Use of otolith microchemistry to investigate
the environmental history of European eel
Anguilla anguilla
'Department o f Zoology, College of Science, National Taiwan University, Taipei, Taiwan 10617, ROC 2 ~ e p a r t r n e n t of Geology and Geophysics. University of Alaska Fairbanks, Alaska 997 75-0760, USA
31nstitute of Freshwater Research, S - 1 7 8 93 Drottningholrn, S w e d e n
ABSTRACT Environmental histoly of t h e E u r o p e a n e e l Angu~lla angullla (L ) collected from S w e d i s h bracklsh w a t e r s a n d l a k e s w a s s t u d l e d by e x a m i n i n g s t r o n t i u m ( S r ) a n d calcium ( C a ) in their otoliths w ~ t h w a v e l e n g t h d ~ s p e r s i v e X-lay s p e c t ~ o m e t r y on a n electron m l c r o p r o b e T h e S r / C a ratios In t h e otollths c h a n g e d w ~ t h both o n t o g e n e t i c d e v e l o p m e n t a n d s a l i n ~ t y of t h e h a b ~ t a t A p e a k S r / C a ratlo of approximately 2 X 10-L w a s o b s e r v e d in t h e m a n n e p h a s e of t h e o t o l ~ t h s c o r r e s p o n d i n g to t h e p e r ~ o d
w h e n a l e p t o c e p h a l u s m e t a m o r p h o s e d into a glass e e l T h e ratios d e c r e a s e d to 5 72 (+ 0 3 2 ) X 1 0 a t
yellow e e l s t a g e w h e n t h e fish lived in brackish w a t e r a n d to 3 7 (t 0 2 0 ) X 10.' In fresh w a t e r T h c ratios
w e r e s ~ g n ~ f i c a n t l y different b e t w e e n brackish a n d fresh w a t e r ( p < 0 00001) Several r e l a t ~ v e l y h ~ g h e ~ Sr b a n d s w e r e found In o t o l ~ t h s of e e l from bracklsh w a t e r s b u t not In t h e e e l s from fresh w a t e l Salin- ity probably influences Sr deposition m o l e t h a n d o e s t e m p e r a t u ~ e 01 somatic g ~ o w t h rates
KEY WORDS: E u r o p e a n e e l . O t o l ~ t h . Strontium/calcium ratlos . Elemental X-ray m a p . Environmental factor M ~ g r a t o r y h ~ s t o r y . Electron m ~ c r o p r o b e
INTRODUCTION
European eel Anguilla angujlla (L.) is the most a b u n - dant of the 18 species of the genus Anguilla in the
world. It is widely distributed in fresh water a n d marine littoral areas of North Africa, the Mediterranean Sea, the British Isles, Iceland, and the western and northern European continent. After spawning in the Sargasso Sea, eel larvae (leptocephali) are transported by the North Equatorial Current, Gulf Stream a n d North At- l a n t ~ c Current to the continental shelf of northern Euro- pean countries. The larvae metamorphose into glass eels in coastal waters. Glass eels become pigmented elvers when they enter estuaries. Some elvers stay In salt or brackish water along the coast while others penetrate rivers and streams to conlplete the growing stage (yellow eel) in fresh water. Finally, they transform into silver eels a n d start the downstream migration back to the Sargasso Sea (Bertin 1956, Tesch 1983).
- .
'E-mail: wnt@ccms.ntu e d u tw
The Baltic Sea is a n important growth area for Euro- pean eel. Elvers reach the Danish Straits, the Sound area between Denmark and Sweden, and the southern part of the Baltic Sea in late spring. The young eels then pro- ceed northwards more slowly but little is known about this gradual dispersal (Svardson 1976). Commercial catches of silver eels along the coasts of southeastern Sweden a n d in the outlets of the Baltic Sea have de- creased considerably since the mid-1960s (Wickstrom 1986). This is because the recruitment of elvers entering the Baltic Sea from the Skagerrak a n d North Sea has been declining steadily for a long period (Erichsen 1976, SvBrdson 1976, Hagstrom & MIickstrom 1990, W e s t ~ n 1990). As a consequence, stocking programs have been undertaken since the 1940s in the Baltic Sea to try to strengthen the recruitment of elvers a n d subsequently improve eel stocks (Leopold 1976, Filuk & Wiktor 1988). For stocking, elvers have been imported from France a n d the Brit~sh Isles. Domestic young eels collected on the western coast of Sweden were also restocked in Swedish lakes and the Baltic coast (Wickstrom 1984).
O Inter-Research 1997
Mar Ecol Prog Ser 149: 73-81, 1997
Therefore, the eel population in Sweden has a number of different origins. Distinguishing the origin of the eels is important for evaluating the effect of stocking.
Strontium content in the otolith of diadromous fish has been found to differ between fresh water and sea water life history phases (Casselman 1982, Radtke et al. 1988, Kalish 1990, Secor 1992, Otake e t al. 1994, Tzeng & Tsai 1994, Limburg 1995). The difference cor- responds to the ambient Sr concentration which is higher in marine than fresh water (Odum 1957, Tzeng & Tsai 1994, Tzeng 1995, 1996). Accordingly, Sr con- tent in fish otoliths offers the potential to distinguish between time spent in sea water and fresh water and give information about the fishes' environmental history.
