Brazilein suppresses migration and invasion of MDA-MB-231 breast
cancer cells
Chi-Ying Hsieh
a, Pei-Chien Tsai
a, Chiao-Lun Chu
a, Fang-Rong Chang
b, Long-Sen Chang
c,
Yang-Chang Wu
d,e,f,⇑, Shinne-Ren Lin
a,⇑a
Department of Medicinal and Applied Chemistry, Kaohsiung Medical University, Kaohsiung 807, Taiwan b Graduate Institute of Natural Products, Kaohsiung Medical University, Kaohsiung 807, Taiwan c Institute of Biomedical Sciences, National Sun Yat-Sen University, Kaohsiung 804, Taiwan d
School of Pharmacy, College of Pharmacy, China Medical University, Taichung 404, Taiwan
e Chinese Medicinal Products Research and Development Center, China Medical University Hospital, Taichung 404, Taiwan f Center for Molecular Medicine, China Medical University Hospital, Taichung 404, Taiwan
a b s t r a c t
Brazilein, a bioactive compound isolated from Caesalpinia sappan L., has long been used in oriental folk medicines. Cancer metastasis is a primary cause of cancer death. However, the anti-metastatic effects of brazilein remain elusive. In this study, we found that brazilein inhibited human breast cancer MDA-MB-231 cell migration and invasion using wound-healing assay and Boyden chamber assay. The results of Western blot, gelatin zymography and reversed transcription-PCR analysis showed that brazilein sup-pressed matrix metalloproteinase-2 (MMP-2) expression in a concentration-dependent manner. Brazilein also decreased the nuclear protein level of nuclear factor kappaB (NF-jB). Brazilein potently suppressed the phosphorylation of p38 mitogen-activated protein kinase (p38 MAPK), phosphatidylinositide-3-kinase (PI3K) and Akt, but did not affect phosphorylation of extracellular signal regulating phosphatidylinositide-3-kinase (ERK)1/2 and c-Jun N-terminal kinase (JNK). Additionally, treatment of SB203580 (p38 MAPK inhibitor) or wortmannin (PI3K inhibitor) resulted in a reduced activity and expression of MMP-2 as well as inhi-bition on cell migration and invasion in MDA-MB-231 cells. Taken together, these results suggest that brazilein inhibition of MDA-MB-231 cells may be mediated through inactivation of both PI3K/Akt and p38 MAPK signaling pathways, leading to inhibitory effect on NF-jB activation. Consequently, brazilein suppresses MMP-2 expression, and thus confers anti-migration and anti-invasion of MDA-MB-231 cells.
.
1. Introduction
Breast cancer ranks among the most common malignant tumors afflicting women worldwide [1]. Cancer metastasis is the leading cause of mortality in patients with breast cancer. Metastasis is multistep process, which includes detachment of cancer cells from primary tumor, migration, adhesion and invasion of cancer cells into the blood or lymphatic vessels. The help of matrix metallopro-teinases (MMPs) is required for extravasation out of the vessel, leading to the movement of cancer cells to reach the target tissue. Among human MMPs, MMP-2 and MMP-9 are key enzymes in the degradation of type IV collagen, which is an important component of extracellular matrix (ECM) [2,3]. Clinical and experimental stud-ies have shown that MMP-2 and MMP-9 are involved in tumor metastasis and their elevated expression has been associated with poor prognosis [3,4]. In breast cancer, both gelatinases are
ex-pressed, and thus it has been suggested that they play an impor-tant role in cancer metastasis [2,3,5].
In addition to MMPs, the mitogen-activated protein kinase fam-ily members (MAPKs) are also known to be involved in metastasis. The three major mammalian MAPKs include extracellular signal-regulated kinases 1 and 2 (ERK1/2), c-Jun N-terminal kinase/ stress-activated protein kinase (JNK/SAPK), and p38 MAPK [6,7]. PI3K/Akt signaling pathway regulates cell metastasis and is closely associated with the development and progress of various tumors [8,9]. Noticeably, PI3K/Akt and MAPK signaling pathways play a crucial role in regulating the expression of MMPs via nuclear-fac-tor-kappaB (NF-
j
B) [10]. NF-j
B is maintained in the cytoplasm through interaction with an inhibitor of NF-j
B (Ij
B). However, upon dissociation, NF-j
B moves into the nucleus and exerts its transcriptional function. Given that NF-j
B has been reported to be involved in regulating MMP-2 expression, suppression ofPI3K/Akt and MAPK pathways as well as NF-
j
B activation may be feasible mechanisms for reducing metastasis.Brazilein, isolated from the dried heartwood of Caesalpinia sap-pan L., has long been used in traditional medicine in China. It shows pharmacological effects such as antimicrobial, anti-inflam-matory, anti-oxidative and anti-atherosclerosis properties [11]. Moreover, brazilein exhibits cytotoxic activity on HepG2 and Hep3B (liver), MDA-MB-231 and MCF-7 (breast), A549 (lung), and Ca9-22 (gingival) human cancer cell lines [12]. Treatment of human hepatocellular carcinoma HepG2 cells with brazilein led to significant decrease in survivin protein expression, which was associated with a strong and prominent caspase-9 and -3 activa-tion as well as PARP cleavage [13]. MDA-MB-231 breast cancer cells, a highly metastatic human breast carcinoma cell line, are de-rived from a metastatic plural effusion fluid and are widely used as a model for studying metastatic cell migration [14]. In terms of that the capacity of brazilein to inhibit the migration and invasion of malignant carcinoma is unclear, the objective of the study was to explore whether brazilein inhibited the migration and invasion of MDA-MB-231 cells, and to further elucidate the underlying mechanism.
2. Materials and methods
2.1. Materials
A Trizol RNA isolation kit was obtained from Life Technolo-gies (Rockville, MD, USA). Primers for RT-PCR, dNTP, reverse transcriptase, Taq polymerase, phosphate-buffered saline (PBS), trypsin-EDTA, and DMEM/F-12 medium were purchased from Gibco BRL (Gaithersburg, MD). Matrigel was obtained from BD transduction Laboratories (San Diego, CA). The following antibod-ies were used for Western blotting: anti-ERK, JNK, p38, Akt, phospho-ERK, phospho-JNK, phospho-p38, phospho-Akt, I
j
Ba
, IKK, C23, and b-actin (Santa Cruz Biotechnology, Santa Cruz, CA); anti-phospho-IKK and phospho-IKB (Cell Signaling Technol-ogy, Beverly, MA); anti-PI3K (p85), phospho-PI3K (p85), NF-j
B (p65), MMP-9, and MMP-2 (Millipore, Beverly, MA). Specific inhibitors of PI3K (wortmannin), p38 MAPK (SB203580), and po-tent selective inhibitor of MMP-2 (MMP-2I) were purchased from Calbiochem (San Diego, CA). Anti-mouse, anti-Goat and anti-rabbit IgG peroxidase-conjugated secondary antibodies were purchased from Pierce (Rockford, IL). The enhanced chemilumi-nescence (ECL) kit was purchased from Amersham Life Science (Amersham, UK).2.2. Cell culture
The human breast cancer cell line, MDA-MB-231, was obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). Cells were maintained in DMEM/F12 medium supplemented with 10% fetal calf serum, 2 mM glutamine, and antibiotics (100 U/ mL penicillin and 100
l
g/mL streptomycin) at 37 OC in a humidi-fied atmosphere of 5% CO2.2.3. Isolation of brazilein
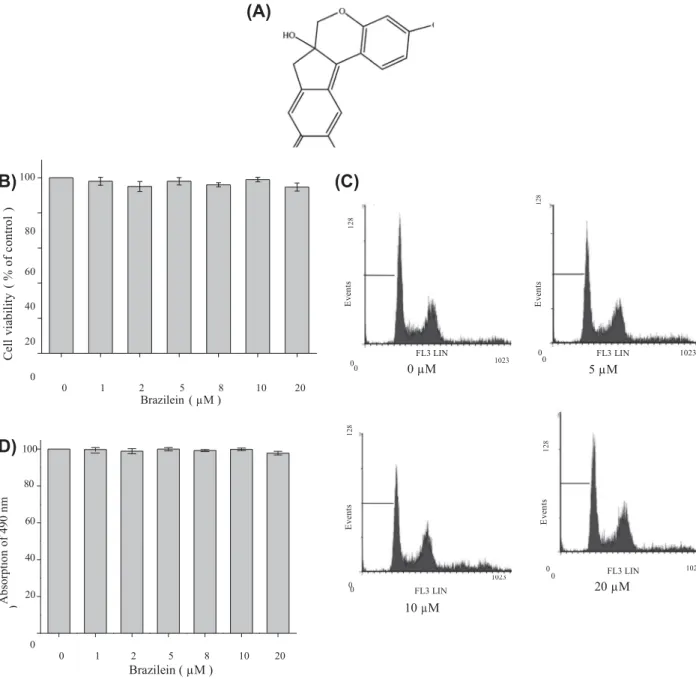
Brazilein (Fig. 1A) was isolated from an ethyl acetate extract of Caesalpinia sappan L. (Legminosae) and can be synthesized by a chemical process as described previously [12,15]. The purity of the brazilein is over 98% based on the high performance liquid chromatography analysis. Brazilein was dissolved in DMSO (less than 0.01%) and made immediately prior to experiments.
