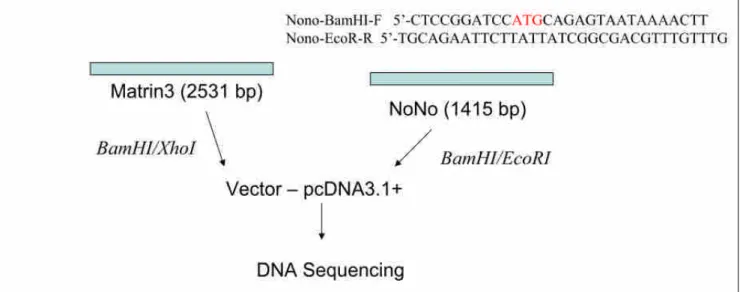
(4) Construction and transfection of Matrin 3 and p54nrb/NonO expression plasmids into Huh7 cells followed by Western blots analysis
Figure 3. To study the function of Matrin3 and p54nrb/NonO, we first construct the expression clone of Matrin3 and p54nrb/NonO into expression vector (pcDNA3.1+).
Results: DNA coding Matrin3 was obtained by reverse transcription of total RNA isolated from Huh7 cells, by the use of SuperScript III Reverse Transcriptase with primers: MATR3-F BamHI and
MATR3-RXhoI. PCR product is cloned into pcDNA3.1+ using XhoI and BamHI restriction sites.
For cloning p54nrb/NonO gene, primers: Nono-BamHI-F and Nono-EcoRI-R were used. The obtained pcDNA-Matrin3 and NoNo is propagated in Escherichia coli XL-10, purified with Plasmid Miniprep Kit, and sequenced to confirm the open reading frame. We have successfully obtained Matrin3 and p54nrb/NonO in expression vector.
Further experiments to complete the investigation 尚未完成的部份:
(1) The effect of Matrin3 and p54nrb/NonO overexpression in HDV RNA replication.
To analyze the effect of Matrin3 overexpression in HDV replication, Plasmids of pcDNA-Matrin3 and HDV 1.9-kb/pKS will be cotransfected into Huh7 cells with Lipofectamine 2000. At the indicated time points (1, 2, 3, 4 days of posttranfection), the protein lysates and RNAs will be prepared for Western and Northern blots analysis. The data will be compared with the samples that cotransfected with pcDNA empty vector and HDV 1.9-kb/pKS to reveal the effect of Matrin3 overexpression by the levels of detected L- and S-HDAg in Western blot. Meanwhile, the levels of monomeric and dimeric HDV RNAs will be compared with the control blots also.
(2) Application of RNA interference of Matrin3 followed by Western and Northern blots analysis of HDAgs and HDV RNA.
For silencing the Matrin3 gene expression a set of 3 target-specific 20-25 bp siRNAMtr3 (Santa Cruz Biotech., CA) was used: siRNAaMtr3 (411) 5'-GCUACCCAGUCUUUAAGUAtt-3', siRNAbMtr3
(926)5'-CCAUGGAAGUCGUUCUCAAtt-3', siRNAcMtr3 (1504)
5'-CUAGCAGAGUUGUUCACAUtt-3'. They hit all two isoforms of matrin 3. As a negative control the functional non-targeting siRNAC is used, containing four mismatches for any human, mouse, and
rat gene. The sequence of its sense strand was 5'-UAGCGACUAAACA CAUCAAUU-3'. Synthetic siRNAs (40 nM) and HDV 1.9-kb/pKS plasmids are cotransfected to 50-60% confluent Huh7 cells with Lipofectamine 2000. MATR3 and HDAgs levels in whole cell lysates, and extracted RNA that collected at 1, 2, 3, and 4 days after transfection, will be estimated by Western and Northern blotting.
According to our postulation, repression of MATR3 would result in reducing levels of HDAgs and HDV RNAs.