The purpose of this paper is to reconstruct the envi- ronmental history of European eels collected from Swedlsh brackish waters and lakes and to identify the environmental history of eels of unknown origin by examining Sr and Ca in their otoliths. The changes in Sr contents in otolith in relation to salinity, temperature and fish growth rate and ontogenetic development are discussed.
MATERIALS AND METHODS
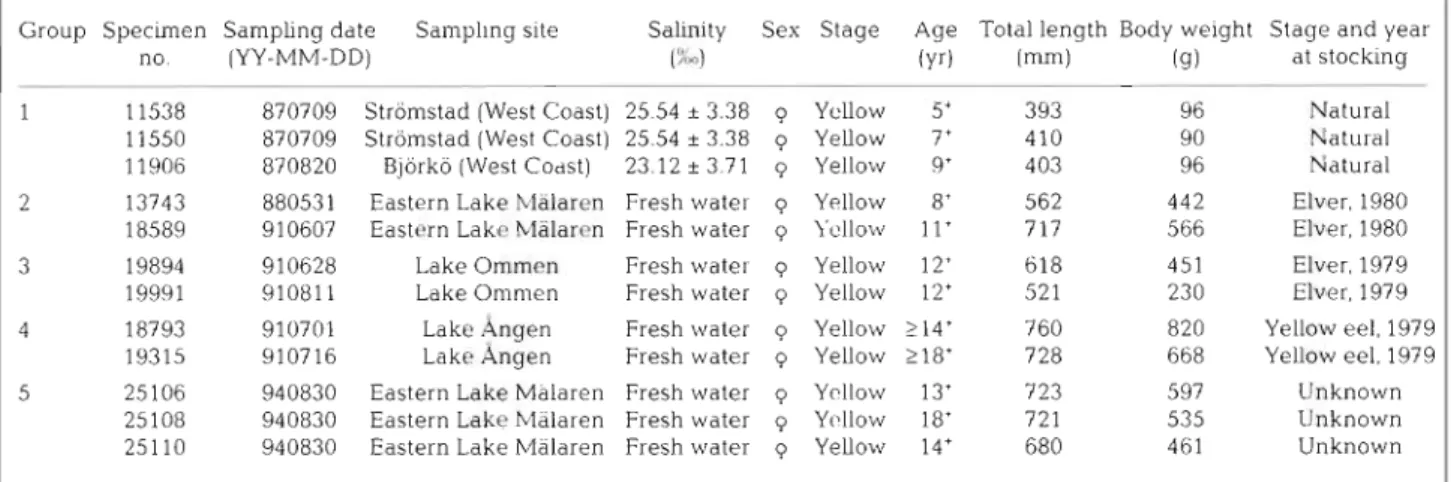
Sampling design. Twelve otollths from 5 groups of European eels collected from 3 lakes and 2 brackish water locations in Sweden were used for microchemi- cal analysis (Fig. 1). The origin of Groups 1 through 4 was clear: these eels were captured from areas where the restocking program is well known. The origin of the eels in Group 5 was unknown. The stocking and sampling dates and biological characteristics of the 5 groups of eels (including sampling date, mean (r SD) salinity of sampllng sites, sex, developmental stage,
a g e , total length, body weight, and the stage and year at stocking) are given in Table 1. The life history aspects of the 5 groups were as follows:
Group 1: collected from the brackish waters off Stromstad and Bjorko on the west coast of Sweden in 1987. This group should have not experienced fresh water life because eels in the western coastal waters a r e recruited directly from the Atlantic (Kattegat- Skagerrak) and no stocking has ever been performed there. The eels ranged from 4 to 11 yr old, with the majority between 6 and 9 yr old (Holmgren & Wick- strom 1988).
Group 2: collected from the eastern part of Lake Malaren in 1988 and 1991. The eels were stocked as elvers, which were imported from the Bay of Biscay, France, in the spring of 1980, and had lived in fresh water for 8 and 11 yr, respectively.
Group 3: collected from Lake Ommen in eastern Sweden In 1991. The eels were stocked at the elver stage, having been imported from France in 1979. The eels had lived in fresh water approximately 12 yr (Tzeng et al. 1995).
Group 4: collected from Lake Angen in eastern Sweden In 1991. The eels were stocked a t the yellow eel stage after being caught in brackish waters along the west coast of Sweden in 1979. The lake was eel- free before stocking in 1979 (Wickstrom 1984). It is estimated that the eels lived in the brackish waters and the fresh water lake for approximately 7 a n d 12 yr, respectively (Holmgren & Wickstrom 1988).