2.4. Cell growth inhibition assay
The viability of cells was determined by the MTT assay and the Trypan blue dye exclusion assay was performed to confirm and verify cell viability. After 24 h of incubation, the cells were treated with vehicle (0.1% DMSO) or a series of concentrations of brazilein for 24 h. MTT solution was added to each well (1.2 mg/mL) and incubated for 4 h. The MTT-formazan product dissolved in DMSO was estimated by measuring absorbance at 570 nm in an ELISA plate reader. For the Trypan blue dye exclusion assay, cells were seeded at a density of 1 x 105 cells/well onto a 12-well plate for
24 h, then brazilein was added to the medium at indicated times and concentrations. After incubation, cells exposed to 0.2% Trypan blue were counted in a hemocytometer.
2.5. Cell cycle analysis
Cells were harvested, washed, and resuspended as described previously [16]. Controlled and treated cells were harvested, washed in cold phosphate-buffered saline (PBS), fixed in 70% etha-nol, and stored at 4 OC. DNA was treated with RNase. A solution (500 U/mL) at 37 OC for 15 min and stained by propidium iodide (50
l
g/mL) in 1.12% sodium citrate at room temperature before analysis. Flow cytometric determination of DNA content was done by Coulter EPICS XL Flow Cytometer (Coulter Corp., Miami, FL, USA). The fractions of the cells in G0/G1, S, and G2/M phase wereanalyzed using Multicycle (Phoenix flow system, San Diego, CA, USA) cell cycle analysis software.
2.6. MTS proliferation assay
3-(4,5-Dimethylthiazol-2yl)-5-(3-carboxymethoxyphenyl)-2(4-sulfophenyl)-2H-tetrazolium (MTS) assays were done as described by the supplier (Promega, Madison, WI). Briefly, MDA-MB-231 cells were seeded at a density of 5 x 103 in 96-well plates, and treated
with indicated concentrations of brazilein. After incubation for 24 h, MTS reagent was added, and the absorption was measured at 490 nm.
2.7. Wound-healing assay
Cells were grown to 90% confluence in a 12-well plate at 37 OC, 5% CO2 incubator. The monolayers were scratched with a plastic
tip, washed by PBS to remove floating cell debris, and then incu-bated in medium in the absence or presence of brazilein for 24 h. Cell migration into the wound surface was determined under an inverted microscope. Migrated cells across the black lines were counted in six random fields from each triplicate treatment, and the data were presented as mean ± SD.
2.8. Cell invasion assay
The ability of MDA-MB-231 cells for passing through Matrigel-coated filters was measured by the Boyden chamber invasion as-say. In brief, cells were treated with various concentrations of bra-zilein. After 24 h, cells were detached by trypsin and resuspended in serum-free medium. Medium containing 10% FBS was applied to the lower chamber as chemoattractant and then the cells were seeded on the upper chamber at a density of 1 x 105 cells/well in
a 50
l
L of serum-free medium. The chamber was incubated at 37 OC for 24 h. At the end of incubation, the cells in the upper sur-face of the membrane were carefully removed with a cotton swab. Cells invaded across the Matrigel to the lower surface of the mem-brane were fixed with methanol and stained with hematoxylin and eosin. The invasive cells on the lower surface of the membrane fil-ter were counted with a microscope.(A)
(B)
100(C)
80 Sub G1 3.0% Sub G1 3.2% G0/G1 41.2% G0/G1 41.9% 60 S M1 G2/M 40 25.9% 29.9% S 25.5% M1 G2/M 29.4% 20 00 0 0 1 2 5 8 10 20 Brazilein ( µM ) FL3 LIN 0 µM 0 1023 0 FL3 LIN 5 µM 1023(D) 100
Sub G1 3.0% G0/G1 42.0% Sub G1 3.1% G0/G1 42.1% 80 S 25.4% S 25.6% M1 G2/M 29.6% 60 M1 G2/M 29.2% 40 0 20 0 FL3 LIN 10 µM 0 1023 0 FL3 LIN 1023 20 µM 0 0 1 2 5 8 10 20 Brazilein ( µM )Fig. 1. (A) Chemical structure of brazilein. (B) Effect of brazilein on the viability in MDA-MB-231 cells. Cells were treated with various concentrations (0, 1, 2, 5, 8, 10, and 20 lM) of brazilein for 24 h. Cell viability was determined by the MTT assay. The cell viability was directly proportional to the production of formazan, which was measured spectrophotometrically at 570 nm. Values are expressed as mean ± SD of three independent experiments. ⁄P < 0.05, compared with the untreated control. (C) Cell cycle analysis of brazilein-treated cells. Cells were treated with various concentrations (0, 5, 10 and 20 lM) of brazilein for 24 h. The cells were fixed and stained with PI, and the cell cycle distribution was then analyzed by flow cytometry. The data indicate the percentage of cells in sub-G1, G0/G1, S, and G2/M phases of the cell cycle. Data was presented from three independent experiments. (D) The effect of brazilein on MDA-MB-231 cell proliferation. Cells were treated with indicated concentrations of brazilein, incubated
for 24 h, and evaluated for cell proliferation by MTS assay. The experiments were independently repeated three times.