Expected results:
Matrin3 enhances HDV RNA replication if HDAgs and RNA levels are increased in Matrin3 cotransfection sets of samples. Otherwise, the results will indicate the inhibition of HDV replication by Matrin3 overexpression. In contrast, repression of Matrin3 by RNA interference would reduce HDV replication. Matrin3 will be detected by monoclonal antibodies (1:1000) to Matrin3. From the early report, Matrin3 is shown to bind Rev/RRE-containing viral RNA. This binding interaction stabilizes unspliced and partially spliced HIV-1 transcripts leading to increased cytoplasmic expression of these viral RNAs (Yedavalli & Jeang, 2011). Therefore, we postulate that high level of Matrin3 expression would enhance the HDV replication.
文獻探討
Abrahem, A. & Pelchat, M. (2008). Formation of an RNA polymerase II preinitiation complex on an RNA promoter derived from the hepatitis delta virus RNA genome. Nucleic Acids Res 36, 5201-5211.
Andersen, J. S., Lyon, C. E., Fox, A. H., Leung, A. K., Lam, Y. W., Steen, H., Mann, M. &
Lamond, A. I. (2002). Directed proteomic analysis of the human nucleolus. Curr Biol 12, 1-11.
Beausoleil, S. A., Jedrychowski, M., Schwartz, D., Elias, J. E., Villen, J., Li, J., Cohn, M. A., Cantley, L. C. & Gygi, S. P. (2004). Large-scale characterization of HeLa cell nuclear phosphoproteins. Proc Natl Acad Sci U S A 101, 12130-12135.
Berezney, R., Mortillaro, M. J., Ma, H., Wei, X. & Samarabandu, J. (1995). The nuclear matrix: a structural milieu for genomic function. Int Rev Cytol 162A, 1-65.
Bergmann, K. F. & Gerin, J. L. (1986). Antigens of hepatitis delta virus in the liver and serum of humans and animals. J Infect Dis 154, 702-706.
Bladen, C. L., Udayakumar, D., Takeda, Y. & Dynan, W. S. (2005). Identification of the polypyrimidine tract binding protein-associated splicing factor.p54(nrb) complex as a candidate DNA double-strand break rejoining factor. J Biol Chem 280, 5205-5210.
Bond, C. S. & Fox, A. H. (2009). Paraspeckles: nuclear bodies built on long noncoding RNA. J Cell Biol 186, 637-644.
Bonino, F., Heermann, K. H., Rizzetto, M. & Gerlich, W. H. (1986). Hepatitis delta virus: protein composition of delta antigen and its hepatitis B virus-derived envelope. J Virol 58, 945-950.
Buxade, M., Morrice, N., Krebs, D. L. & Proud, C. G. (2008). The PSF.p54nrb complex is a novel Mnk substrate that binds the mRNA for tumor necrosis factor alpha. J Biol Chem 283, 57-65.
Cao, D., Haussecker, D., Huang, Y. & Kay, M. A. (2009). Combined proteomic-RNAi screen for host factors involved in human hepatitis delta virus replication. RNA 15, 1971-1979.
Casey, J. L. & Gerin, J. L. (1995). Hepatitis D virus RNA editing: specific modification of adenosine in the antigenomic RNA. J Virol 69, 7593-7600.
Chang, F. L., Chen, P. J., Tu, S. J., Wang, C. J. & Chen, D. S. (1991). The large form of hepatitis delta antigen is crucial for assembly of hepatitis delta virus. Proc Natl Acad Sci U S A 88, 8490-8494.
Chao, M., Hsieh, S. Y. & Taylor, J. (1990). Role of two forms of hepatitis delta virus antigen:
evidence for a mechanism of self-limiting genome replication. J Virol 64, 5066-5069.
Chen, C. W., Tsay, Y. G., Wu, H. L., Lee, C. H., Chen, D. S. & Chen, P. J. (2002). The
double-stranded RNA-activated kinase, PKR, can phosphorylate hepatitis D virus small delta antigen at functional serine and threonine residues. J Biol Chem 277, 33058-33067.
Chen, P. J., Kalpana, G., Goldberg, J., Mason, W., Werner, B., Gerin, J. & Taylor, J. (1986).
Structure and replication of the genome of the hepatitis delta virus. Proc Natl Acad Sci U S A 83, 8774-8778.
Chou, H. C., Hsieh, T. Y., Sheu, G. T. & Lai, M. M. (1998). Hepatitis delta antigen mediates the nuclear import of hepatitis delta virus RNA. J Virol 72, 3684-3690.
