Human ATP-binding Cassette Transporter ABCB1 Confers Resistance to Volasertib (BI 6727), a Selective Inhibitor of Polo-Like Kinase 1
Chung-Pu Wu †,‡,#,*, Chia-Hung Hsieh ※,&, Sung-Han Hsiao ‡, Shi-Yu Luo ‡, Ching-Ya Su ‡, Yan-Qing Li †, Yang-Hui Huang #, Chiun-Wei Huang ∥and Sheng-Chieh Hsu ‡,#,⊥,* *
Authors' Affiliations:
† Department of Physiology and Pharmacology, ‡ Graduate Institute of Biomedical Sciences,# Molecular Medicine Research Center, and ⊥ Department of Biomedical Sciences,College of Medicine, Chang Gung University, Tao-Yuan 333, Taiwan. ※ Graduate Institute of Basic Medical Science and & Department of Medical Research, China Medical University Hospital, Taichung, Taiwan. ∥Center for Advanced
Molecular Imaging and Translation, Chang Gung Memorial Hospital, Tao-Yuan, Taiwan.
* Corresponding author at: 259 Wen-Hwa 1st Road, Kwei-Shan, Tao-Yuan 333, Taiwan. Phone: 886-3-2118800, ext. 3754. Fax: 886-3-2118700.
** Corresponding author at: 259 Wen-Hwa 1st Road, Kwei-Shan, Tao-Yuan 333, Taiwan. Phone: 886-3-2118800, ext. 3690. Fax: 886-3-2118700.
E-mail address: [email protected] (C.-P. Wu), [email protected]
(S.-C. Hsu).
Running Title: ABCB1 confers resistance to volasertib
Abbreviations: MDR, multidrug resistance; ABC, ATP-binding cassette; Plk-1,
Polo-like kinase 1; MTT, 3-(4,5-dimethylthiazol-yl)-2,5-diphenyllapatinibrazolium bromide.
ABSTRACT
The overexpression of the serine/threonine specific polo-like kinase 1 (Plk1) is associated with poor prognosis in many types of cancer. Consequently, Plk1 has emerged as a valid therapeutic target for anticancer drug design. Volasertib is a potent inhibitor of Plk1 that inhibits the proliferation of multiple human cancer cell lines by promoting cell cycle arrest at nanomolar concentrations. However, the risk of
developing drug resistance, which is often associated with the overexpression of the ATP-binding cassette (ABC) transporter ABCB1 (P-glycoprotein), can present a therapeutic challenge for volasertib and many other therapeutic drugs. Although volasertib is highly effective against the proliferation of numerous cancer cell lines, we found that the overexpression of ABCB1 in cancer cells leads to cellular resistance to volasertib and reduces the level of volasertib-stimulated G2/M cell cycle arrest and subsequent onset of apoptosis. Furthermore, we demonstrate that volasertib
competitively inhibits the function of ABCB1 and stimulates the basal ATPase activity of ABCB1 in a concentration-dependent manner, which is consistent with substrate transport by ABCB1. More importantly, we discovered that the co-administration of an inhibitor or drug substrate of ABCB1 restored the anticancer activity of volasertib in ABCB1-overexpressing cancer cells. In conclusion, the results of our study reveal that ABCB1 negatively affects the efficacy of volasertib and supports its combination with a modulator of ABCB1 to improve clinical responses.
INTRODUCTION
Polo-like kinase 1 (Plk1) is a serine/threonine protein kinase that plays a crucial role in various aspects that regulate mitosis,1-3 such as mitotic entry,4 mitotic exit,5
centrosome maturation6 and bipolar spindle formation.7 Plk1 is reportedly
overexpressed in numerous types of cancer8-11 and is associated with poor prognosis
and survival. Studies demonstrate that Plk1 inhibition induces significant cell cycle arrest, apoptosis and eventual cancer cell death, prompting the development of potent and selective Plk1 inhibitors as a new therapeutic option for cancer.16-18 BI 2536 is the
first small-molecule inhibitor of Plk1 developed. This compound inhibits cancer cell proliferation by promoting cell cycle arrest and apoptosis in numerous cancer types. Unfortunately, phase II trials of BI 2536 revealed unfavorable pharmacological properties that limit BI 2536 from further clinical development.20-23
Volasertib (BI 6727) is a Plk1 inhibitor that has been improved by modifying the structure of BI 2536 to enhance Plk1 inhibition. This agent limits cancer cell
proliferation by inducing significant mitotic arrest and apoptosis in cancer cells. Preclinical studies demonstrated that volasertib is effective against neuroblastoma tumor-initiating cells,25 improves local tumor control when applied simultaneously to
fractionated irradiation,26 and exerts a synergistic antitumor effect with histone
deacetylase inhibitors in combination therapy.27 Volasertib exhibits a high volume of
distribution, long terminal half-life and good antitumor activity in multiple cancer models, with half maximal effective concentration (EC50) values ranging from 10 to 40 nM. Volasertib has also been demonstrated to possess encouraging anticancer activity in multiple xenograft and animal models.24 Recent preclinical data suggest
that volasertib is an effective treatment for bladder cancer,28 glioblastoma and acute
myeloid leukemia (AML)31 either as a single agent or in combination with other
established AML drugs.32 More importantly, volasertib exhibited promising antitumor
activity and pharmacological profiles in its Phase I and II studies, especially in patients with AML33-35 or advanced solid tumors.
One of the most common mechanisms for the development of multidrug resistance (MDR) in cancer cells and a major obstacle in chemotherapy is the overexpression of ATbinding cassette (ABC) drug transporter ABCB1 (Pgp; P-glycoprotein). Human ABCB1 is the first identified member of the mammalian ABC protein family that can recognize a diverse range of therapeutic agents and utilize ATP hydrolysis to actively transport drug substrates out of cancer cells, thus leading to MDR, cancer relapse and death. Moreover, ABCB1 is highly expressed at several sites, such as the blood-brain barrier (BBB), liver and intestinal walls; thus, this transporter significantly contributes to the oral bioavailability and distribution of therapeutic drugs. In addition to conventional anticancer agents, ABCB1 interacts with many newly developed small-molecule kinase inhibitors, reducing the bioavailability and overall effectiveness of these targeted anticancer agents.42-48
Therefore, studying the pharmacological and biochemical interactions between ABCB1 and these small-molecule kinase inhibitors is important.
Previously, we discovered that the effectiveness of BI 2536, the first-generation selective inhibitor of Plk1, is significantly reduced in cells overexpressing human ABCB1.45 In the present study, we investigated the interactions between the
second-generation Plk1 inhibitor volasertib and ABCB1. We reveal that ABCB1
Furthermore, although volasertib competitively inhibits ABCB1-mediated drug transport, it is not sufficiently potent to reverse ABCB1-mediated drug resistance at non-toxic concentrations. More importantly, we demonstrate that the reduced volasertib-induced G2/M cell cycle arrest, apoptosis and cytotoxicity in ABCB1-overexpressing cells is restored by transiently inhibiting the function of ABCB1, providing a rationale for development of new combination therapies based on volasertib and ABCB1 modulators.