2.9. Determination of MMP-2 and MMP-9 by Gelatin zymography assay
The enzymatic activities of MMP-2 and MMP-9 were deter-mined by gelatin zymography. Briefly, conditioned media were prepared with standard SDS-gel loading buffer containing 0.01% SDS without b-mercaptoethanol and not boiled before loading. Then, prepared samples were subjected to electrophoresis with 8% SDS polyacrylamide gels containing 0.1% gelatin. After electro-phoresis, gels were washed twice with 100 mL distilled water containing 2% Triton X-100 for 30 min at room temperature to re-move SDS and then incubated in 100 mL reaction buffer (40 mM Tris–HCl, pH 8.0, 10 mM CaCl2, 0.02% NaN3) for 24 h at 37 OC,
stained with Coomassie brilliant blue R-250 and destained with destaining solution (20% methanol, 10% acetic acid, and 70%
water). 12 8 C el l vi ab ili ty ( % o f co nt ro l ) 12 8 E ve nt s E ve nt s 12 8 12 8 E ve nt s A bs or pt io n of 4 90 n m (% o f co nt ro l) E ve nt s
2.10. Preparation of whole-cell lysates and nuclear extracts
The cells were lysed with iced-cold RIPA buffer (1% NP-40, 50 mM Tris-base, 0.1% SDS, 0.5% deoxycholic acid, and 150 mM NaCl, pH 7.5), and then phenylmethylsulfonyl fluoride (10 mg/mL), leupeptin (17 mg/mL), and sodium orthovanadate (10 mg/mL) were added. The samples were mixed for 30 min on ice and then centrifuged at 14,800 rpm for 30 min. Then, the super- natants were collected, denatured, and subjected to SDS– PAGE and Western blotting. Nuclear extracts were prepared as follows. The
cells were lysed with 500
l
L of lysis buffer (50 mM KCl, 25 mM Tris, 0.5% NP-40, 1 mM phenylmethylsulfonyl fluoride, 10l
g/mL of leupeptin, 20l
g/mL of aprotinin, and 100l
M DTT). After centrifugation at 2000 rpm for 10 min, the nuclear pellet was resuspended in nuclear extract buffer (1.5 mM MgCl2, 10 mMphenylmethylsulfonyl fluoride, 25% glycerol, and 420 mM NaCl). The nuclear suspension was incubation on ice for 20 min and then centrifuged at 14,800 rpm for 30 min. The supernatant (corre-sponding to the soluble nuclear fraction) was saved, and then used for protein detection.
2.11. Western blot analysis
(A)
0 µM 5 µM 10 µM 20 µM
Briefly, cells were lysed with RIPA buffer containing protease inhibitor. Equal amounts of protein (50
l
g) were separated by SDS–polyacrylamide gel electrophoresis and then were electro-transferred to PVDF membrane. The membrane was blocked with a solution containing 5% nonfat dry milk TBST buffer (20 mM Tris–HCl, pH 7.4, 150 mM NaCl and 0.1% Tween 20) for 1 h and washed with TBST buffer. The indicated primary antibodies were incubated, washed, and monitored by immunoblotting using spe-cific antibodies. These proteins were detected by enhanced chemiluminescence. 100 80 60*
40*
20 0 2.12. Isolation of total RNA, reverse transcriptase polymerase chainreaction (RT-PCR), and DNA electrophoresis
Total RNA was isolated from MDA-MB-231cells using Trizol re-agent according to supplier’s protocol. Total RNA (2
l
g) was tran-scribed to 20l
L of cDNA with 1l
L of dNTPs (2.5 mM), 1l
L of Oligo dT (10 pmol/l
L), and 1l
L of RTase (200 U), 1l
L of RNase inhibitor, and 5x reaction buffer. The appropriate primers were as followed: sense of MMP-2, 50 -GGCCCTGTCACTCCCTGAGAT-30nt 1337–1356; antisense of MMP-2, 50 -GGCATCCAGGTTATCGGG
GA-30 nt 2026–2007; sense of MMP-9, 50 -AGGCCTCTACAGAG
TCTTTG-30 nt 1201–1220; antisense of MMP-9, 50 -CAGTCC
AACAAGAAAGGACG-30 nt 1702–1683; sense of GADPH, 50 -CG
GAGTCAACGGATTGGTGTT-30 nt 94–126; antisense of GADPH,
50 -AGCCTTCTCCATGGTTGGTGAAGAC-30 nt 399–375) were used
for PCR amplifications. PCR was performed under the following conditions: 30 cycles at 94 OC for 1 min, 50 OC for 1 min, 72 OC for 2 min and followed by a final incubation at 72 OC for 10 min. PCR products were analyzed by 1% agarose gel and visualized by ethi- dium bromide staining.
(B)
0 5 10 20 Brazilein (μM) 0 µM 5 µM 10 µM 20 µM 2.13. Inhibitor treatmentTo investigate the effect of MAPK and PI3K inhibitors on brazil-ein-induced cell migration and invasion, confluent cell culture was preincubated for 1 h with one of the following inhibitors before the addition of brazilein: 10
l
M SB203580 (a p38 MAPK inhibitor) or 10l
M wortmannin (a PI3K inhibitor).2.14. Statistical analysis
Data were reported as the mean ± SD of three independent experiments done in triplicate. Statistical analysis between two samples was performed using Student’s t- test. Statistical compar-isons of more than two groups were performed using one-way analysis of variance (ANOVA) with Bonferroni’s post hoc test. In all cases, p < 0.05 was considered as significant.
3. Results
3.1. Cytotoxicity of brazilein to MDA-MB-231 cells
The chemical structure of brazilein is shown in Fig. 1A. To verify the effect of brazilein on cell viability, cells were treated with bra-zilein at indicated concentrations for 24 h. Compared with that of
100
*
80 60 40*
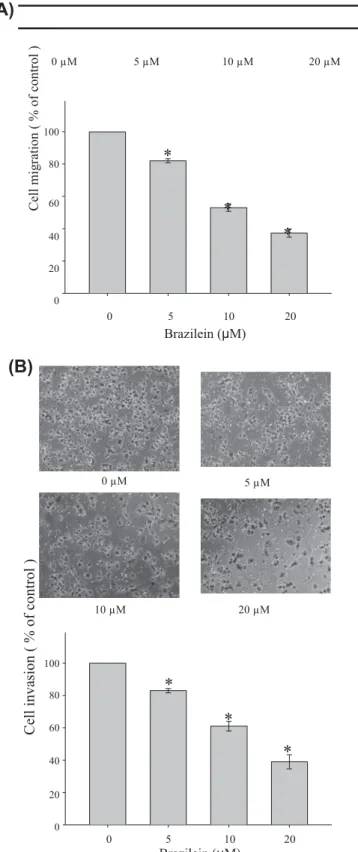
20 0 0 5 10 20 Brazilein (μM)Fig. 2. (A) Effect of brazilein on the migration of MDA-MB-231 cells. In the wound-healing assay, cell monolayers were scratched by a sterile micropipette tip and the cells were treated with various concentrations of brazilein for 24 h. The number of cells in the denuded zone was quantified by inverted microscope. Black lines indicate the wound edge. Migrated cells across the black lines were counted in six random fields from each treatment, and the data were presented the mean ± SD of triplicate wells from three independent experiments. ⁄P < 0.05, compared with the untreated control. (B) Effect of brazilein on the invasion of cells. In the Boyden chamber invasion assay, cells were treated with various concentrations of brazilein for 24 h and the invasion ability of cells was quantified by counting the number of cells that invaded the underside of the porous polycarbonate membrane under microscope. Values are expressed as mean ± SD of three independent experiments. ⁄P < 0.05, compared with the untreated control.