Group 5: collected in the eastern part of Lake Malaren in 1994 a few km away from where Group 2 was collected. The origin of this group was unknown. They were derived either from a natural (wild) popula- tion that had migrated from brackish Baltic Sea, or from a stocked population caught from brackish waters on the west coast of Sweden and released at the yellow eel
Fig. 1. Map showing the locations where eels were collected from brackish waters along the western coast of Sweden (1, Stromstad; 2, Bjorko) and 3 lakes on the eastern coast (A, Lake Malaren; B. Lake Angen; C, Lake Ommen). Temperature and salinity were measured in the western coastal waters
Table 1. L ~ f e history of the 5 groups of European eels used in thls study
Group Specunen Samphng date Salnpllng slte Salrnlty Sex Stage Age Total length Body w e ~ g h t Stage and year
no (YY-MM-DD) I r 4 (yr) [mm) ( 9 ) at stocking
1 11538 870709 Stromstad (West Coast) 25 54 k 3 38 q Yellow 5' 393 9 6 Natural 11550 870709 Stromstad (West Coast) 25 54 t 3 38 q Yellow 7' 410 9 0 h'aturdl 11900 870820 Blorko (West Codst) 23 12 t 3 71 q Yellow 9' 403 96 Natural 2 13743 880531 Eastern Lake hlalarcn Fresh w a t e ~ q Y ~ l l o w 8' 562 442 Elver 1980
18589 910607 Eastern Lakc hlalarrn Fresh water q 1 cllo~v 11' 717 566 Elver 1980 3 19894 910628 Lake O m r n ~ n Fresh w a t e ~ q Yellow 12' 618 451 Elver 1979 l9991 910811 Ldke Oninien Fresh water q Yellobv 12' 521 230 Elver 1979 4 18793 910701 Lakc 4ngen Fresh water q Yellow > 14' 760 820 Yellow eel 1979
19315 910716 Lakc Angen Fresh water q Yellow 218* 728 6b8 Yellow eel 1979 5 2.5106 940830 Eastern Lake Malaren Fresh water q Ycllow 13' 723 597 IJnknolvn
25 108 940830 Edstern Lakc hlalaren Flesh water q Y[lllo\v 18' 721 535 Unknown 25110 940830 Easteln Lake Malaren Flesh water q Yellow 14' 680 461 Unknown
stage at approximately 40 cm in total length, or from a method described in Williams (1987). The spatial distri- stocked population released at the elver stage Imported bution of Sr across each otolith was mapped at a n 8 pm from France or the British Isles (Wickstrom 1984). resolution using a 15 kV, 100 nA beam. X-rays were
Microchemical analysis. The otoliths removed from collected from each point (pixel) for 0.5 S .
the eel were cleaned with distilled water and dried in air. Maximum lengths of the otoliths were measured to
the nearest 0.1 pm 1 ~ 1 t h the aid of an image-processing RESULTS
system (VIPRO 512). T h e n , the otoliths were embed-
ded in thermo-epoxy (Petropoxy 154, Palouse Petro Seasonal changes in temperature and salinity
Products, USA) a n d cured for 1 h a t 135°C. Embedded
otoliths were ground from the proximal side of the Fig. 2 shows seasonal changes in surface tempera- sagittal plane of the fish until the primordium of the ture a n d salinity measured every 10 to 13 d (except the otolith was revealed. For microprobe analysis, the pol- years 1986-88) in the coastal waters off Kungsbacka ished otoliths were coated under vacuum with a 30 nm on the westel-n coast of Sweden from January 1980 layer of carbon to reduce X-ray diffraction a n d in- through July 1991. Temperature ranged from -1.6 crease electron conductance. The embedding, grind- to 21.1°C. Salinlty ranged 9.74 to 32.78%0. Seasonal ing, and coating procedures were similar to a previous change of temperature was regular, but not salinity. study (Tzeng & Tsai 1994). Salinity became higher w h e n temperature decreased.