EXPERIMENTAL SECTION
Chemicals. Dulbecco's Modified Eagle's medium (DMEM), RPMI medium, fetal calf serum (FCS), phosphate-buffered saline (PBS), trypsin-EDTA, penicillin, and
streptomycin were purchased from Gibco under Life Technologies (Grand Island, NY, USA). MTT dye, Cell Counting Kit-8 (CCK-8) and all other chemicals were purchased from Sigma (St. Louis, MO, USA) unless stated otherwise. Tariquidar was a generous gift from Dr. Susan Bates (National Cancer Institute, NIH, Bethesda, MD, USA). Volasertib (99% purity by HPLC, Chiral HPLC) was purchased from
ChemieTek (Indianapolis, IN, USA).
Cell Lines and Culture Conditions. KB-3-1, KB-C-1, KB-V-1, OVCAR-8, NCI-ADR-RES, NIH3T3, NIH3T3-G185, pcDNA3.1-HEK293 and MDR19-HEK293 cells were cultured in DMEM supplemented with 10% FCS, 2 mM L-glutamine and 100 units of penicillin/streptomycin/mL. KB-V-1 cells were maintained in media containing 1 µg/µL vinblastine.49 NIH3T3-G185 cells were maintained in the
presence of 60 ng/mL colchicine.50 pcDNA3.1-HEK293 and MDR19-HEK293 cells
from Dr. Suresh V. Ambudkar (National Cancer Institute, NIH, Bethesda, MD, USA) and were maintained at 37 °C in 5% CO2 humidified air.
Cytotoxicity Assay. MTT and CCK-8 assays were used to determine the general sensitivities of cells to the tested chemicals, as described previously.46 For the reversal
of cytotoxicity assays, a nontoxic concentration of ABCB1 modulator was added to the cytotoxicity assay, and the extent of reversal was then calculated based on the relative resistance (R.R.) values.
Cell Cycle Analysis and Apoptosis Assay. Cell-cycle experiments were performed using a standard propidium iodide (PI) staining method, and the cells were analyzed using a FACSort flow cytometer equipped with the CellQuest software. Briefly, cells were treated with the indicated regimens for 24 hrs before being harvested in PBS and fixed in ethanol overnight. The cells were washed once with PBS then treated with 0.5% TritonX-100 and 0.05% RNase in PBS at 37 °C for 1 hr. The cells were washed again, and they were then incubated with PI (50 μg/mL) at 4 °C for 20 min prior to analysis. To determine the percentage of apoptotic cells, the cells were treated with the indicated regimens for 48 hrs before harvest, centrifuged and resuspended in FACS buffer containing 1.25 µg/mL annexin V–FITC (PharMingen) and 0.1 mg/mL PI. The cells were incubated in this buffer for 15 min at room temperature. The labeled cells (10,000 per sample) were then analyzed by FACScan (BD Biosciences) using the CellQuest software. Cells in the lower right dot-plot quadrant (PS-positive and PI-negative cells) were considered apoptotic and have intact plasma membranes, whereas cells in the upper right dot-plot quadrant (PS-positive and PI-positive cells) have leaky membranes and were considered either necrotic or late apoptotic.
ATPase Assay of ABCB1. The vanadate (Vi)-sensitive ABCB1-specific ATPase activities were recorded using the Pgp-Glo assay system (Promega, WI, USA)
according to the manufacturer's instructions.47 The drug-stimulated ATPase activity of
ABCB1 was determined based on endpoint Pi assay as described previously.51
Fluorescent Drug Accumulation Assay. The ABCB1-mediated efflux assay was performed using a FACSort flow cytometer equipped with CellQuest software (Becton-Dickinson) as described previously.52 The effect of volasertib and tariquidar
on the ABCB1-mediated calcein-AM efflux was measured and analyzed as described previously.52 The mean fluorescence intensity was calculated with the histogram stat
program in the CellQuest software.
Immunoblotting. Antibodies C219 (1:1000) and α-tubulin (1:2000) were used to detect ABCB1 and tubulin, respectively. The latter served as a positive control for Western blotting. The horseradish peroxidase-conjugated goat anti-mouse IgG antibody (1:10,000) was used as the secondary antibody. The signals were detected as described previously.53
Statistical Analysis. Data are presented as the mean ± standard error of the mean (SEM), whereas the half maximal inhibitory concentration (IC50) values are the calculated mean ± standard deviation (SD) from at least three independent experiments. Differences between mean values were analyzed using two-sided Student’s t-tests and were considered significant at P < 0.05.
RESULTS
The Cytotoxicity of Volasertib Is Significantly Reduced in ABCB1
Overexpressing Cells. First, we examined the effect of ABCB1 on the efficacy of volasertib by comparing the cytotoxicity of volasertib in drug-sensitive and ABCB1-overexpressing drug-resistant cell lines. The results of this experiment are
summarized in Table 1, and the resistance factor (RF) value represents the degree of resistance to volasertib caused by the presence of ABCB1, which was calculated by dividing the IC50 value of the ABCB1-overexpressing sub-line by the IC50 value of the respective parental line. The ABCB1-overexpressing human epidermal KB-C-1 and KB-V-1 cells were significantly less sensitive to volasertib compared with parental KB-3-1 cells (RF values of 4, P < 0.001 and 160, P < 0.001, respectively) (Fig. 1A, Table 1). Similarly, the ABCB1-overexpressing human ovary NCI-ADR-RES cells were significantly more resistant to volasertib than the parental OVCAR-8 cells (RF value of 31, P < 0.01). We further confirmed our findings by examining the
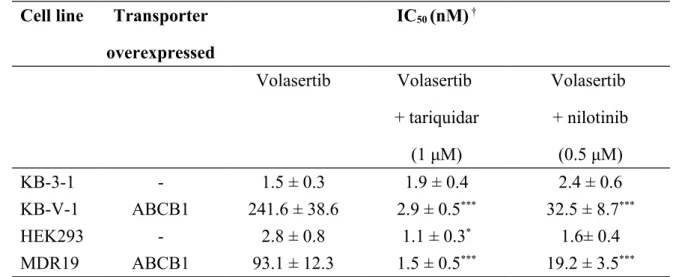
cytotoxicity of volasertib in cells transfected with human ABCB1 protein. We found that ABCB1-transfected human embryonic kidney MDR19-HEK293 cells (Fig. 1B) and ABCB1-transfected mouse fibroblast NIH3T3-G185 cells were both significantly less sensitive to volasertib compared with their respective parent cells (Table 1).