Circle, D. A., Neel, O. D., Robertson, H. D., Clarke, P. A. & Mathews, M. B. (1997). Surprising specificity of PKR binding to delta agent genomic RNA. RNA 3, 438-448.
Dingle, K., Bichko, V., Zuccola, H., Hogle, J. & Taylor, J. (1998). Initiation of hepatitis delta virus genome replication. J Virol 72, 4783-4788.
Dong, B., Horowitz, D. S., Kobayashi, R. & Krainer, A. R. (1993). Purification and cDNA cloning of HeLa cell p54nrb, a nuclear protein with two RNA recognition motifs and extensive homology to human splicing factor PSF and Drosophila NONA/BJ6. Nucleic Acids Res 21, 4085-4092.
Dong, X., Sweet, J., Challis, J. R., Brown, T. & Lye, S. J. (2007). Transcriptional activity of androgen receptor is modulated by two RNA splicing factors, PSF and p54nrb. Mol Cell Biol 27, 4863-4875.
Dreger, M., Bengtsson, L., Schoneberg, T., Otto, H. & Hucho, F. (2001). Nuclear envelope
proteomics: novel integral membrane proteins of the inner nuclear membrane. Proc Natl Acad Sci U S A 98, 11943-11948.
Fey, E. G. & Penman, S. (1988). Nuclear matrix proteins reflect cell type of origin in cultured human cells. Proc Natl Acad Sci U S A 85, 121-125.
Fu, T. B. & Taylor, J. (1993). The RNAs of hepatitis delta virus are copied by RNA polymerase II in nuclear homogenates. J Virol 67, 6965-6972.
Glenn, J. S., Taylor, J. M. & White, J. M. (1990). In vitro-synthesized hepatitis delta virus RNA initiates genome replication in cultured cells. J Virol 64, 3104-3107.
Greco-Stewart, V. S., Miron, P., Abrahem, A. & Pelchat, M. (2007). The human RNA polymerase II interacts with the terminal stem-loop regions of the hepatitis delta virus RNA genome. Virology 357, 68-78.
Greco-Stewart, V. S., Schissel, E. & Pelchat, M. (2009). The hepatitis delta virus RNA genome interacts with the human RNA polymerases I and III. Virology 386, 12-15.
Greco-Stewart, V. S., Thibault, C. S. & Pelchat, M. (2006). Binding of the polypyrimidine
tract-binding protein-associated splicing factor (PSF) to the hepatitis delta virus RNA. Virology 356, 35-44.
Gudima, S., Dingle, K., Wu, T. T., Moraleda, G. & Taylor, J. (1999). Characterization of the 5' ends for polyadenylated RNAs synthesized during the replication of hepatitis delta virus. J Virol 73, 6533-6539.
Hakes, D. J. & Berezney, R. (1991). DNA binding properties of the nuclear matrix and individual nuclear matrix proteins. Evidence for salt-resistant DNA binding sites. J Biol Chem 266, 11131-11140.
Hata, K., Nishimura, R., Muramatsu, S., Matsuda, A., Matsubara, T., Amano, K., Ikeda, F., Harley, V. R. & Yoneda, T. (2008). Paraspeckle protein p54nrb links Sox9-mediated transcription with RNA processing during chondrogenesis in mice. J Clin Invest 118, 3098-3108.
Huang, W. H., Mai, R. T. & Lee, Y. H. (2008). Transcription factor YY1 and its associated
acetyltransferases CBP and p300 interact with hepatitis delta antigens and modulate hepatitis delta virus RNA replication. J Virol 82, 7313-7324.
Huang, W. H., Yung, B. Y., Syu, W. J. & Lee, Y. H. (2001). The nucleolar phosphoprotein B23 interacts with hepatitis delta antigens and modulates the hepatitis delta virus RNA replication. J Biol Chem 276, 25166-25175.
Inagaki, H., Matsushima, Y., Nakamura, K., Ohshima, M., Kadowaki, T. & Kitagawa, Y. (1996).
A large DNA-binding nuclear protein with RNA recognition motif and serine/arginine-rich domain. J Biol Chem 271, 12525-12531.