Quantitative measurements and mapping of
the concentration of trace elements In otolith of
-
35 the eel were conducted using a n electron mi-g
30
croprobe equipped with wavelength dispersive
C
spectrometers (Cameca SX-50). Quantitative .E . - - 25
measurements of Na, Mg, Cl, P, S , K , C a , a n d Sr ca 0
were made at approximately 18 pm intervals 20 along a transect from the p]-imordium to the
5
e d g e of the otolith. Of these elements Sr was '- l5the best for d ~ s t ~ n g u i s h i n g between fresh water
2
10
and marine environments. For the measure-
$
ments of Sr and Ca, calclte (USNM136321) a n d 5strontian~te (Nh4NH10065) from the Depart-
ment of Mineral Sciences, N a t ~ o n a l Museum v
of Natural History, Smithsonian Institution,
p
-5Washington DC, USA were used as standards 1980 1982 1984 1986 1988 1990 1992
for callbration. A 15 kV, 4 nA, 15 pm diameter Time (yr)
beam was used. Counting times 20 Fig. 2. Seasonal changes in water temperature a n d s a l i n ~ t y In t h e and 120 S (Sr). Detection limits were 0,077 \vt'%l coastal waters off Kungsbacka In western S w e d e n from J a n u a r y 1980
76 Mar Ecol Prog Ser 149: 73-81, 1997
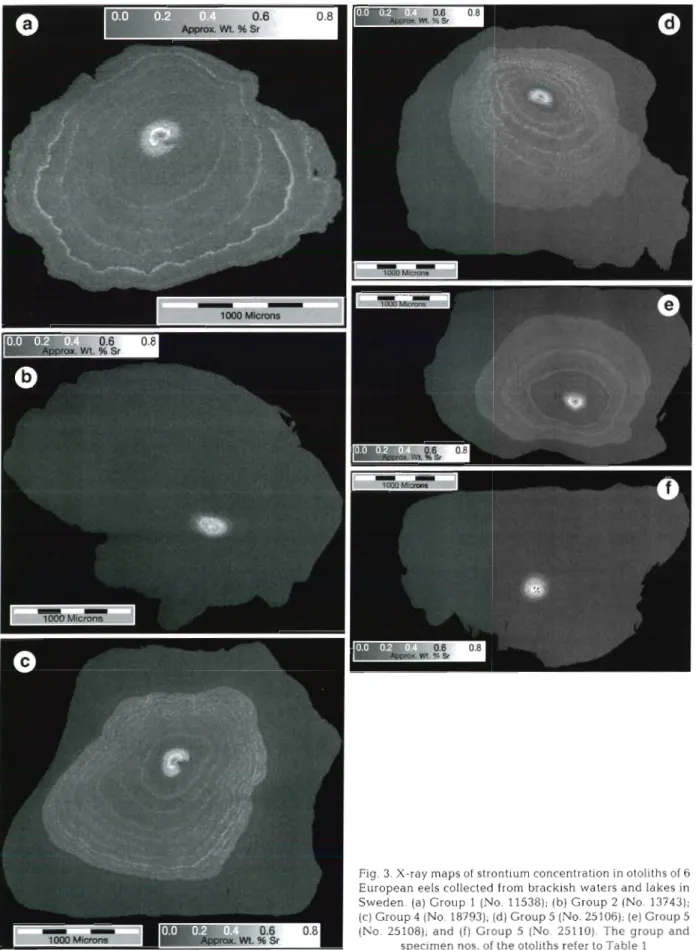
Fig. 3. X-ray maps of strontium concentration in otoliths of 6 European eels collected from brackish waters and lakes in Sweden. (a) Group 1 (No. 11538); (b) Group 2 (No. 13743); (c) Group 4 (No. 18793); (d) Group 5 (No. 25106); (e) Group 5 (No. 25108); and (f) Group 5 (No. 25110). The group and
X-ray m a p of Sr contents in otoliths Twelve X-ray maps of Sr contents in otoliths of the 5 groups of European eel were made (Table 1). Six of them a r e shown in Fig. 3. The brighter areas in the X-ray m a p indicate higher Sr con- centrations. The bright area of highest Sr content approximately equaled the size of the otolith of an elver, approxi- mately 156.6 pm in radius (Tzeng et al. 1995). In other words, the nucleus was deposited during marine life phase while the eel larvae migrated from its spawning ground to the estuary.
Beyond the nucleus the brightness in the X-ray images decreased, indicat- ing that the Sr concentrations d e - creased (Fig. 3). The amount of d e - crease d e p e n d e d on the salinity content of the habitat where the eel lived after elver stage. Sr content was higher for eels that lived in brackish waters (Fig. 3 a , Group 1) than for those from a fresh water environment (Fig. 3b, Group 2). This difference was more obvious in a n individual eel which had passed the brackish-fresh water gra- dient environment (Fig. 3c, Group 4). This suggested that the Sr content pat- tern in otoliths could be used to re- construct the history of salinity experi- enced by the eel.
The origin of Group 5 was unknown (Fig. 3d-f). The pattern of Sr content in otoliths of Nos. 25106 a n d 25108 (Fig. 3 d , e ) was similar to that of Group 4 (Fig. 3c), while No. 25110 (Fig. 3f) was s i m ~ l a r to that of Group 2 (Fig. 3 b ) .
Furthermore, the X-ray maps of Sr contents in otoliths for the eels in Groups 1 & 4 , a n d Nos. 25106 a n d 25108 in Group 5, revealed several bands of high Sr content, while no bands of high Sr content were found in the otoliths from Groups 2 a n d 3 .