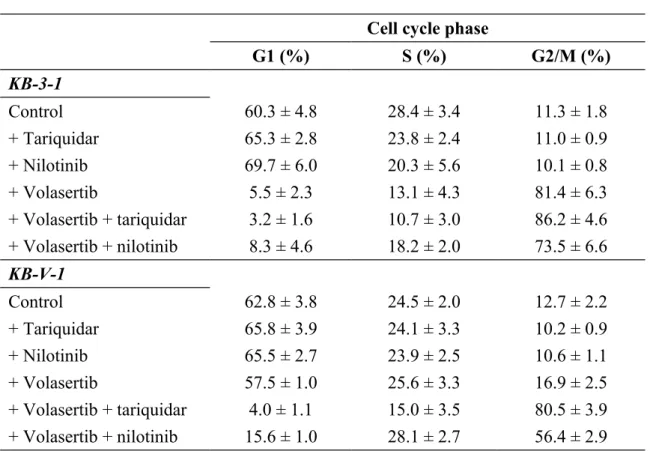
The G2/M Cell Cycle Arrest, Apoptosis and Cytotoxicity Induced by Volasertib in ABCB1-Overexpressing Cells Can Be Restored by Inhibiting the Function of ABCB1. Next, we determined the impact of ABCB1 function on volasertib-induced G2/M arrest and apoptosis in KB-V-1 cells given that G2/M cell cycle arrest is a hallmark of Plk1 inhibitors and that KB-V-1 cells are highly resistant to volasertib
(Table 1). We observed substantial G2/M cell cycle arrest induced by volasertib in KB-3-1 cells (Fig. 2B, upper left panel) but not in ABCB1-overexpressing KB-V-1 cells (Fig. 2B, lower left panel). The exposure of cells to 10 nM volasertib for 24 hrs resulted in a considerable increase in G2/M phase arrest in KB-3-1 cells from a basal level of 11% to approximately 81% after treatment. In contrast, the increase in G2/M phase arrest in KB-V-1 cells was small from a basal level of 12% to only 17% after treatment. Although the ABCB1 inhibitor tariquidar (1 µM) and ABCB1 drug
substrate nilotinib (5 μM) each did not affect cell cycle arrest in KB cancer cells when applied individually (Fig. 2A), these agents substantially increased G2/M cell cycle arrest in KB-V-1 cells when the function of ABCB1 was blocked (Fig. 2B, lower
panels) from a basal level of 12% to approximately 81% and 56%, respectively. The
overall effects of volasertib, tariquidar and nilotinib on the phases of the cell cycle in KB cells are summarized in Table 2.
We next examined the level of apoptosis induced by volasertib in KB cancer cell lines in the presence or absence of tariquidar and nilotinib. In the absence of
volasertib, the basal level of apoptosis was approximately 13% in both cell lines (Fig. 3A). When treated with volasertib (20 nM), the percentage of apoptotic cells
significantly increased in KB-3-1 cells, from 13 to 78% (Fig. 3B, upper left panel). This effect was considerably less pronounced in KB-V-1 cells; the percentage of apoptotic cells increased from 13 to 21% (Fig. 3B, lower left panel). However, when the function of ABCB1 in ABCB1-overexpressing KB-V-1 cells was inhibited by tariquidar (1 μM) (Fig. 3B, lower middle panel) or nilotinib (5 μM) (Fig. 3B, lower
right panel), the level of apoptosis induced by volasertib dramatically increased to
KB-V-1 to volasertib can be significantly restored by tariquidar and nilotinib in the same manner, from an RF value of 160 to values of 2 and 14, respectively (Table 3). Our findings were confirmed with ABCB1-transfected MDR19-HEK293 cells as both tariquidar and nilotinib were able to restore volasertib sensitivity from an RF value of 34 to values of 1 and 11, respectively (Table 3).
Volasertib Stimulates ABCB1 ATPase Activity. Because ABCB1 substrates are known to stimulate Vi-sensitive ABCB1 ATPase activity,51 we next examined the
effect of volasertib on ABCB1-mediated ATP hydrolysis, as described in the
Experimental Section. Volasertib stimulated Vi-sensitive ABCB1 ATPase activity in a
concentration-dependent manner with a maximum stimulation of approximately 3-fold and a concentration of approximately 100 nM required for 50% simulation (Fig. 4). These results demonstrate that volasertib stimulates ABCB1 ATP hydrolysis in the same manner as other well-established substrates.
Volasertib Inhibits the Transport Function of ABCB1 But Does Not Reverse ABCB1-Mediated Drug Resistance in Cancer Cells at Non-Toxic Concentrations. To determine whether volasertib interacts with ABCB1, we examined the mechanism by which volasertib directly affects the function of ABCB1 by evaluating the ability of volasertib to inhibit the ABCB1-mediated efflux of calcein-AM, an established fluorescent substrate of ABCB1,46 in short-term drug accumulation assays as
described in the Experimental Section. The calcein-AM efflux experiments were performed in the absence (solid lines) or presence of 10 μM volasertib (shaded, solid
lines) or 3 μM tariquidar (dotted lines) in KB-3-1 and KB-V-1 cancer cells as well as
in HEK293 and MDR19-HEK293 cells. Although volasertib did not significantly affect parental cells (Fig. 5A and B, left panels), it completely inhibited
ABCB1-mediated calcein-AM efflux in ABCB1-ovexpressing KB-V-1 (Fig. 5A, right panel) and MDR-19-HEK293 cells (Fig. 5B, right panel). Moreover, volasertib inhibited the function of ABCB1 in KB-V-1 cells in a concentration-dependent manner with a calculated IC50 value of approximately 4 μM (Fig. 5C).
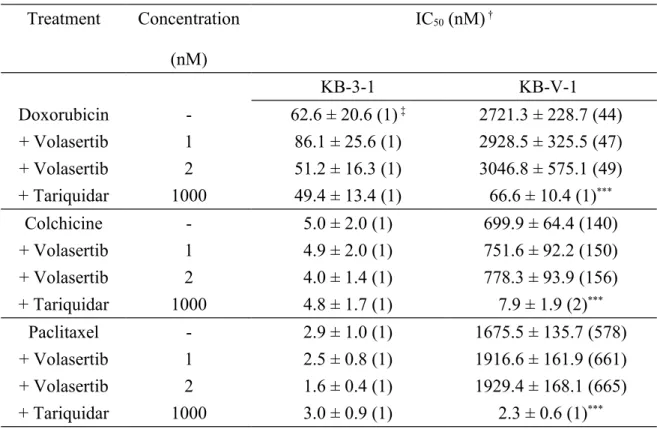
Next, we evaluated the ability of volasertib to restore drug sensitivity in human KB-V-1 epidermal cancer cells. Some drug substrates of ABCB1 reverse drug
resistance in cells overexpressing ABCB1 by competing with the transport of another drug substrate. The maximum concentrations of volasertib (1 nM and 2 nM) were selected based on the cytotoxicity curve (Fig. 1A), and the ability of volasertib to reverse ABCB1-mediated drug resistance in KB-V-1 cells is summarized in Table 4. Tariquidar (1 μM) was used as a positive control to reverse ABCB1-mediated drug resistance. The R.R. value was calculated by dividing the IC50 value of ABCB1-overexpressing KB-V-1 cells by the IC50 value of the parental KB-3-1 cells in the presence of a test drug. Our results indicated that volasertib did not significantly affect ABCB1-mediated resistance to doxorubicin, colchicine or paclitaxel at the tested concentrations.