Ito, T., Watanabe, H., Yamamichi, N., Kondo, S., Tando, T., Haraguchi, T., Mizutani, T., Sakurai, K., Fujita, S., Izumi, T., Isobe, T. & Iba, H. (2008). Brm transactivates the telomerase
reverse transcriptase (TERT) gene and modulates the splicing patterns of its transcripts in concert with p54(nrb). Biochem J 411, 201-209.
Kameoka, S., Duque, P. & Konarska, M. M. (2004). p54(nrb) associates with the 5' splice site within large transcription/splicing complexes. EMBO J 23, 1782-1791.
Kaneko, S., Rozenblatt-Rosen, O., Meyerson, M. & Manley, J. L. (2007). The multifunctional protein p54nrb/PSF recruits the exonuclease XRN2 to facilitate pre-mRNA 3' processing and transcription termination. Genes Dev 21, 1779-1789.
Kos, A., Dijkema, R., Arnberg, A. C., van der Meide, P. H. & Schellekens, H. (1986). The hepatitis delta (delta) virus possesses a circular RNA. Nature 323, 558-560.
Kuo, M. Y., Chao, M. & Taylor, J. (1989). Initiation of replication of the human hepatitis delta virus genome from cloned DNA: role of delta antigen. J Virol 63, 1945-1950.
Kuo, M. Y., Goldberg, J., Coates, L., Mason, W., Gerin, J. & Taylor, J. (1988a). Molecular cloning of hepatitis delta virus RNA from an infected woodchuck liver: sequence, structure, and
applications. J Virol 62, 1855-1861.
Kuo, M. Y., Sharmeen, L., Dinter-Gottlieb, G. & Taylor, J. (1988b). Characterization of
self-cleaving RNA sequences on the genome and antigenome of human hepatitis delta virus. J Virol 62, 4439-4444.
Lee, C. H., Chang, S. C., Chen, C. J. & Chang, M. F. (1998). The nucleolin binding activity of
hepatitis delta antigen is associated with nucleolus targeting. J Biol Chem 273, 7650-7656.
Lee, C. Z., Lin, J. H., Chao, M., McKnight, K. & Lai, M. M. (1993). RNA-binding activity of hepatitis delta antigen involves two arginine-rich motifs and is required for hepatitis delta virus RNA replication. J Virol 67, 2221-2227.
Li, Y. J., Macnaughton, T., Gao, L. & Lai, M. M. (2006). RNA-templated replication of hepatitis delta virus: genomic and antigenomic RNAs associate with different nuclear bodies. J Virol 80, 6478-6486.
Liao, F. T., Hsu, L. S., Ko, J. L., Lin, C. C. & Sheu, G. T. (2012). Multiple genomic sequences of hepatitis delta virus are associated with cDNA promoter activity and RNA double rolling-circle replication. J Gen Virol 93, 577-587.
Liao, F. T., Lee, Y. J., Ko, J. L., Tsai, C. C., Tseng, C. J. & Sheu, G. T. (2009). Hepatitis delta virus epigenetically enhances clusterin expression via histone acetylation in human hepatocellular carcinoma cells. J Gen Virol 90, 1124-1134.
Lin, J. H., Chang, M. F., Baker, S. C., Govindarajan, S. & Lai, M. M. (1990). Characterization of hepatitis delta antigen: specific binding to hepatitis delta virus RNA. J Virol 64, 4051-4058.
Lin, S. S., Chang, S. C., Wang, Y. H., Sun, C. Y. & Chang, M. F. (2000). Specific interaction between the hepatitis delta virus RNA and glyceraldehyde 3-phosphate dehydrogenase: an enhancement on ribozyme catalysis. Virology 271, 46-57.
Lo, K., Sheu, G. T. & Lai, M. M. (1998). Inhibition of Cellular RNA polymerase II transcription by delta antigen of hepatitis delta virus. Virology 247, 178-188.
Luo, G. X., Chao, M., Hsieh, S. Y., Sureau, C., Nishikura, K. & Taylor, J. (1990). A specific base transition occurs on replicating hepatitis delta virus RNA. J Virol 64, 1021-1027.