*
C el l m ig ra ti on ( % o f co nt ro l )C
el
l i
nv
as
io
n
(
%
o
f
co
nt
ro
l )
*
(A)
MMP-9 Brazilein (µM) 0 5 10 20 92 kDa(B)
MMP-9 Brazilein (µM) 0 5 10 20 92 kDa MMP-2 72 kDa GAPDH MMP-2 GAPDH 72 kDa 100 80 60 40 20*
MMP-9*
MMP-2*
100 80 60 40 20 0*
MMP-9 MMP-2*
0 0 5 10 20 0 Brazilein (μM)5 10 20 Brazilein (μM)(C)
MMP-9
MMP-2
GAPDH
Brazilein (µM) 0 5 10 20(D)
100 80 60 * * 40 * 100 80 60 40 20 0*
*
MMP-9 MMP-2*
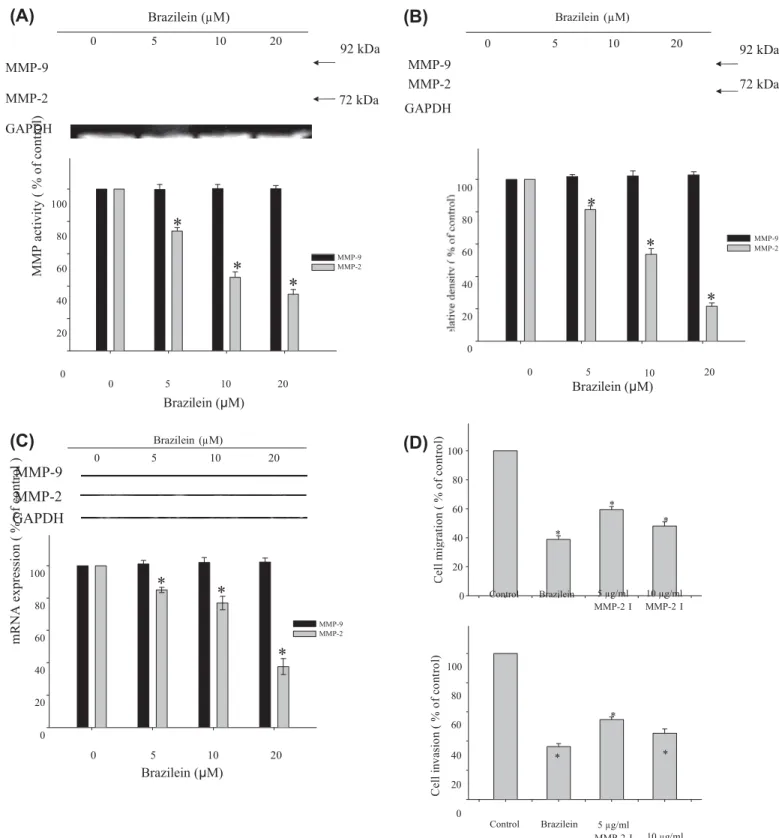
20 0 100 80 60 Control Brazilein 5 µg/ml MMP-2 I * 10 µg/ml MMP-2 I 0 5 10 20 Brazilein (μM) 40 * 20 0 Control Brazilein 5 µg/ml MMP-2 I * 10 µg/ml MMP-2 IFig. 3. Effect of brazilein on the activities and expression levels of MMPs in MDA-MB-231 cells. Cells were treated with various concentrations (0, 5, 10, and 20 lM) of brazilein for 24 h, and then subjected to (A) gelatin zymography to analyze the activities of MMP-2 and MMP-9. The conditioned media were collected and MMP-2 and
MMP-9 activities were determined. GAPDH was used as loading control. The activities of MMP-2 and MMP-MMP-9 were quantified by densitometric analysis. (B) Western blotting to analyze the protein levels of MMP-2 and MMP-9. GAPDH served as an internal control of protein level. The relative density of MMP-2 and MMP-9 was normalized to GAPDH, which was determined by densitometric analysis. The values represented as the mean ± SD of at least three independent experiments (⁄p < 0.05). (C) Cells were treated with various concentrations of brazilein for 24 h, and then RNA samples were extracted and subjected to a semi-quantitative RT-PCR for MMP-2 and MMP-9 with GADPH being an
internal control. The PCR products were quantified by densitometric analysis with that of the untreated group being 100% as shown just below the gel data. The values are expressed as mean ± SD of three independent experiments. ⁄P < 0.05, compared with that of the untreated control. (D) Effect of MMP-2 inhibitor, MMP-2I, on the cell migration and invasion in MDA-MB-231 cells. Cells were pretreated with MMP-2I (5 lg/mL or 10 lg/mL) for 1 h and then incubated for 24 h. Later, the cells were subjected to analyze for cell migration and invasion, as described in Section 2. Data are expressed as mean ± SD of three independent experiments. ⁄P < 0.05, compared with the untreated control. M M P a ct iv it y ( % o f co nt ro l)
*
C el l m ig ra ti on ( % o f co nt ro l) m R N A e xp re ss io n ( % o f co nt ro l ) C el l i nv as io n ( % o f co nt ro l)NF-κB C23 p-IKK IKK p-IKB α IKB α β-Actin Brazilein (µM) 0 5 10 20
Nuclear
NF-κB 100 80*
60 40*
20 0*
0 5 10 20 Brazilein ( µM )Cytosolic
100 p-IKK 100 IKK 80 80*
60 60 40*
40*
20 20*
0 0 5 10 20 Brazilein ( µM ) 0 0 5 10 20 Brazilein ( µM ) 100 p-IKBα IKBα 300*
*
80 60 40*
20*
*
0 200*
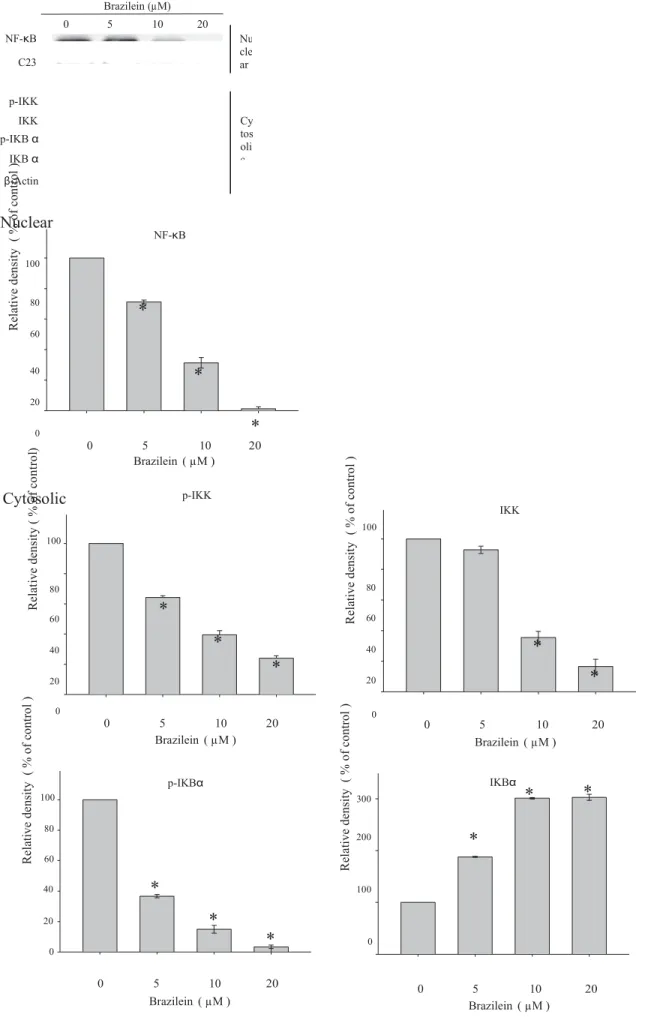
100 0 0 5 10 20 Brazilein ( µM ) 0 Brazilein ( µM )5 10 20Fig. 4. Inhibitory effects of brazilein on the expression of NF-jB in MDA-MB-231 cells. Cells were treated with various concentrations (0, 5, 10 or 20 lM) of brazilein for 24 h and then nuclear extracts (NF-jB) and cytosolic extracts (p-IKK, IKK, p-IjBa and IjBa) were subjected to SDS–PAGE following by Western blot. C23 and b-actin were served as a nuclear and cytosolic internal control, respectively. Signals of proteins were visualized with an ECL detection system. The relative density of these proteins was
Nu cle ar Cy tos oli c R el at iv e de ns ity ( % o f co nt ro l ) R el at iv e de ns it y ( % o f co nt ro l) R el at iv e de ns ity ( % o f co nt ro l )
*
R el at iv e de ns ity ( % o f co nt ro l ) R el at iv e de ns ity ( % o f co nt ro l )normalized to C23 or b-actin, respectively, which was determined by densitometric analysis. Data represented the mean ± SD of three independent experiments performed in triplicate (⁄p < 0.05).