Volasertib Did Not Significantly Affect ABCB1 Protein Expression. Given that ABCB1 expression depends on the cell cycle57 and numerous ABCB1-interacting
compounds up-regulate or down-regulate ABCB1,58-60 we examined the ability of
volasertib to regulate the expression of ABCB1 protein in human epidermal KB cells. The cells were treated with increasing concentrations of volasertib for 72 hrs and then harvested and processed for the immunodetection of ABCB1 as detailed in the
Experimental Section. Volasertib did not significantly affect ABCB1 protein
DISCUSSION
The development of MDR due to the overexpression of ABCB1 has been a major problem in medical oncology for decades.41 Furthermore, numerous studies
indicate that the overexpression of ABCB1 reduces the bioavailability and efficacy of various tyrosine kinase inhibitors, including some recently developed Plk1 inhibitors. Given that Plk1 is overexpressed in numerous tumor types and its inhibition leads to the induction of G2/M cell cycle arrest, the subsequent onset of apoptosis and eventual cell death has sparked growing interest in developing new agents that selectively inhibit the activity of Plk1. Volasertib is a second-generation Plk1
inhibitor that specifically blocks Plk1 activity and inhibits the proliferation of cancer cells at nanomolar concentrations. Moreover, the favorable pharmacological and safety profile of volasertib identified in clinical trials make it a true candidate for cancer drug development. Because the overexpression of human ABCB1 in cancer cells leads to significant resistance to BI 2536, the first identified selective inhibitor of Plk1,45 we examined the potential impact of ABCB1 overexpression on the
effectiveness of volasertib in human cancer cells.
We herein demonstrate that volasertib is highly cytotoxic to cancer cells, with IC50 values in the nanomolar range, which is consistent with previous reports. However, we found that the overexpression of ABCB1 or transfection with human ABCB1 confers significant resistance to volasertib in cancer cells (Fig. 1). ABCB1 expression has been extensively characterized in the cell lines used in this study, and ABCB1 is only overexpressed in MDR cell lines (Table 1). Notably, the KB-C-1 and KB-V-1 cell lines are both MDR sub-lines of the drug-sensitive human KB-3-1 cell
line and exhibit increased ABCB1 protein expression. Because KB-C-1 cells were more sensitive to volasertib than KB-V-1 cells, the cellular sensitivity of human epidermal cancer cells to volasertib likely inversely correlates with the expression level of ABCB1 (Table 1). Furthermore, we revealed that the volasertib resistance in ABCB1-overexpressing cells is attributed to reduced G2/M cell cycle arrest and subsequent apoptosis induced by volasertib. The induction of G2/M arrest is a hallmark of Plk1 inhibitors. Our data demonstrate that the G2/M cell cycle arrest and apoptosis induced by volasertib were significantly reduced in ABCB1-overexpressing KB-V-1 cells compared with ABCB1-negative KB-3-1 cells. More importantly, volasertib sensitivity was restored to ABCB1-overexpressing cells upon treatment with a competitive inhibitor or a drug substrate of ABCB1 given that ABCB1-mediated drug resistance in cancer cells is often reversed by inhibiting ABCB1 function. We found that inhibiting the function of ABCB1 with tariquidar or nilotinib completely reversed volasertib-induced G2/M arrest and subsequent apoptosis in drug-resistant KB-V-1 cells without affecting the state of the cell cycle or apoptosis in drug-sensitive KB-3-1 cells (Fig. 2 and 3). Notably, combining volasertib with either tariquidar or nilotinib appears to produce an even more pronounced increase in late apoptosis in KB-V-1 cells compared with KB-3-1 cells (Fig. 3B). Moreover, in accordance with cell cycle and apoptosis data, ABCB1-mediated volasertib resistance in KB-V-1 cancer cells and ABCB1-transfected MDR19-HEK293 cells was
significantly reversed by a non-toxic concentration of tariquidar or nilotinib (Table 3).
The combination of paclitaxel with volasertib has recently been shown to affect ABCB1-overexpressing cell lines in a synergistic manner.63 Therefore, we
ABCB1 as well as the ability of this drug to restore drug sensitivity in ABCB1-overexpressing KB-V-1 cells. The stimulation of ABCB1 ATPase activity is coupled with ABCB1-mediated substrate transport, whereas the inhibition of ATPase activity is associated with the presence of an inhibitor or substrate with a significantly reduced transport rate. Therefore, we assessed the effect of volasertib on vanadate-sensitive ABCB1 ATPase activity. Volasertib rapidly stimulated ABCB1 basal ATPase activity in a concentration-dependent manner, supporting the notion that volasertib is a substrate of ABCB1 (Fig. 4). In contrast, To et al. reported that volasertib inhibited the ABCB1 basal ATPase activity in a concentration-dependent manner. Given that the concentrations tested by To et al. (20 μM to 10 mM) significantly exceeded our tested concentrations and pharmacologically relevant concentrations of volasertib,63
the discrepancy between these studies is not easily reconciled. Next, we examined the effect of volasertib on ABCB1-mediated transport and drug resistance. A maximum concentration of 2 nM volasertib was used to reverse ABCB1-mediated resistance to doxorubicin, colchicine and paclitaxel in human KB cells. Although low
concentrations of volasertib competitively inhibit ABCB1-mediated transport (Fig. 5), these concentrations were insufficient to reverse ABCB1-mediated drug resistance (Table 4) or alter the expression of ABCB1 (Fig. 6) in human KB cancer cells, as reported for other ABCB1-interacting drugs. We suspect that the lack of effect of volasertib on ABCB1-mediated drug resistance is due to the highly cytotoxic nature of volasertib and the high level of resistance conferred by ABCB1. These findings do not warrant further testing of volasertib as an MDR-reversing agent.
In summary, the function of ABCB1 significantly reduced the efficacy of
cycle arrest, subsequent apoptosis and cell death. ABCB1 is an important factor in drug disposition and in MDR. Our data indicate that volasertib treatment will likely be less effective against cancer cells that express high levels of ABCB1, and the effect of ABCB1 on the oral pharmacokinetics or tissue distribution of volasertib in vivo remains to be determined. Moreover, although the transient inhibition of ABCB1 appeared to reverse mediated resistance to volasertib in
ABCB1-overexpressing cancer cells, this strategy has demonstrated limited success in clinical trials. Therefore, in addition to combining volasertib with a competitive inhibitor or drug substrate of ABCB1, other novel treatment strategies merit further investigation.
ACKNOWLEDGMENTS.