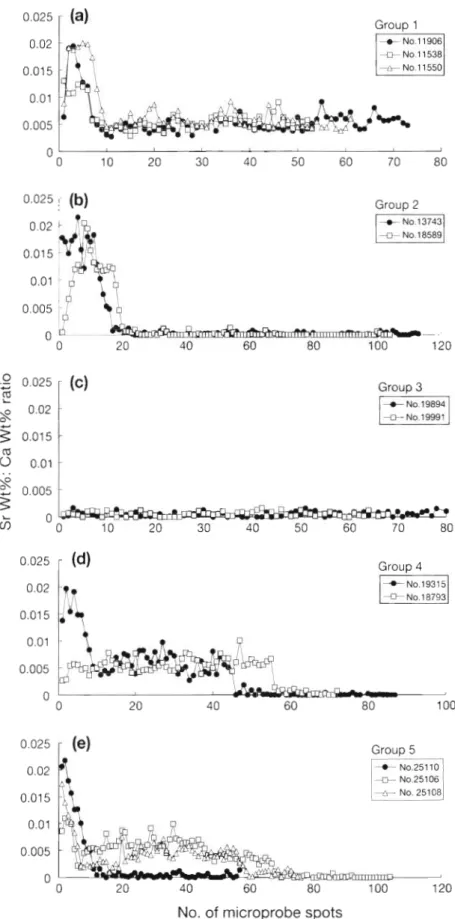
Sr/Ca ratios in otoliths of natural eels from brackish water ( G r o u p 1) Sr/Ca ratios measured along a tran- sect from the primordium to the otoli.th e d g e of the natural eels collected in the coastal waters off Stromstad a n d
Group 1 Group 2
I-+
No 137431 Group 3 No 19991 0.025[
(d)
0.02 .q, 0.015 Group 4 -CC No.18793 0.025r
(e)
Group 5 -. -:rNo 25106 ? No 251081No. of microprobe spots
Fig. 4 Sr/Ca ratios scanned along a transect from the primordium to the edge of otoliths of 12 eels collected from brackish waters and lakes of Sweden. The
Mar Ecol Prog Ser 149: 73-81, 1997
Bjorko o n the west coast of Sweden are shown in Fig. 4a. The ratios increased from the primordium and reached a peak of approximately 2 X 10-' about 113 pm from the primordium, then decreased and maintained a low level of approximately 5.72 X 10-3 until the edge. The Sr/Ca ratios beyond approximately the first 11 spots showed a periodic fluctuation. The peaks of the fluctuation correspond to the bright bands of the Sr X-ray maps as indicated in Fig 3a.
SrICa ratios in otoliths of eels stocked in fresh water lakes
Stocked a t elver stage (Groups 2 & 3). Sr/Ca ratios measured along a transect from the primordium to the otolith edge of eels collected from Lake Malaren (Group 2) and Lake Ommen (Group 3) are shown In Fig. 4b, c. The Sr/Ca ratios in the nuclei of the otoliths of Group 2 were similar to those of Group 1 (Fig. 4a). No peak Sr/Ca ratios were found in Group 3 because the otoliths were not ground through the primordium (Fig. 4c). The Sr/Ca ratios were significantly different between fresh water environment (Groups 2 & 3 ) and
brackish waters (Group 1) ( l - w a y ANOVA, F , , s = 643.85, p < 0.0001). The mean (i SD) Sr/Ca rat10
beyond the nuclei of the otoliths in Groups 2 & 3 was 3.7 (+ 0.20) X 1 0 - ~ , lower than Group 1 [5.72 (k 0.32) X 10-3].
Stocked from a brackish water yellow eel (Group 4). Sr/Ca ratios measured along a transect from the prl- mordium to the otolith edge of the eels collected from Lake Angen are shown in Fig. 4d. The eels were stocked at the yellow eel stage after being caught from brackish waters on the western coast of Sweden. They had experienced 3 different environments, including a marine phase before the elver stage (Sr/Ca ratio being highest), a brackish water phase (Sr/Ca ratio moder- ate), and a fresh water phase (Sr/Ca ratio lowest). The pattern of Sr/Ca ratios in the Group 4 otoliths is a com- bination of Group 1 and Groups 2 or 3. This pattern is clearly discernible in Sr X-ray maps (Fig. 3). These indicated that the environmental history of the eels could be distinguished from the time-series changes of the Sr/Ca ratios.
Sr/Ca ratios in otoliths of eels of unknown origin (Group 5)
Sr/Ca ratios measured along a transect from the pri- mordium to the otolith edge of the 3 unknown-origin eels collected away from Lake Malaren are shown in Fig. 4e. The pattern of Sr/Ca ratios in the first 2 eels (Nos. 25106 a n d 25108) was similar to those of Group 4
(No. 19315). The third one (No. 25110) was similar to Group 2. This pattern was clearly discernible in Sr X-ray maps (Fig. 3). This indicated that Nos. 25106 and 25108 experienced 3 different environments, while No. 251 10 experienced only 2; i.e. the former lived in brackish waters for a long period before moving to fresh water, while the latter moved to a fresh water environment at elver stage.
DISCUSSION AND CONCLUSION Advantage of X-ray map
The X-ray maps proved to be very useful although they are qualitative. The comparison of Sr patterns among otoliths is simple in the images, but rather dif- ficult in the data from the transects. Although we at- tempted to start our transects at the exact nucleus, in some cases the nucleus was not quantitatively analysed. The maps show that all otoliths had a high Sr area in their centers. In addition, they show that the deposition of Sr is approximately radially symmet- rical. This indicates that the direction chosen for tak- ing a quantitative transect is relatively unimportant and that a transect in any direction will cross all high and low Sr regions. However, the spacing of high and low Sr regions will vary depending on the direc- tion chosen, possibly complicating comparisons among transects.