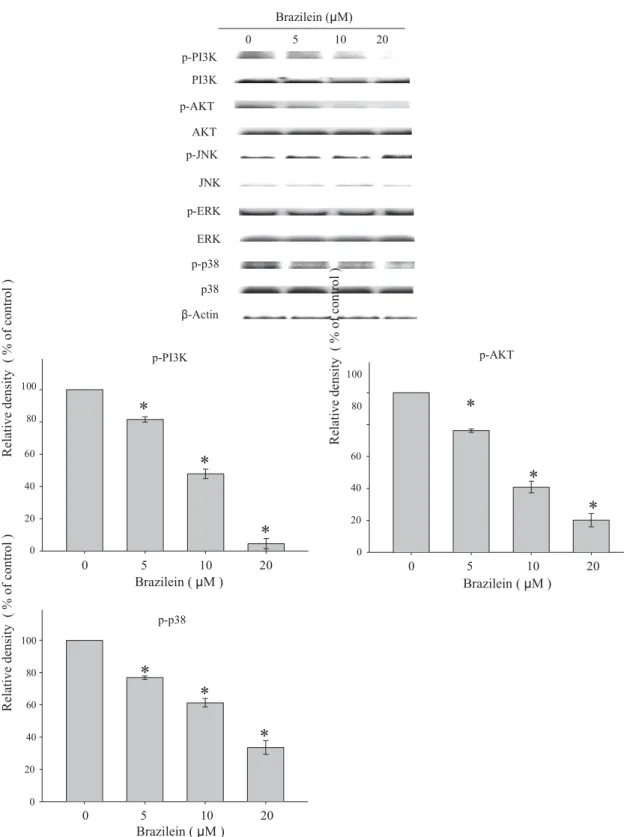
p-PI3K PI3K p-AKT AKT p-JNK JNK p-ERK ERK p-p38 p38 β-Actin Brazilein (μM) 0 5 10 20 100 80 p-PI3K
*
100 80 p-AKT*
60 60*
40 40*
20 20*
0 0 0 5 10 20 0 5 10 20 Brazilein ( μM ) Brazilein ( μM ) 100 80 60 40 p-p38*
*
*
20 0 0 5 10 20 Brazilein ( μM )Fig. 5. Effects of brazilein on the phosphorylation of MAPKs and PI3K/Akt signalings. Cells were treated with 0, 5, 10 or 20 lM of brazilein for 24 h and then the cell lysates were subjected to SDS–PAGE followed by Western blots with anti-ERK1/2, anti-p38, anti-JNK1/2, anti-PI3K, and anti-Akt (total and phosphorylated) antibodies as described in Section 2. b-Actin was used as loading control. The relative density of these proteins was normalized to b-actin, which was determined by densitometric analysis. Data represented the mean ± SD of three independent experiments done in triplicate (⁄p < 0.05).
untreated control cells, the viability of MDA-MB-231 cells was not significantly affected by treating with brazilein at a concentration between 0 and 20
l
M (Fig. 1B). In order to ascertain that the doses applied in all subsequent experiments did not affect cell growth, the effects of brazilein on cell cycle progression were also analyzed by flow cytometry. Brazilein did not change the proportion of MDA-MB-231 cells in G0/G1, S, and G2/M phases. Furthermore,bra-zilein did not induce an increase in the proportion of sub-G1 peaks,
indicating the absence of apoptotic cells (Fig. 1C). Furthermore, cel-lular proliferation inhibitory potency of brazilein was studied using MTS assay, the treatment of brazilein (1–20
l
M) to MDA-MB-231 cells did not show inhibitory effect on cell proliferation (Fig. 1D). Thus, non-cytotoxic concentrations of brazilein (5–20l
M) were used for subsequent experiments.R el at iv e de ns it y ( % o f co nt ro l ) R el at iv e de ns it y ( % o f co nt ro l )
*
R el at iv e de ns it y ( % o f co nt ro l )(A)
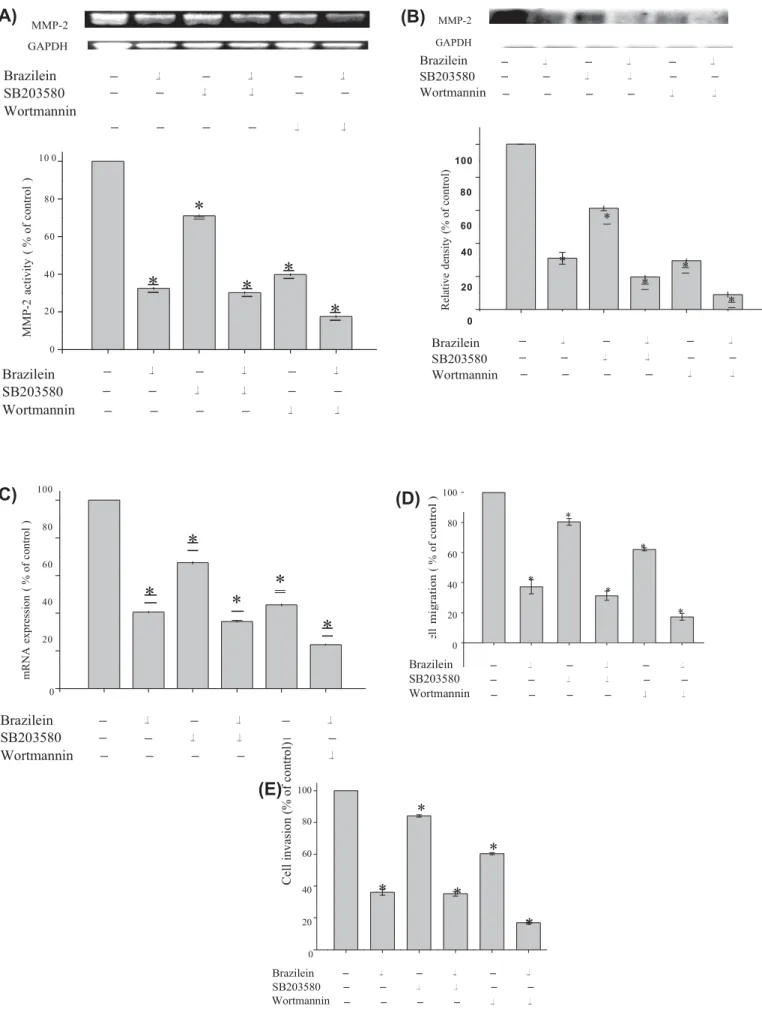
MMP-2 GAPDH(B)
MMP-2 GAPDHBrazilein
SB203580
Wortmannin
Brazilein SB203580 Wortmannin 10 0 100 80 80*
60 60 40*
*
20*
0Brazilein
SB203580
Wortmannin
40*
*
20*
*
0 Brazilein SB203580 Wortmannin(C)
100 80 60 40 20*
*
*
*
*
(D)
100 80 * 60 * 40 * * 20 * 0 0Brazilein
SB203580
Wortmannin
Brazilein SB203580 Wortmannin(E)
100*
80 60*
40*
*
20*
0 Brazilein SB203580 WortmanninFig. 6. Effect of PI3 K inhibitor (wortmannin) or p38 MAPK inhibitor (SB203580) on MMP-2 activity and expression as well as cell migration and cell invasion. Cells were pretreated with wortmannin (10 lM) or SB202580 (10 lM) for 1 h and then incubated in the presence or absence of brazilein (20 lM) for 24 h. Afterward, cells were subjected to analyze for (A) gelatin zymography to analyze the activity of MMP-2, (B) Western blotting and (C) RT-PCR to analyze the protein level and mRNA of MMP-2 as
R el at iv e de ns ity ( % o f co nt ro l) M M P-2 ac tiv ity ( % o f co nt ro l )
*
*
C el l m ig ra ti o n ( % o f co n tr o l ) m R N A e xp re ss io n ( % o f co nt ro l ) C el l i nv as io n (% o f co nt ro l)described in Section 2. Data represented as the mean ± SD of at least three independent experiments (⁄p < 0.05). Effects of inhibitors and brazilein on cell migration (D) and cell invasion (E) as described in Section 2. Data represented as the mean ± SD of three independent experiments. ⁄P < 0.05, compared with that of the untreated control.