Makino, S., Chang, M. F., Shieh, C. K., Kamahora, T., Vannier, D. M., Govindarajan, S. & Lai, M. M. (1987). Molecular cloning and sequencing of a human hepatitis delta (delta) virus RNA.
Nature 329, 343-346.
Malyavantham, K. S., Bhattacharya, S., Barbeitos, M., Mukherjee, L., Xu, J., Fackelmayer, F. O.
& Berezney, R. (2008). Identifying functional neighborhoods within the cell nucleus:
proximity analysis of early S-phase replicating chromatin domains to sites of transcription, RNA polymerase II, HP1gamma, matrin 3 and SAF-A. J Cell Biochem 105, 391-403.
Modahl, L. E. & Lai, M. M. (1998). Transcription of hepatitis delta antigen mRNA continues throughout hepatitis delta virus (HDV) replication: a new model of HDV RNA transcription and replication. J Virol 72, 5449-5456.
Modahl, L. E. & Lai, M. M. (2000). The large delta antigen of hepatitis delta virus potently inhibits genomic but not antigenomic RNA synthesis: a mechanism enabling initiation of viral
replication. J Virol 74, 7375-7380.
Modahl, L. E., Macnaughton, T. B., Zhu, N., Johnson, D. L. & Lai, M. M. (2000).
RNA-Dependent replication and transcription of hepatitis delta virus RNA involve distinct cellular RNA polymerases. Mol Cell Biol 20, 6030-6039.
Mota, S., Mendes, M., Freitas, N., Penque, D., Coelho, A. V. & Cunha, C. (2009). Proteome analysis of a human liver carcinoma cell line stably expressing hepatitis delta virus ribonucleoproteins. J Proteomics 72, 616-627.
Mota, S., Mendes, M., Penque, D., Coelho, A. V. & Cunha, C. (2008). Changes in the proteome of Huh7 cells induced by transient expression of hepatitis D virus RNA and antigens. J
Proteomics 71, 71-79.
Patton, J. G., Porro, E. B., Galceran, J., Tempst, P. & Nadal-Ginard, B. (1993). Cloning and characterization of PSF, a novel pre-mRNA splicing factor. Genes Dev 7, 393-406.
Perrotta, A. T. & Been, M. D. (1990). The self-cleaving domain from the genomic RNA of hepatitis delta virus: sequence requirements and the effects of denaturant. Nucleic Acids Res 18, 6821-6827.
Pohl, C., Baroudy, B. M., Bergmann, K. F., Cote, P. J., Purcell, R. H., Hoofnagle, J. & Gerin, J. L.
(1987). A human monoclonal antibody that recognizes viral polypeptides and in vitro translation products of the genome of the hepatitis D virus. J Infect Dis 156, 622-629.
Polson, A. G., Bass, B. L. & Casey, J. L. (1996). RNA editing of hepatitis delta virus antigenome by dsRNA-adenosine deaminase. Nature 380, 454-456.
Polson, A. G., Ley, H. L., 3rd, Bass, B. L. & Casey, J. L. (1998). Hepatitis delta virus RNA editing is highly specific for the amber/W site and is suppressed by hepatitis delta antigen. Mol Cell Biol 18, 1919-1926.
Robertson, H. D., Manche, L. & Mathews, M. B. (1996). Paradoxical interactions between human delta hepatitis agent RNA and the cellular protein kinase PKR. J Virol 70, 5611-5617.
Rosonina, E., Ip, J. Y., Calarco, J. A., Bakowski, M. A., Emili, A., McCracken, S., Tucker, P., Ingles, C. J. & Blencowe, B. J. (2005). Role for PSF in mediating transcriptional
activator-dependent stimulation of pre-mRNA processing in vivo. Mol Cell Biol 25, 6734-6746.
Sharmeen, L., Kuo, M. Y., Dinter-Gottlieb, G. & Taylor, J. (1988). Antigenomic RNA of human hepatitis delta virus can undergo self-cleavage. J Virol 62, 2674-2679.
Shav-Tal, Y. & Zipori, D. (2002). PSF and p54(nrb)/NonO--multi-functional nuclear proteins. FEBS Lett 531, 109-114.