The authors thank Drs. Susan Bates and Suresh V. Ambudkar (National Cancer Institute, NIH) for generously providing tariquidar and various cell lines. This work was supported by funds from the Chang Gung Medical Research Program
(CMRPD1D0152), the Ministry of Science and Technology (MOST-103-2320-B-182-011) and a grant to Chang Gung University from the Ministry of Education of Taiwan (EMRPD1E1571).
REFERENCES
(1) Petronczki, M.; Lenart, P.; Peters, J. M. Polo on the Rise-from Mitotic Entry to Cytokinesis with Plk1. Dev. Cell. 2008, 14 (5), 646-659.
(2) van Vugt, M. A.; Medema, R. H. Getting in and out of mitosis with Polo-like kinase-1. Oncogene 2005, 24 (17), 2844-2859.
(3) Jackson, J. R.; Patrick, D. R.; Dar, M. M.; Huang, P. S. Targeted anti-mitotic therapies: can we improve on tubulin agents? Nat. Rev. Cancer 2007, 7 (2), 107-117.
(4) Toyoshima-Morimoto, F.; Taniguchi, E.; Shinya, N.; Iwamatsu, A.; Nishida, E. Polo-like kinase 1 phosphorylates cyclin B1 and targets it to the nucleus during prophase. Nature 2001, 410 (6825), 215-220.
(5) Eckerdt, F.; Strebhardt, K. Polo-like kinase 1: target and regulator of anaphase-promoting complex/cyclosome-dependent proteolysis. Cancer Res. 2006, 66 (14), 6895-6898.
(6) Lane, H. A.; Nigg, E. A. Antibody microinjection reveals an essential role for human polo-like kinase 1 (Plk1) in the functional maturation of mitotic centrosomes.
J. Cell Biol. 1996, 135 (6 Pt 2), 1701-1713.
(7) Sumara, I.; Gimenez-Abian, J. F.; Gerlich, D.; Hirota, T.; Kraft, C.; de la Torre, C.; Ellenberg, J.; Peters, J. M. Roles of polo-like kinase 1 in the assembly of
functional mitotic spindles. Curr. Biol. 2004, 14 (19), 1712-1722.
(8) Holtrich, U.; Wolf, G.; Brauninger, A.; Karn, T.; Bohme, B.; Rubsamen-Waigmann, H.; Strebhardt, K. Induction and down-regulation of PLK, a human serine/threonine kinase expressed in proliferating cells and tumors. Proc. Natl. Acad.
Sci. U. S. A. 1994, 91 (5), 1736-1740.
(9) Renner, A. G.; Dos Santos, C.; Recher, C.; Bailly, C.; Creancier, L.; Kruczynski, A.; Payrastre, B.; Manenti, S. Polo-like kinase 1 is overexpressed in acute myeloid leukemia and its inhibition preferentially targets the proliferation of leukemic cells.
Blood 2009, 114 (3), 659-662.
(10) Schoffski, P. Polo-like kinase (PLK) inhibitors in preclinical and early clinical development in oncology. Oncologist 2009, 14 (6), 559-570.
(11) Nappi, T. C.; Salerno, P.; Zitzelsberger, H.; Carlomagno, F.; Salvatore, G.; Santoro, M. Identification of Polo-like kinase 1 as a potential therapeutic target in anaplastic thyroid carcinoma. Cancer Res. 2009, 69 (5), 1916-1923.
(12) Strebhardt, K. Multifaceted polo-like kinases: drug targets and antitargets for cancer therapy. Nat. Rev. Drug Discov. 2010, 9 (8), 643-660.
(13) Weiss, L.; Efferth, T. Polo-like kinase 1 as target for cancer therapy. Exp.
(14) Liu, X.; Choy, E.; Harmon, D.; Yang, S.; Yang, C.; Mankin, H.; Hornicek, F. J.; Duan, Z. Inhibition of polo-like kinase 1 leads to the suppression of
osteosarcoma cell growth in vitro and in vivo. Anticancer Drugs 2011, 22 (5), 444-453.
(15) Guan, R.; Tapang, P.; Leverson, J. D.; Albert, D.; Giranda, V. L.; Luo, Y. Small interfering RNA-mediated Polo-like kinase 1 depletion preferentially reduces the survival of p53-defective, oncogenic transformed cells and inhibits tumor growth in animals. Cancer Res. 2005, 65 (7), 2698-2704.
(16) Yim, H. Current clinical trials with polo-like kinase 1 inhibitors in solid tumors. Anticancer Drugs 2013, 24 (10), 999-1006.
(17) Gleixner, K. V.; Ferenc, V.; Peter, B.; Gruze, A.; Meyer, R. A.;
Hadzijusufovic, E.; Cerny-Reiterer, S.; Mayerhofer, M.; Pickl, W. F.; Sillaber, C.; Valent, P. Polo-like kinase 1 (Plk1) as a novel drug target in chronic myeloid
leukemia: overriding imatinib resistance with the Plk1 inhibitor BI 2536. Cancer Res. 2010, 70 (4), 1513-1523.
(18) Ackermann, S.; Goeser, F.; Schulte, J. H.; Schramm, A.; Ehemann, V.; Hero, B.; Eggert, A.; Berthold, F.; Fischer, M. Polo-like kinase 1 is a therapeutic target in high-risk neuroblastoma. Clin. Cancer Res. 2011, 17 (4), 731-741.
(19) Steegmaier, M.; Hoffmann, M.; Baum, A.; Lenart, P.; Petronczki, M.; Krssak, M.; Gurtler, U.; Garin-Chesa, P.; Lieb, S.; Quant, J.; Grauert, M.; Adolf, G. R.; Kraut, N.; Peters, J. M.; Rettig, W. J. BI 2536, a potent and selective inhibitor of polo-like kinase 1, inhibits tumor growth in vivo. Curr. Biol. 2007, 17 (4), 316-322.
(20) Sebastian, M.; Reck, M.; Waller, C. F.; Kortsik, C.; Frickhofen, N.; Schuler, M.; Fritsch, H.; Gaschler-Markefski, B.; Hanft, G.; Munzert, G.; von Pawel, J. The efficacy and safety of BI 2536, a novel Plk-1 inhibitor, in patients with stage IIIB/IV non-small cell lung cancer who had relapsed after, or failed, chemotherapy: results from an open-label, randomized phase II clinical trial. J. Thorac. Oncol. 2010, 5 (7), 1060-1067.
(21) Mross, K.; Dittrich, C.; Aulitzky, W. E.; Strumberg, D.; Schutte, J.; Schmid, R. M.; Hollerbach, S.; Merger, M.; Munzert, G.; Fleischer, F.; Scheulen, M. E. A randomised phase II trial of the Polo-like kinase inhibitor BI 2536 in chemo-naive patients with unresectable exocrine adenocarcinoma of the pancreas - a study within the Central European Society Anticancer Drug Research (CESAR) collaborative network. Br. J. Cancer 2012, 107 (2), 280-286.