3.2. Effect of brazilein on migration and invasion of MDA-MB-231 cells
To investigate the inhibitory effect of brazilein on migration of MDA-MB-231 cells, the wound-healing assay was performed. The confluent monolayer was scraped by a sterile micropipette tip to create a scratch wound. After incubation with brazilein for 24 h, the cells migrated to the denuded zone and the level of the wound closure area was analyzed. The results demonstrated that brazilein suppressed migration of MDA-MB-231 cells to the denuded zone in a dose-dependent manner. Treatment with brazilein (5, 10 and 20
l
M) inhibited by 18.1%, 47.4%, and 63.2% of cell migration, respectively (Fig. 2A). To further examine the effect of brazilein on the invasive ability of MDA-MB-231 cells, a Boyden chamber coated with Matrigel was used. Our data showed that the number of cells invading the lower chamber was significantly reduced by brazilein and a 61% reduction in cell invasion was observed for treatment with 20l
M brazilein (Fig. 2B). These results suggest that brazilein is effective in preventing MDA-MB-231 cell migration and invasion.3.3. Effects of brazilein on activity and expression of MMP-2
To clarify if MMPs are involved in the inhibition of migration and invasion by brazilein, the effects of brazilein on MMP activities were investigated by gelatin zymography. As shown in Fig. 3A, bra-zilein treatment elicited a dose-dependent reduction in MMP-2 activity, whereas MMP-9 activity was not affected. The results of Western blotting and reverse transcription (RT)-PCR showed that brazilein suppressed the expression of MMP-2 protein and mRNA (Fig. 3B and C). To explore the causal relationship between cell migration/invasion and MMP-2 in MDA-MB-231 cells, cell migra-tion and invasion assay were conducted using a potent selective inhibitor of MMP-2, MMP-2I [17]. Fig. 3D shows that MMP-2 inhib-itor blocked MDA-MB-231 cell migration and invasion.
3.4. Inhibition of NF-
j
B by brazileinNF-
j
B has been known to translocate to the nucleus and regu-late the expression of MMP-2 [4,18]. Thus, the effect of brazilein on the translocation of NF-j
B into the nucleus in MDA-MB-231 cells was examined by immunoblotting analysis of the nuclear extracts prepared from the treated cells. The results showed that the pro-tein levels of NF-j
B in the nucleus were decreased by brazilein treatment in a dose-dependent manner (Fig. 4). To characterize the signaling pathway in inhibition of NF-j
B activity in brazilein-treated MDA-MB-231 cells, change of its upstream regulators was determined. As shown in Fig. 4, decrease in protein level of IKK and phosphorylation of IKK and Ij
Ba
, as well as increase in to-tal Ij
Ba
protein expression were observed, suggesting that brazil-ein suppressed NF-j
B pathway activation.3.5. Effect of brazilein on phosphorylation of MAPKs and PI3K/Akt pathways
To assess whether brazilein inhibits phosphorylation of JNK1/2, p38 MAPK, ERK1/2, Akt, and PI3K, phosphorylated MAPK family members (JNK1/2, ERK1/2, and p38 MAPK) and PI3K/Akt in brazil-ein-treated MDA-MB-231 cells were examined. Brazilein sup-pressed phosphorylation of p38 MAPK, PI3K, and Akt in a dose-dependent manner, but the levels of phospho-ERK1/2 and phos-pho-JNK were not affected by brazilein (Fig. 5). Under the used concentration ranging from 5 to 20
l
M), brazilein-inhibited the activation of PI3 K/Akt and p38 MAPK was coincided with the kinetics of cell migration and invasion (Fig. 2).3.6. Inhibitory effect of brazilein on MMP-2 activity and cell migration via PI3K/Akt and p38 MAPK signaling
Induction of p38 MAPK- or PI3K/Akt-signaling pathway has been reported to be involved in MMP-2 expression [19–21]. A selective PI3K inhibitor wortmannin or a p38 MAPK inhibitor SB203580 had been proven to specifically block PI3K and p38 MAPK activities [22]. The effect of wortmannin and SB203580 on the migration assay of MDA-MB-231 cells was examined. Results of gelatin zymography
assay showed that treatment of brazilein, SB203580 (10
l
M) or wortmannin (10l
M) alone reduced MMP-2 activity in culture med- ium of MDA-MB-231 cells by 67.1%, 28.9%, and 60.2%, respectively.Combined treatments could further reduce MMP-2 activation by 69.8% (10
l
M SB203580 + 20l
M brazilein) and 82.4% (10l
M wort-mannin + 20l
M brazilein) (Fig. 6A). Western blot and RT-PCR anal-yses revealed that MMP-2 down-regulation was significantly inhibited by pretreatment with SB203580 or wortmannin. Co-treat-ment of brazilein with SB203580 or wortmannin led to potently in-hibit the MMP-2 expression compared with that of SB203580 or wortmannin (Fig. 6B and C). In addition, the migration assay has re-vealed that treatment with brazilein, SB203580 or wortmannin alone decreased cell migration by 62.7%, 19.6% and 38.8%, respec-tively. Treatment with 10l
M SB203580/20l
M brazilein and 10l
M wortmannin/20l
M brazilein reduced cell migration by 68.6% and 82.7%, respectively (Fig. 6D). Furthermore, the invasion assay had revealed that a sole treatment with SB203580 or wort-mannin led to decreased cell invasion, while the combination treat-ment could further reduce cell invasion, compared with the control (Fig. 6E). Taken together, these data suggest that PI3K/Akt and p38 MAPK pathway activation are involved in brazilein-induced inacti-vation of MMP-2 as well as cell migration and cell invasion of MDA-MB-231 cells. We also investigated the effect of these inhibi-tors [wortmannin (10l
M) and SB203580 (10l
M) ] on the viability of MDA-MB-231 cells, and found that these inhibitors did not influ-ence in vitro proliferative capability of MDA-MB-231 cells, indicat-ing that the anti-migratory and anti-invasive effects of these inhibitors on MDA-MB-231 cells did not arise from interference of cellular proliferative capability.4. Discussion
In recent years, considerable emphasis has been given to iden-tify new anti-cancer agents from natural sources, which could be useful for human beings. Brazilein, a naturally occurring tetracyclic homoisoflavonoid, has a wide range of pharmacologic effects, such as inhibition of cell proliferation, inhibition of cell cycle progres-sion, and induction of apoptosis in various cancer cell lines. Meta-static spread of cancer is responsible for 90% of human cancer-related deaths and thus remains one of the important impediments on cancer curing [23,24]. However, whether brazilein exerts an inhibitory effect on tumor metastasis in breast cancer has not been elucidated previously. When MDA-MB-231 cells were treated with brazilein at non-toxic doses, cell migration and invasion of MDA-MB-231 cells were inhibited (Fig. 2A and B). These results indicated that brazilein-inhibited MDA-MB-231 cell migration and invasion was not due to its cytotoxic effect. In this study, it was found that brazilen-inhibited migration and invasion of MDA-MB-231 cells is closely related to reduction in the activity and expression of MMP-2. Suppression of MMP-2 expression is tightly correlated with inac-tivation of PI3K/Akt, p38 MAPK and NF-
j
B.The enhancement of MMPs expression is involved in tumor invasion, metastasis, and angiogenesis, and the inhibition of MMPs expression should play an important role in cancer therapy be-cause MMPs trigger the degradation of ECM and induce tumor invasion. MMPs are a family of zinc-containing endopeptidases,
and of these, MMP-2 and MMP-9 are highly expressed in aggres-sive breast tumors and are associated with poor clinical outcome [5,25]. To further explore the exact mechanism of brazilein-in-duced inhibition of cell invasion and migration, we performed experiments including gelatin zymography, western blotting and RT-PCR to detect MMP-2 and MMP-9 expression. Our data show that the activity and expression of MMP-2 is notably reduced by brazilein treatment (Fig. 3).