Peak SrICa ratio in the otolith nucleus Regardless of whether the otoliths were removed from natural eels from brackish waters or stocked eels from fresh water lakes, all showed a peak Sr/Ca ratio (approximately 2 x 10-') in the nucleus at approxi- mately 100 pm from primordium. The ratios then decreased a n d maintained a lower level, approxi- mately 5.72 X 1 0 - ~ , for the eels from brackish waters and 3.7 X 10-4 for those from fresh water lakes (Fig. 4). Such peak ratios were previously reported in otoliths of Japanese eel elvers, and were proposed to corre- spond to the period when the eel metamorphosed from a leptocephalus to a glass eel (Otake et al. 1994, Tzeng & Tsai 1994). The leptocephalus is known to contain extensive amounts of gelatinous extracellu- lar matrix composed of sulfated glycosaminoglycans (GAG) which drastically decrease during the process of metamorphosis (Pfeiler 1984). GAG has a n affinity to alkali earth elements, especially to Sr (Nishizawa 1978). GAG decreases may reduce the absorption of Sr from sea water and result in a drastic decrease of otolith Sr content and, consequently, Sr/Ca ratios. The
early life history of European eels is analogous to that of Japanese eels (Schmidt 1925, Bertin 1956, Tesch 1983, Boetius & Harding 1985, McCleave & Kleckner 1985, Castonguay 1987, McCleave e t al. 1987). They pass through similar developmental stages during their migration from spawning grounds to estuaries. Accordingly, the mechanism of dramatic decrease of Sr/Ca ratios in the nuclei of the otoliths of European eel may b e similar to that of Japanese eel.
Environmental effect o n otolith Sr contents a n d SrICa ratios
Sr contents (or Sr/Ca ratios) in the otoliths were sig- nificantly different between brackish and fresh water (Group 1 vs Groups 2 a n d 3 in Fig. 4). The difference was also true for the otolith of a n individual eel that had both brackish a n d fresh water life history phases (Group 4 in Figs. 3 & 4 ) . Salinity was quite different between coastal brackish waters and inland lakes (Table 1). Sr contents in otolith of the fish were posi- tively correlated to ambient salinity (Kalish 1990. Lim- burg 1995, Tzeng 1996). Thus the difference in Sr content in otoliths of European eels between brackish and fresh water environments was probably d u e to salinity effects. A similar phenomenon has been ob- served in diadromous fishes a s the freshwater-marine gradient was traversed, e.g. migrating eels in the St. Lawrence, Canada (Casselman 1982), anadromous a n d non-anadromous salmonids in Tasmania (Kalish 1990), stri.ped bass in Chesapeake Bay, USA (Secor 1992), and Japanese eel in Taiwan (Tzeng & Tsai 1994). These studies indicated that salinity was an important factor influencing the Sr contents or Sr/Ca ratios in otoliths of the fish.
Several narrow bands of high Sr were found in the otoliths of the eels from Groups 1 a n d 4 (Fig. 3 ) . These bands were deposited while the eels lived in brackish water. We suggest that they w e r e deposited during a period of high salinity (Tzeng 1996) and low tempera- ture, a time when eel growth is slowed (Bruun 1963, Sinha & Jones 1967, Tzeng e t al. 1995). Sr contents (or Sr/Ca ratios) in fish otoliths have been negatively correlated to both temperature (Radtke 1984, 1989, Gauldie et al. 1986, Radtke et al. 1990, Townsend et al. 1992) and fish growth rates (Sadovy & Sevei-in 1992, 1994). All the eels in this study were exposed to periods of low temperature a n d , presumably, slow growth, whether they had lived in the fresh water lakes (Groups 2 and 3) or in brackish water (Groups 1 and 4). However, only the eels from brackish water had bands of high Sr in their otoliths (Groups 1 a n d 4, Fig. 3 ) . The fresh water eels from Groups 2 a n d 3 did not display any high Sr bands in their otoliths. From this
we conclude that salinity is more important in control- ling Sr incorporation into eel otoliths than is tempera- ture or growth rate.
Possible environmental history of g r o u p of u n k n o w n origin
T h r e e eels of unknown origin, Nos. 25106, 25108 a n d 25110, were collected from the eastern part of Lake Miilaren a few km away from the stocking locality. Their origins were proposed to be from (1) eels that w e r e stocked from elvers imported from France or British Isles, (2) eels that were stocked a t t h e yellow eel stage after being caught from the coastal waters of western Sweden, or ( 3 ) the natural (wild) eels which gradually migrated from the western coastal waters to the lakes on the eastern coast of Sweden.