(22) Haupenthal, J.; Bihrer, V.; Korkusuz, H.; Kollmar, O.; Schmithals, C.; Kriener, S.; Engels, K.; Pleli, T.; Benz, A.; Canamero, M.; Longerich, T.;
Kronenberger, B.; Richter, S.; Waidmann, O.; Vogl, T. J.; Zeuzem, S.; Piiper, A. Reduced efficacy of the Plk1 inhibitor BI 2536 on the progression of hepatocellular
carcinoma due to low intratumoral drug levels. Neoplasia 2012, 14 (5), 410-419. (23) Muller-Tidow, C.; Bug, G.; Lubbert, M.; Kramer, A.; Krauter, J.; Valent, P.; Nachbaur, D.; Berdel, W. E.; Ottmann, O. G.; Fritsch, H.; Munzert, G.; Garin-Chesa, P.; Fleischer, F.; Taube, T.; Dohner, H. A randomized, open-label, phase I/II trial to investigate the maximum tolerated dose of the Polo-like kinase inhibitor BI 2536 in elderly patients with refractory/relapsed acute myeloid leukaemia. Br. J. Haematol. 2013, 163 (2), 214-222.
(24) Rudolph, D.; Steegmaier, M.; Hoffmann, M.; Grauert, M.; Baum, A.; Quant, J.; Haslinger, C.; Garin-Chesa, P.; Adolf, G. R. BI 6727, a Polo-like kinase inhibitor with improved pharmacokinetic profile and broad antitumor activity. Clin.
Cancer Res. 2009, 15 (9), 3094-3102.
(25) Grinshtein, N.; Datti, A.; Fujitani, M.; Uehling, D.; Prakesch, M.; Isaac, M.; Irwin, M. S.; Wrana, J. L.; Al-Awar, R.; Kaplan, D. R. Small molecule kinase
inhibitor screen identifies polo-like kinase 1 as a target for neuroblastoma tumor-initiating cells. Cancer Res. 2011, 71 (4), 1385-1395.
(26) Krause, M.; Kummer, B.; Deparade, A.; Eicheler, W.; Pfitzmann, D.; Yaromina, A.; Kunz-Schughart, L. A.; Baumann, M. Simultaneous PLK1 inhibition improves local tumour control after fractionated irradiation. Radiother. Oncol. 2013,
108 (3), 422-428.
(27) Wissing, M. D.; Mendonca, J.; Kortenhorst, M. S.; Kaelber, N. S.;
Gonzalez, M.; Kim, E.; Hammers, H.; van Diest, P. J.; Carducci, M. A.; Kachhap, S. K. Targeting prostate cancer cell lines with polo-like kinase 1 inhibitors as a single agent and in combination with histone deacetylase inhibitors. FASEB J. 2013, 27 (10), 4279-4293.
(28) Brassesco, M. S.; Pezuk, J. A.; Morales, A. G.; de Oliveira, J. C.; Roberto, G. M.; da Silva, G. N.; Francisco de Oliveira, H.; Scrideli, C. A.; Tone, L. G. In vitro targeting of Polo-like kinase 1 in bladder carcinoma: comparative effects of four potent inhibitors. Cancer Biol. Ther. 2013, 14 (7), 648-657.
(29) Pezuk, J. A.; Brassesco, M. S.; Morales, A. G.; de Oliveira, J. C.; de Paula Queiroz, R. G.; Machado, H. R.; Carlotti, C. G., Jr.; Neder, L.; Scrideli, C. A.; Tone, L. G. Polo-like kinase 1 inhibition causes decreased proliferation by cell cycle arrest, leading to cell death in glioblastoma. Cancer Gene. Ther. 2013, 20 (9), 499-506.
(30) Danovi, D.; Folarin, A.; Gogolok, S.; Ender, C.; Elbatsh, A. M.; Engstrom, P. G.; Stricker, S. H.; Gagrica, S.; Georgian, A.; Yu, D.; U, K. P.; Harvey, K. J.; Ferretti, P.; Paddison, P. J.; Preston, J. E.; Abbott, N. J.; Bertone, P.; Smith, A.; Pollard, S. M. A high-content small molecule screen identifies sensitivity of glioblastoma stem cells to inhibition of polo-like kinase 1. PLoS One 2013, 8 (10), e77053.
(31) Munch, C.; Dragoi, D.; Frey, A. V.; Thurig, K.; Lubbert, M.; Wasch, R.; Bogatyreva, L.; Hauschke, D.; Lassmann, S.; Werner, M.; May, A. M. Therapeutic polo-like kinase 1 inhibition results in mitotic arrest and subsequent cell death of blasts in the bone marrow of AML patients and has similar effects in non-neoplastic cell lines. Leuk. Res. 2015, 39 (4), 462-470.
(32) Rudolph, D.; Impagnatiello, M. A.; Blaukopf, C.; Sommer, C.; Gerlich, D. W.; Roth, M.; Tontsch-Grunt, U.; Wernitznig, A.; Savarese, F.; Hofmann, M. H.; Albrecht, C.; Geiselmann, L.; Reschke, M.; Garin-Chesa, P.; Zuber, J.; Moll, J.; Adolf, G. R.; Kraut, N. Efficacy and mechanism of action of volasertib, a potent and selective inhibitor of Polo-like kinases, in preclinical models of acute myeloid leukemia. J. Pharmacol. Exp. Ther. 2015, 352 (3), 579-589.
(33) Lin, C. C.; Su, W. C.; Yen, C. J.; Hsu, C. H.; Su, W. P.; Yeh, K. H.; Lu, Y. S.; Cheng, A. L.; Huang, D. C.; Fritsch, H.; Voss, F.; Taube, T.; Yang, J. C. A phase I study of two dosing schedules of volasertib (BI 6727), an intravenous polo-like kinase inhibitor, in patients with advanced solid malignancies. Br. J. Cancer 2014, 110 (10), 2434-2440.
(34) Janning, M.; Fiedler, W. Volasertib for the treatment of acute myeloid leukemia: a review of preclinical and clinical development. Future Oncol. 2014, 10 (7), 1157-1165.
(35) Stadler, W. M.; Vaughn, D. J.; Sonpavde, G.; Vogelzang, N. J.; Tagawa, S. T.; Petrylak, D. P.; Rosen, P.; Lin, C. C.; Mahoney, J.; Modi, S.; Lee, P.; Ernstoff, M. S.; Su, W. C.; Spira, A.; Pilz, K.; Vinisko, R.; Schloss, C.; Fritsch, H.; Zhao, C.; Carducci, M. A. An open-label, single-arm, phase 2 trial of the Polo-like kinase inhibitor volasertib (BI 6727) in patients with locally advanced or metastatic urothelial cancer. Cancer 2014, 120 (7), 976-982.
(36) Schoffski, P.; Awada, A.; Dumez, H.; Gil, T.; Bartholomeus, S.; Wolter, P.; Taton, M.; Fritsch, H.; Glomb, P.; Munzert, G. A phase I, dose-escalation study of the novel Polo-like kinase inhibitor volasertib (BI 6727) in patients with advanced solid tumours. Eur. J. Cancer 2012, 48 (2), 179-186.