The transcription of MMPs is reported to be regulated by NF-
j
B [26]. NF-j
B is normally sequestered in the cytoplasm in an inactive form associated with its inhibitory protein, inhibitor of kappaBa
(Ij
Ba
). Upon application of an appropriate stimulus, NF-j
B (p65) subunit is released to the nucleus to initiate targeted gene tran-scription as a result of Ij
B kinase (IKK) complex-mediated phos-phorylation and subsequent proteasomal degradation of Ij
Ba
[27,28]. Results from this study found that brazilein treatment led to inhibition of NF-j
B activation, which was accompanied by inhibition of IKK phosphorylation, followed by decrease in Ij
Ba
phosphorylation and prevention of Ij
Ba
proteasomal degradation (Fig. 4). Taken together, these results suggest that the inhibitory ef-fect of brazilein on NF-j
B signaling pathway can be attributed to its inhibition on NF-j
B nuclear translocation and thus suppression of NF-j
B transcriptional activity. These findings were in line with previous results showing that NF-j
B is a pivotal transcription fac-tor in regulating MMP-2 expression in cancer cells [29,30].It is well established that mitogen-activated protein kinase (MAPK) plays important roles in tumor progression and metastasis [7,8]. As shown in Fig. 5, brazilein significantly suppressed the acti-vation of p38 MAPK. In contrast, brazilein did not significantly affect the levels of phospho-ERK1/2 and JNK1/2. p38 MAPK signal trans-duction has been identified to be required for NF-
j
B-dependent gene expression [31–33]. Our data revealed that brazilein treatment inhibited p38 MAPK phosphorylation and caused a concurrent reduction in the level of MMP-2, indicating a possible inhibition mechanism of MMP-2 expression by brazilein. The involvement of p38 MAPK pathway was further supported by the use of p38 MAPK inhibitor (SB203580) in our experimental model. Treatment with SB203580 could inhibit MMP-2 activity and expression as well as cell migration and invasion. Additionally, the PI3K-Akt signaling pathway also played a crucial role in MMP gene regulation, cell sur-vival and tumor cell invasion [15,18]. In our studies, treatment of brazilein inhibited phosphorylation of PI3K and Akt, which was fur-ther supported by experiments using PI3K inhibitor. Moreover, PI3K inhibitor could lead to a reduction in MMP-2 activation, cell migra- tion and cell invasion. Pterostilbene has recently been shown to in- duce MMP-2 gene expression via suppressing multiple signal transduction pathways [34]. Shen et al. demonstrated that acacetin inhibited the invasion and migration of human prostate cancer DU145 cells via inactivation of p38 MAPK signaling pathway [20]. Lee et al. also suggested that PI3K/Akt pathway was required for the inhibitory effects of andrographolide on migration and invasion of A549 cells [35]. The results support the notion that brazilein inhibits invasion and migration of MDA-MB-231 cells could be med- iated through p38 MAPK- and pI3K/Akt-mediated suppression of MMP-2 expression. Additionally, as brazilein is a polyphenol that also contains alpha–beta-unsaturated carbonyl, it is likely to acti- vated nuclear factor E2-related factor 2 (Nrf2) and other pathways [36,37] in addition to inhibit those described here, but further work is needed to explore this possibility.5. Conclusion
In conclusion, the suppressed effect of brazilein on expression and activity of MMP-2 might be correlated with the inactivation of p38 MAPK, PI3K/Akt and NF-
j
B pathway. The suppressive effecton MMP-2 expression contributes to anti-migration and invasion activity of brazilein on MDA-MB-231 cells. These findings reveal a new therapeutic utility of brazilein on anti-metastatic therapy.
Conflict of interest
None.
Acknowledgments
This work was supported by grants from the National Science Council of Taiwan (NSC97-2320-B-037-008-MY3) and NSYSU-KMU Joint Research Project (NSYSUNSYSU-KMU-102-004).
References
[1] A. Jemal, R. Siegel, E. Ward, Y. Hao, J. Xu, M.J. Thun, Cancer statistics, CA Cancer J.
Clin. 59 (2009) (2009) 225–249.
[2] H. Nagase, J.F. Woessner Jr., Matrix metalloproteinases, J. Biol. Chem. 274 (1999) 21491–21494.
[3] E.I. Deryugina, J.P. Quigley, Matrix metalloproteinases and tumor metastasis, Cancer Metastasis Rev. 25 (2006) 9–3 4 .
[4] S. Chakraborti, M. Mandal, S. Das, A. Mandal, T. Chakraborti, Regulation of matrix metalloproteinases: an overview, Mol. Cell Biochem. 253 (2003) 269– 285.
[5] G.S. Delassus, H. Cho, J. Park, G.L. Eliceiri, New pathway links from cancer-progression determinants to gene expression of matrix metalloproteinases in breast cancer cells, J. Cell Physiol. 217 (2008) 739–744.
[6] G.L. Johnson, R. Lapadat, Mitogen-activated protein kinase pathways mediated by
ERK, JNK, and p38 protein kinases, Science 298 (2002) 1911–1912. [7] K.B. Reddy, S.M. Nabha, N. Atanaskova, Role of MAP kinase in tumor
progression and invasion, Cancer Metastasis Rev. 22 (2003) 395–403. [8] M. Qiao, S. Sheng, A.B. Pardee, Metastasis and AKT activation, Cell Cycle 7
(2008) 2991–2996.
[9] Ye, M. D. Hu, L. Tu, X. Zhou, F. Lu, B. Wen, W. Wu, Y. Lin, Z. Zhou, J. Qu, Involvement of PI3K/Akt signaling pathway in hepatocyte growth factor-induced migration of uveal melanoma cells, Invest. Ophthalmol. Vis. Sci. 49 (2008) 497–50 4 .
[10] P.N. Chen, Y.S. Hsieh, H.L. Chiou, S.C. Chu, Silibinin inhibits cell invasion through inactivation of both PI3K-Akt and MAPK signaling pathways, Chem. Biol. Interact. 156 (2005) 141–15 0 .