T h e changes in Sr contents in the otolith of specimen No. 25110 were similar to those in Groups 2 a n d 3 , which were caught from fresh water lakes of eastern Sweden a n d stocked from imported elvers. Because the Baltic Sea is semi-enclosed, the possibility that eel larvae drifted with Atlantic currents a n d recruited directly to the east coast of Sweden is very low. Now- adays glass eels a r e not recruited to the Baltic Sea, except in the most southwestern parts (Svardson 1976) Most wild unstocked eels in the coastal waters of east- e r n Sweden must b e from the west coast a n d must have migrated through brackish waters before enter- ing fresh water lakes. If this were the case, the Sr con- tents in otolith No. 25110 should have several high Sr content bands a s do the otoliths in Group 1 (Fig. 3 ) . However, the Sr/Ca ratio in this otolith drastically decreased a t a n early stage. Accordingly, eel No. 25110 seems to be best assigned to Case ( l ) , which were released to fresh water a t the elver stage after being imported from France or the British Isles.
Similarly, Nos. 25106 a n d 25108 a r e best assigned to Case (2), because the maps of Sr contents a n d the time- series changes in Sr contents or Sr/Ca ratios in them were similar to those of Group 4 (Figs. 3 & 4 ) , which were stocked into the fresh water lakes a t the yellow e e l stage after being collected from domestic brackish waters; or possibly C a s e (3), because the maps of Sr contents a n d the time-series c h a n g e in Sr/Ca ratios in otolith of the eels exhibited a similar geographic decline in salinity gradient from western to eastern coasts of Sweden. However C a s e (2) is most likely because the c h a n g e in Sr contents or Sr/Ca ratios w a s very rapid. To verify Case (3), it would b e necessary to examine the Sr contents in otoliths of European eels around the coastal waters of S w e d e n .
In conclusion, Sr X-ray maps a n d Sr/Ca ratios of the otoliths were significantly different between eels col-
80 Mar Ecol Prog Ser 149: 73-81, 1997
Lepto- glass eel
cephalus or elvers yellow eel
<
..-/./
1.
>
Metamorphosis
Life history
Fig. 5 . Schematic diagram of the environmental history of the studied eel as Indicated by the time series change of Sr/Ca
ratios in otoliths
l e c t e d f r o m b r a c k i s h and f r e s h w a t e r e n v i r o n m e n t s , and c o u l d b e u s e d as a n e n v i r o n m e n t a l i n d i c a t o r to d e t e c t t h e i r m i g r a t o r y history b e t w e e n s e a w a t e r a n d f r e s h w a t e r . A s c h e m a t i c d i a g r a m of t h e m i g r a t o r y history of t h e e e l s i n this s t u d y is s h o w n in Fig. 5.
Acknowledgements. This cooperative study was conducted
with financial support from the National Science Council,
Taiwan (Project No. NSC 84-231 1-B-002-035 and 86-231 1-B- 002-042). The authors are grateful to Dr H. Westerberg and
the Swedish Meteorological and Hydrological Institute for
supplying oceanographic data, Miss Y C. Tsai for preparing otolith specimens for WDX analysis, Miss C . E. Wu for typing
the manuscript, and Dr R. W. Gauldie and 3 anonymous
reviewers for reviewing the manuscript.
LITERATURE CITED
Bertin L (1956) Eels-a biological study.Cleaver-Hume Press, London
Boetius J. Harding EF (1985) A re-examination of Johannes Schmidt's Atlantic eel investlgations. Dana 4.129-162 Bruun AF (1963) The breedlng of the north Atlantic fresh-
water eel. Adv Mar Biol 1:137-169
Casselman J M (1982) Chemical analyses of optically different zones in eel otoliths. In: Loftus KH (ed) Proceedings of the 1980 North American Eel Conference. Ontario Fish Tech Rep Ser 4 , p 74-82
Castonguay M (1987) Growth of American and European eel leptocephali as revealed by otolith m~crostructure. Can J 2001 65:875-878
Erichsen L (1976) Statistik over alyngeluppsamling i svenska vattendrag. Inform Inst Freshwat Res, Drottningholm 8: 1-36 (in Swedish)
Filuk J , Wiktor J (1988) Management of eel stocks in the Vistula and Szczecin lagoons in the light of the process of eel stocking. Biul Morsk Inst Ryb 5-6:36-44
Gauldie RW, Fournier DA, Dunlop DE, Coote G (1986) Atomic emission and microprobe studies of the ion content of otoliths of Chinook salmon aimed at recovering the tem- perature life history of individuals. Comp Biochem Physiol 84A(4) 607-615
Hagstrom 0, Wickstrom H (1990) Immigration of young eels to the Skagerrak-Kattegatt area 1900 to 1989. Int Revue
Hydrobiol 75(6):707-716
Holmgren K, Wickstrom H (1988) The quality of Swedish yellow eels used for stocking in 1987-a study of sex, size a n d wounds. Inform Inst Freshwat Res. Drottningholm 8, 1-38 (in Swedish w ~ t h English summary).
Kalish JM (1990) Use of otolith microchemi.stry to distin-
guish the progeny of sympatric anadromous and non- anadromous salmonlds. Fish Bull US 88:657-,666
Leopold M (1976) Stocking as a main factor determining the level of eel catches in Poland. Int Counc Explor Sea (EIFAC) Symp Eel Res Mgmt 14:l-13
Limbu.rg KE (1995) Otolith strontium trace environmental history of subyearling American shad Alosa sapidissima. Mar Ecol Prog Ser 119:25-35
McCleave JD, Kleckner RC (1985) Oceanic migrations of Atlantic eels (Anguilla spp.): adults and their offspring.