(37) Gillet, J. P.; Gottesman, M. M. Mechanisms of multidrug resistance in cancer. Methods Mol. Biol. 2010, 596, 47-76.
(38) Wu, C. P.; Hsieh, C. H.; Wu, Y. S. The emergence of drug transporter-mediated multidrug resistance to cancer chemotherapy. Mol. Pharm. 2011, 8 (6), 1996-2011.
(39) Juliano, R. L.; Ling, V. A surface glycoprotein modulating drug
permeability in Chinese hamster ovary cell mutants. Biochim. Biophys. Acta. 1976,
455 (1), 152-162.
M. M. Targeting multidrug resistance in cancer. Nat. Rev. Drug Discov. 2006, 5 (3), 219-234.
(41) Gottesman, M. M.; Fojo, T.; Bates, S. E. Multidrug resistance in cancer: role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2 (1), 48-58.
(42) Burger, H.; van Tol, H.; Brok, M.; Wiemer, E. A.; de Bruijn, E. A.;
Guetens, G.; de Boeck, G.; Sparreboom, A.; Verweij, J.; Nooter, K. Chronic imatinib mesylate exposure leads to reduced intracellular drug accumulation by induction of the ABCG2 (BCRP) and ABCB1 (MDR1) drug transport pumps. Cancer Biol. Ther. 2005, 4 (7), 747-752.
(43) Hegedus, C.; Ozvegy-Laczka, C.; Szakacs, G.; Sarkadi, B. Interaction of ABC multidrug transporters with anticancer protein kinase inhibitors: substrates and/or inhibitors? Curr. Cancer Drug Targets 2009, 9 (3), 252-272.
(44) Mittapalli, R. K.; Vaidhyanathan, S.; Sane, R.; Elmquist, W. F. Impact of P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) on the brain distribution of a novel BRAF inhibitor: vemurafenib (PLX4032). J. Pharmacol. Exp.
Ther. 2012, 342 (1), 33-40.
(45) Wu, C. P.; Hsiao, S. H.; Sim, H. M.; Luo, S. Y.; Tuo, W. C.; Cheng, H. W.; Li, Y. Q.; Huang, Y. H.; Ambudkar, S. V. Human ABCB1 (P-glycoprotein) and ABCG2 mediate resistance to BI 2536, a potent and selective inhibitor of Polo-like kinase 1. Biochem. Pharmacol. 2013, 86 (7), 904-913.
(46) Wu, C. P.; Sim, H. M.; Huang, Y. H.; Liu, Y. C.; Hsiao, S. H.; Cheng, H. W.; Li, Y. Q.; Ambudkar, S. V.; Hsu, S. C. Overexpression of ATP-binding cassette transporter ABCG2 as a potential mechanism of acquired resistance to vemurafenib in BRAF(V600E) mutant cancer cells. Biochem. Pharmacol. 2013, 85 (3), 325-334.
(47) Wu, C. P.; Hsiao, S. H.; Luo, S. Y.; Tuo, W. C.; Su, C. Y.; Li, Y. Q.; Huang, Y. H.; Hsieh, C. H. Overexpression of human ABCB1 in cancer cells leads to reduced activity of GSK461364, a specific inhibitor of polo-like kinase 1. Mol.
Pharm. 2014, 11 (10), 3727-3736.
(48) Wu, C. P.; Hsiao, S. H.; Su, C. Y.; Luo, S. Y.; Li, Y. Q.; Huang, Y. H.; Hsieh, C. H.; Huang, C. W. Human ATP-Binding Cassette transporters ABCB1 and ABCG2 confer resistance to CUDC-101, a multi-acting inhibitor of histone
deacetylase, epidermal growth factor receptor and human epidermal growth factor receptor 2. Biochem. Pharmacol. 2014, 92 (4), 567-576.
(49) Shen, D. W.; Fojo, A.; Chin, J. E.; Roninson, I. B.; Richert, N.; Pastan, I.; Gottesman, M. M. Human multidrug-resistant cell lines: increased mdr1 expression can precede gene amplification. Science 1986, 232 (4750), 643-645.
(50) Currier, S. J.; Kane, S. E.; Willingham, M. C.; Cardarelli, C. O.; Pastan, I.; Gottesman, M. M. Identification of residues in the first cytoplasmic loop of
P-glycoprotein involved in the function of chimeric human MDR1-MDR2 transporters.
J. Biol. Chem. 1992, 267 (35), 25153-25159.
(51) Ambudkar, S. V. Drug-stimulatable ATPase activity in crude membranes of human MDR1-transfected mammalian cells. Methods Enzymol. 1998, 292, 504-514.
(52) Gribar, J. J.; Ramachandra, M.; Hrycyna, C. A.; Dey, S.; Ambudkar, S. V. Functional characterization of glycosylation-deficient human P-glycoprotein using a vaccinia virus expression system. J. Membr. Biol. 2000, 173 (3), 203-214.
(53) Wu, C. P.; Shukla, S.; Calcagno, A. M.; Hall, M. D.; Gottesman, M. M.; Ambudkar, S. V. Evidence for dual mode of action of a thiosemicarbazone,
NSC73306: a potent substrate of the multidrug resistance linked ABCG2 transporter.
Mol. Cancer Ther. 2007, 6 (12 Pt 1), 3287-3296.
(54) Hamada, H.; Tsuruo, T. Characterization of the ATPase activity of the Mr 170,000 to 180,000 membrane glycoprotein (P-glycoprotein) associated with
multidrug resistance in K562/ADM cells. Cancer Res. 1988, 48 (17), 4926-4932. (55) Dai, C. L.; Tiwari, A. K.; Wu, C. P.; Su, X. D.; Wang, S. R.; Liu, D. G.; Ashby, C. R., Jr.; Huang, Y.; Robey, R. W.; Liang, Y. J.; Chen, L. M.; Shi, C. J.; Ambudkar, S. V.; Chen, Z. S.; Fu, L. W. Lapatinib (Tykerb, GW572016) reverses multidrug resistance in cancer cells by inhibiting the activity of ATP-binding cassette subfamily B member 1 and G member 2. Cancer Res. 2008, 68 (19), 7905-7914.
(56) Brozik, A.; Hegedus, C.; Erdei, Z.; Hegedus, T.; Ozvegy-Laczka, C.; Szakacs, G.; Sarkadi, B. Tyrosine kinase inhibitors as modulators of ATP binding cassette multidrug transporters: substrates, chemosensitizers or inducers of acquired multidrug resistance? Expert Opin. Drug Metab. Toxicol. 2011, 7 (5), 623-642.
(57) Zhang, W.; Ling, V. Cell-cycle-dependent turnover of P-glycoprotein in multidrug-resistant cells. J. Cell. Physiol. 2000, 184 (1), 17-26.