[11] I.M. Lockhart, 3-Chromanols and 4-chromanols, in: G.P. Ellis (Ed.), Chemistry of
Heterocyclic Compounds Chromenes Chromanones and Chromones, John Wiley & Sons, New York, 1977, pp. 141–192.
[12] C.T. Yen, K. Nakagawa-Goto, T.L. Hwang, P.C. Wu, S.L. Morris-Natschke, W.C. Lai, K.F. Bastow, F.R. Chang, Y.C. Wu, K.H. Lee, Antitumor agents 271: total synthesis and evaluation of brazilein and analogs as anti-inflammatory and cytotoxic agents, Bioorg. Med. Chem. Lett. 20 (2010) 1037–103 9 .
[13] X. Zhong, B. Wu, Y.J. Pan, S. Zheng, Brazilein inhibits survivin protein and mRNA expression and induces apoptosis in hepatocellular carcinoma HepG2 cells, Neoplasma 56 (2009) 387–392.
[14] V. Betapudi, L.S. Licate, T.T. Egelhoff, Distinct roles of nonmuscle myosin II isoforms in the regulation of MDA-MB-231 breast cancer cell spreading and migration, Cancer Res. 66 (2006) 4725–4733.
[15] D. Kim, S. Kim, H. Koh, S.O. Yoon, A.S. Chung, K.S. Cho, J. Chung, Akt/PKB promotes cancer cell invasion via increased motility and metalloproteinase production, FASEB J. 15 (2001) 1953–1962.
[16] K.L. Lin, J.C. Su, C.M. Chien, C.H. Tseng, Y.L. Chen, L.S. Chang, S.R. Lin, Naphtho [1,2-b]furan-4,5-dione induces apoptosis and S-phase arrest of MDA-MB-231 cells through JNK and ERK signaling activation, Toxicol. Vitro 24 (2010) 61–7 0 . [17] B. Zhang, J. Zhang, H.Z. Huang, W.L. Chen, Q. Tao, D.L. Zeng, L.T. Zhang, J.H. Xu,
Inhibition of ameloblastoma invasion in vitro and in vivo by inhibitor of metalloproteinase-2 activity, J. Oral Pathol. Med. 38 (2009) 731–736. [18] J. Westermarck, V.M. Kahari, Regulation of matrix metalloproteinase
expression in tumor invasion, FASEB J. 13 (1999) 781–792.
[19] B. Kumar, S. Koul, J. Petersen, L. Khandrika, J.S. Hwa, R.B. Meacham, S. Wilson, H.K. Koul, P38 mitogen-activated protein kinase-driven MAPKAPK2 regulates invasion of bladder cancer by modulation of MMP-2 and MMP-9 activity, Cancer Res. 70 (2010) 832–841.
[20] K.H. Shen, S.H. Hung, L.T. Yin, C.S. Huang, C.H. Chao, C.L. Liu, Y.W. Shih, Acacetin, a flavonoid, inhibits the invasion and migration of human prostate cancer DU145 cells via inactivation of the p38 MAPK signaling pathway, Mol. Cell. Biochem. 333 (2010) 279–29 1 .
[21] Y.W. Shih, P.S. Chen, C.H. Wu, Y.F. Jeng, C.J. Wang, Alpha-chaconine-reduced metastasis involves a PI3K/Akt signaling pathway with downregulation of NF-kappaB in human lung adenocarcinoma A549 cells, J. Agric. Food Chem. 55 (2007) 11035–1104 3 .
[22] K.D. Shin, M.Y. Lee, D.S. Shin, S. Lee, K.H. Son, S. Koh, Y.K. Paik, B.M. Kwon, D.C. Han, Blocking tumor cell migration and invasion with biphenyl isoxazole derivative KRIBB3, a synthetic molecule that inhibits Hsp27 phosphorylation, J.
[23] D. Hanahan, R.A. Weinberg, The hallmarks of cancer, Cell 100 (2000) 57–7 0 . [24] P.S. Steeg, Tumor metastasis: mechanistic insights and clinical challenges, Nat.
Med. 12 (2006) 895–904.
[25] M.J. Duffy, T.M. Maguire, A. Hill, E. McDermott, N. O’Higgins, Metalloproteinases: role in breast carcinogenesis, invasion and metastasis, Breast Cancer Res. 2 (2000) 252–25 7 .
[26] Sliva,D. Signaling pathways responsible for cancer cell invasion as targets for cancer therapy, Curr. Cancer Drug Targets 4 (2004) 327–336.
[27] S. Ghosh, D. Baltimore, Activation in vitro of NF-kappa B by phosphorylation of its
inhibitor I kappa B, Nature 344 (1990) 678–682.
[28] S. Ghosh, M.S. Hayden, New regulators of NF-kappa B in inflammation, Nat. Rev. Immunol. 8 (2008) 837–848.
[29] Y.T. Yang, C.J. Weng, C.T. Ho, G.C. Yen, Resveratrol analog-3,5,40
-trimethoxy-trans-stilbene inhibits invasion of human lung adenocarcinoma cells by suppressing the MAPK pathway and decreasing matrix metalloproteinase-2 expression, Mol. Nutr. Food Res. 53 (2009) 407–416.
[30] H.H. Lin, J.H. Chen, F.P. Chou, C.J. Wang, Protocatechuic acid inhibits cancer cell metastasis involving the down-regulation of Ras/Akt/NF-kappaB pathway and MMP-2 production by targeting RhoB activation, Br. J. Pharmacol. 162 (2011) 237–254.
[31] A.B. Carter, K.L. Knudtson, M.M. Monick, G.W. Hunninghake, The p38 mitogen-activated protein kinase is required for NF-kappaB-dependent gene expression. The role of TATA-binding protein (TBP), J. Biol. Chem. 274 (1999) 30858–30863.
[32] E.S. Kim, M.S. Kim, A. Moon, TGF-beta-induced upregulation of MMP-2 and MMP-9 depends on p38 MAPK, but not ERK signaling in MCF10A human breast epithelial cells, Int. J. Oncol. 25 (2004) 1375–1382.
[33] M.L. Lin, Y.C. Lu, J.G. Chung, S.G. Wang, H.T. Lin, S.E. Kang, C.H. Tang, J.L. Ko, S.S. Chen, Down-regulation of MMP-2 through the p38 MAPK-NF-kappaB-dependent pathway by aloe-emodin leads to inhibition of nasopharyngeal carcinoma cell invasion, Mol. Carcinog. 49 (2010) 783–79 7 .
[34] M.H. Pan, Y.S. Chiou, W.J. Chen, J.M. Wang, V. Badmaev, C.T. Ho, Pterostilbene inhibited tumor invasion via suppressing multiple signal transduction pathways in human hepatocellular carcinoma cells, Carcinogenesis 30 (2009) 1234–1242.
[35] Y.C. Lee, H.H. Lin, C.H. Hsu, C.J. Wang, T.A. Chiang, J.H. Chen, Inhibitory effects of
andrographolide on migration and invasion in human non-small cell lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway, Eur. J. Pharmacol. 632 (2010) 23–32.
[36] Y. Nakamura, C. Yoshida, A. Murakami, H. Ohigashi, T. Osawa, K. Uchida, Zerumbone, atropical ginger sesquiterpene, activates phase II drug metabolizing enzymes, FEBS Lett. 572 (2004) 245–250.
[37] K. Umemura, T. Itoh, N. Hamada, Y. Fujita, Y. Akao, Y. Nozawa, N. Matsuura, M. Iinuma, M. Ito, Preconditioning by sesquiterpene lactone enhances H2O2-induced Nrf2/ARE activation, Biochem. Biophys. Res. Commun. 368 (2008) 948–954.