Sheu, G. T. & Lai, M. M. (2000). Recombinant hepatitis delta antigen from E. coli promotes hepatitis delta virus RNA replication only from the genomic strand but not the antigenomic strand.
Virology 278, 578-586.
Sikora, D., Greco-Stewart, V. S., Miron, P. & Pelchat, M. (2009). The hepatitis delta virus RNA genome interacts with eEF1A1, p54(nrb), hnRNP-L, GAPDH and ASF/SF2. Virology 390, 71-78.
Taylor, J. M. (1990). Hepatitis delta virus: cis and trans functions required for replication. Cell 61, 371-373.
Wang, K. S., Choo, Q. L., Weiner, A. J., Ou, J. H., Najarian, R. C., Thayer, R. M., Mullenbach, G.
T., Denniston, K. J., Gerin, J. L. & Houghton, M. (1986). Structure, sequence and expression of the hepatitis delta (delta) viral genome. Nature 323, 508-514.
Weiner, A. J., Choo, Q. L., Wang, K. S., Govindarajan, S., Redeker, A. G., Gerin, J. L. &
Houghton, M. (1988). A single antigenomic open reading frame of the hepatitis delta virus encodes the epitope(s) of both hepatitis delta antigen polypeptides p24 delta and p27 delta. J Virol 62, 594-599.
Wong, S. K. & Lazinski, D. W. (2002). Replicating hepatitis delta virus RNA is edited in the nucleus by the small form of ADAR1. Proc Natl Acad Sci U S A 99, 15118-15123.
Wu, H. N., Lin, Y. J., Lin, F. P., Makino, S., Chang, M. F. & Lai, M. M. (1989). Human hepatitis delta virus RNA subfragments contain an autocleavage activity. Proc Natl Acad Sci U S A 86, 1831-1835.
Yamaguchi, Y., Filipovska, J., Yano, K., Furuya, A., Inukai, N., Narita, T., Wada, T., Sugimoto, S., Konarska, M. M. & Handa, H. (2001). Stimulation of RNA polymerase II elongation by hepatitis delta antigen. Science 293, 124-127.
Yang, Y. S., Hanke, J. H., Carayannopoulos, L., Craft, C. M., Capra, J. D. & Tucker, P. W. (1993).
NonO, a non-POU-domain-containing, octamer-binding protein, is the mammalian homolog of Drosophila nonAdiss. Mol Cell Biol 13, 5593-5603.
Yedavalli, V. S. & Jeang, K. T. (2011). Matrin 3 is a co-factor for HIV-1 Rev in regulating post-transcriptional viral gene expression. Retrovirology 8, 61.
Zeitz, M. J., Malyavantham, K. S., Seifert, B. & Berezney, R. (2009). Matrin 3: chromosomal distribution and protein interactions. J Cell Biochem 108, 125-133.
Zhang, Z. & Carmichael, G. G. (2001). The fate of dsRNA in the nucleus: a p54(nrb)-containing complex mediates the nuclear retention of promiscuously A-to-I edited RNAs. Cell 106, 465-475.
已發表論文:
Hsiao YP, Yang JH, Wu WJ, Lin MH, Sheu* GT. (2013 Oct) E6 and E7 of human papillomavirus type 18 and UVB irradiation corporately regulate interleukin-6 and interleukin-8 expressions in basal cell carcinoma. Exp Dermatol. 22(10):672-674. doi: 10.1111/exd.12223, PMID: 24079741 (SCI) (IF=3.578 R/C=6/58=10.3%, DERMATOLOGY) (NSC-101-2320-B-040-006)
國科會補助計畫衍生研發成果推廣資料表
日期:2013/10/17
國科會補助計畫
計畫名稱: 細胞核基質蛋白Matrin 3 (MATR3)在人類D型肝炎病毒RNA的合成過程中的重 要性
計畫主持人: 許國堂
計畫編號: 101-2320-B-040-006- 學門領域: 寄生蟲學、醫事技術及實驗診斷
無研發成果推廣資料
101 年度專題研究計畫研究成果彙整表
計畫主持人:許國堂 計畫編號:101-2320-B-040-006-
計畫名稱:細胞核基質蛋白 Matrin 3 (MATR3)在人類 D 型肝炎病毒 RNA 的合成過程中的重要性 量化
成果項目 實際已達成
數(被接受 或已發表)
預期總達成 數(含實際已
達成數)
本計畫實 際貢獻百
分比
單位
備 註 ( 質 化 說 明:如 數 個 計 畫 共 同 成 果、成 果 列 為 該 期 刊 之 封 面 故 事 ...