Contr Mar Sci 27(Suppl):316-337
McCleave JD, Kleckner RC, Castonguay M (1987) Reproduc- tive sympatry of American and European eels and impli- cations for migration and taxonomy. Am Fish Soc Symp 1:286-297
Nishizawa K (1978) Marine algae from a viewpo~nt of phar-
maceutical studies. J a p J Phycol 26:73-78
Odum HT (1957) Biochemical deposition of strontium. Texas Univ Inst Mar Sci 4:39-114
Otake T, Ishii T, Nakahara M , Nakamura R (1994) Drastic
changes in otolith strontium/calcium ratios in leptocephali and glass eels of Japanese eel Anguilla japonlcd. Mar Ecol
Prog Ser 12:189-193
Pfeiler E (1984) Glycosaminoglycan breakdown during meta-
morphosis of larval bone fish Albula. Mar Biol Lett 5: 241-249
Radtke RL (1984) Cod otoliths: ~nformation storage structures.
In: Dahl E, Danielssen DS, Moksness E, Solendal P (eds)
The propagation of cod Gadus morhua L. Fladevigen
Rapp 1, p 273-298
Radtke RL (1989) Strontium-calcium concentration ratios in fish otoliths as environmental indicators. Comp Biochem
Physiol 92(A):189-193
Radtke RL, Kinzie I11 RA, Folsom SD (1988) Age at recruit- ment of Hawaiian freshwater gobies. Environ Biol Fish 23: 205-213
Radtke RL, Townsend DW, Folson SD, Morrison MA (1990) Strontium:calcium concentration ratios in otoliths of her- ring larvae as indicators of environmental h~stories. Envi- ron Biol F ~ s h 27:51-61
Sadovy Y, Severin KP (1992) Trace element in biogenic arag-
onite: correlation of body growth rate and strontium levels
in the otoliths of the white grunt, Haernulon plumieri (Pisces:Haemulidae). Bull Mar Sci 50:237-257
Sadovy Y, Severin KP (1994) Elemental patterns in red hind
(Epinephelus guttatus) otoliths from Bermuda and Puerto
Rico reflect growth rate, not temperature. Can J Fish Aquat Sci 51:133-141
Schmidt J (1925) The breeding places of the eel. Ann Rep Smithson Inst 1924:279-316
Secor DH (1992) Application of otolith microchemistry analy- sis to investigate anadromy in Chesapeake Bay striped bass Morone saxatilis. Fish Bull US 90:798-806
Sinha VRP, Jones JW (1967) On the a g e and growth of the
freshwater eel (Anguilla anguilla). J Zoo1 Soc Lond 153:
99-117
Svardson G (1976) The decline of the Baltic eel population.
Rep Inst Freshwat Res 55136-143
Tesch FW (1983) Der Aal: Biologie und Fischerei, Vol. 2. Parey, Hamburg
tium:calcium ratios In juvenile Atlantic herring Clupea harengus L. otoliths a s a function of water temperature. J Exp Mar Biol Ecol 160:131-140
Tzeng WN (1995) Migratory history recorded in otoliths of the Japanese eel, .4nguilla japonlca, elvers a s revealed from SEM and WDS analyses. Zool Stud 34(SuppI 1):234-236 Tzeng LVN (1996) Effect of salinity and ontoyenetic move-
ment on Sr/Ca ratio in otolith of Japanese eel. J Exp Mar Biol Ecol 199 111-122
Tzeng W N , T s a ~ YC (1994) Changes In otolith ~nlcrochem~stry of the J a p a n e s e eel, Angullla jdponlca, d u r ~ n g its n?igr,~- tlon from the ocean to the rlvers of Taiwan. J Fish B1ol45: 67 1-684
This article was s u b n ~ i t t e d to the editor
Tzeng WN, Wu HF, Wickstrijm H (1995) S c a n n ~ n g electron lnicroscopic analysis of annulus microstructure in otollth of European eel, Anguilla anguilla. J Fish Biol 45:479-492 Westin L (1990) Orientation mechanisms in migrating Euro- pean silver eel (Anguilla a n g ~ ~ i l l a ) : temperature and olfac- t ~ o n . h4ar Biol 106:175-179
LV~ckstro~n H (1984) T h e S w e d ~ s h eel stockmy programme. EIFAC/FAO Techn Pap 42 (Suppl Vol 1 ) 68-83
Wlckstrorn H (1986) Growth of cultured eels stocked in two S w e d ~ ~ h lakes. Vie h4ilieu 36 273-277
'Vlll~an~s K L (19871 An introduct~on to X-ray spectrometry. X-ray fluorescence a n d electron mlcroprobe analysis. Allen & Unwin. Boston
Manuscript first r e c e ~ v e d : M ~ f y 15, 1996 R r ~ , i s e d version accepted: J a n u a r y 9, 1997