等)
期刊論文 0 0 50%
研究報告/技術報告 0 0 100%
研討會論文 0 0 100%
論文著作 篇
專書 0 0 100%
申請中件數 0 0 100%
專利 已獲得件數 0 0 100% 件
件數 0 0 100% 件
技術移轉
權利金 0 0 100% 千元
碩士生 0 0 100%
博士生 0 0 100%
博士後研究員 0 0 100%
國內
參與計畫人力
(本國籍)
專任助理 0 0 100%
人次
期刊論文 1 2 50%
研究報告/技術報告 0 0 100%
研討會論文 0 0 100%
論文著作 篇
專書 0 0 100% 章/本
申請中件數 0 0 100%
專利 已獲得件數 0 0 100% 件
件數 0 0 100% 件
技術移轉
權利金 0 0 100% 千元
碩士生 0 0 100%
博士生 1 0 100%
博士後研究員 0 0 100%
國外
參與計畫人力
(外國籍)
專任助理 0 0 100%
人次
其他成果
(無法以量化表達之成
果如辦理學術活動、獲 得獎項、重要國際合 作、研究成果國際影響 力及其他協助產業技 術發展之具體效益事 項等,請以文字敘述填 列。)
無
成果項目 量化 名稱或內容性質簡述
測驗工具(含質性與量性) 0
課程/模組 0
電腦及網路系統或工具 0
教材 0
舉辦之活動/競賽 0
研討會/工作坊 0
電子報、網站 0
科 教 處 計 畫 加 填 項
目 計畫成果推廣之參與(閱聽)人數 0
國科會補助專題研究計畫成果報告自評表
請就研究內容與原計畫相符程度、達成預期目標情況、研究成果之學術或應用價 值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)、是否適 合在學術期刊發表或申請專利、主要發現或其他有關價值等,作一綜合評估。
1. 請就研究內容與原計畫相符程度、達成預期目標情況作一綜合評估
□達成目標
■未達成目標(請說明,以 100 字為限)
□實驗失敗
□因故實驗中斷
■其他原因 說明:
已達成目標約 70% 未達成目標原因
說明:此計畫之後續實驗因為其複雜性甚高,需要資深的研究生來操作。當下資深的研究 生已經畢業,而新的學生尚無能力執行後段複雜的研究。若能有三年的計畫來訓練和執行應 該會有比較好的結果,只有壹年的計畫實在很不容易吸引研究生,尤其是在私立大學的招生 相對為弱勢之下。
2. 研究成果在學術期刊發表或申請專利等情形:
論文:□已發表 □未發表之文稿 ■撰寫中 □無 專利:□已獲得 □申請中 ■無
技轉:□已技轉 □洽談中 ■無 其他:(以 100 字為限)
已發表 Hsiao YP, Yang JH, Wu WJ, Lin MH, Sheu* GT. (2013 Oct) E6 and E7 of human papillomavirus type 18 and UVB irradiation corporately regulate interleukin-6 and interleukin-8 expressions in basal cell carcinoma. Exp Dermatol. 22(10):672-674. doi:
10.1111/exd.12223, PMID: 24079741 (SCI) (IF=3.578 R/C=6/58=10.3%, DERMATOLOGY) (NSC-101-2320-B-040-006)
3. 請依學術成就、技術創新、社會影響等方面,評估研究成果之學術或應用價 值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)(以 500 字為限)
如果我們可以辨識細胞內負責合成 HDV 病毒的 HDV 核酸聚合酶(RdRP)的工作羣,我們將有 機會瞭解到 pol II 工作羣如何和 HDV RNA 進行作用,這對於醫學和生物學上都可以有相 當的影響性。