(58) Cuestas, M. L.; Castillo, A. I.; Sosnik, A.; Mathet, V. L. Downregulation of mdr1 and abcg2 genes is a mechanism of inhibition of efflux pumps mediated by polymeric amphiphiles. Bioorg. Med. Chem. Lett. 2012, 22 (21), 6577-6579.
(59) Natarajan, K.; Bhullar, J.; Shukla, S.; Burcu, M.; Chen, Z. S.; Ambudkar, S. V.; Baer, M. R. The Pim kinase inhibitor SGI-1776 decreases cell surface expression of P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) and drug transport by Pim-1-dependent and -independent mechanisms. Biochem. Pharmacol. 2013, 85 (4), 514-524.
(60) Ludwig, J. A.; Szakacs, G.; Martin, S. E.; Chu, B. F.; Cardarelli, C.; Sauna, Z. E.; Caplen, N. J.; Fales, H. M.; Ambudkar, S. V.; Weinstein, J. N.; Gottesman, M. M. Selective toxicity of NSC73306 in MDR1-positive cells as a new strategy to circumvent multidrug resistance in cancer. Cancer Res. 2006, 66 (9), 4808-4815.
Nat. Rev. Cancer 2006, 6 (4), 321-330.
(62) Shukla, S.; Ohnuma, S.; Ambudkar, S. V. Improving cancer chemotherapy with modulators of ABC drug transporters. Curr. Drug. Targets 2011, 12 (5), 621-630.
(63) To, K. K.; Poon, D. C.; Chen, X. G.; Fu, L. W. Volasertib (BI 6727), a novel polo-like kinase inhibitor, reverses ABCB1 and ABCG2-mediated multidrug resistance in cancer cells. J. Cancer Ther. Res. 2013, 2 (13), 1-11.
(64) Ambudkar, S. V.; Cardarelli, C. O.; Pashinsky, I.; Stein, W. D. Relation between the turnover number for vinblastine transport and for vinblastine-stimulated ATP hydrolysis by human P-glycoprotein. J. Biol. Chem. 1997, 272 (34), 21160-21166.
(65) Ambudkar, S. V.; Dey, S.; Hrycyna, C. A.; Ramachandra, M.; Pastan, I.; Gottesman, M. M. Biochemical, cellular, and pharmacological aspects of the multidrug transporter. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 361-398.
FIGURE LEGENDS
Figure 1. The cytotoxicity of volasertib is reduced in cells overexpressing human ABCB1 protein. The cytotoxicity of volasertib in (A) sensitive human epidermal KB-3-1 cells (○) and the ABCB1-overexpressing sub-line KB-V-1 (●) as well as in (B) parental HEK293 (○) and ABCB1-tranfected MDR19-HEK293 (●) cells was determined as described previously.46 Points indicate the mean from at least three
independent experiments; bars indicate the SEM.
Figure 2. Volasertib-induced G2/M cell cycle arrest is reduced in
ABCB1-overexpressing cancer cells. Human KB-3-1 (top panels) and ABCB1-ABCB1-overexpressing KB-V-1 (lower panels) cells were plated and maintained in the absence or presence of volasertib alone or in combination with the ABCB1 inhibitor tariquidar or ABCB1 substrate nilotinib for 24 hrs before being harvested for cell-cycle analyses. (A) KB cells were treated with either DMSO (left panels), 1 µM tariquidar (middle panels), 5 µM nilotinib (right panels), (B) 10 nM volasertib (left panels), 10 nM volasertib and 1 µM tariquidar (middle panels) or 10 nM volasertib and 5 µM nilotinib (right panels). Representative histograms of three independent experiments are presented.
Figure 3. Volasertib-induced apoptosis is reduced in ABCB1-overexpressing cancer cells. KB-3-1 (top panels) and KB-V-1 (lower panels) cells were isolated 48 hrs after treatment with either (A) DMSO (left panels), 1 µM tariquidar (middle panels), 5 µM nilotinib (right panels), (B) 20 nM volasertib (left panels), 20 nM volasertib and 1 µM tariquidar (middle panels) or 20 nM volasertib and 5 µM nilotinib (right panels). Apoptotic cells were quantified by flow cytometry as described previously.47
presented.
Figure 4. Volasertib stimulates Vi-sensitive ABCB1 ATP hydrolysis consistent with transport by ABCB1. The effect of 0 to 20 µM and 0 to 2 µM (inset) volasertib on Vi-sensitive ABCB1 ATPase activity was measured as described previously.47 Points
indicate the mean of at least three independent experiments; bars indicate the SEM.
Figure 5. Volasertib inhibits the ABCB1-mediated efflux of calcein-AM. The accumulation of fluorescent calcein in (A) parental KB-3-1 (left panel) and KB-V-1 (right panel) cells as well as in (B) parental HEK293 (left panel) and ABCB1-transfected MDR19-HEK293 (right panel) cells in the presence (shaded, solid lines) or absence (solid lines) of 10 μM volasertib or 3 μM tariquidar (dotted lines) was measured by flow cytometry as described in the Experimental Section. Representative histograms of three independent experiments are shown. (C) The concentration-dependent inhibition of ABCB1-mediated calcein-AM efflux by volasertib in KB-V-1 cells was also determined. Data points represent the mean ± SEM from at least three independent experiments. The IC50 value was calculated based on the concentration that inhibited the efflux to 50% of the control values.
Figure 6. The effect of volasertib on ABCB1 protein expression in human KB epidermal cancer cells. (A) Human ABCB1 immunoblot detection and (B) quantification of total lysate protein (10 µg) from KB-V-1 cells treated with
increasing concentrations of volasertib for 72 hrs as described previously.47 α-tubulin
was used as an internal control for equal loading. Values are presented as the mean ± SD calculated from three independent experiments.
Cell line Cancer origin Transporter expressed Volasertib IC50 (nM) † RF‡ KB-3-1 epidermal - 1.5 ± 0.3 1 KB-C-1 epidermal ABCB1 6.5 ± 1.2** 4 KB-V-1 epidermal ABCB1 241.6 ± 38.6*** 161 OVCAR-8 ovary - 15.3 ± 3.0 1
NCI-ADR-RES ovary ABCB1 477.0 ± 129.5** 31
NIH3T3 - - 20.6 ± 3.3 1
NIH3T3-G185 - ABCB1 465.4 ± 132.9** 23
pcDNA-HEK293 - - 2.8 ± 0.8 1
MDR19-HEK293 - ABCB1 93.1 ± 12.3*** 33
† IC50 values are mean ± SD calculated from dose-response curves obtained from at least three independent experiments using a cytotoxicity assay as described in the
Experimental Section. * P < 0.05; ** P < 0.01; *** P < 0.001. ‡RF: resistance factors were calculated by dividing the IC50 values of ABCB1-expressing sub-line cells by the IC50 values of respective parental cells.
Table 2. Cell Cycle Distribution.
Cell cycle phase
G1 (%) S (%) G2/M (%) KB-3-1 Control 60.3 ± 4.8 28.4 ± 3.4 11.3 ± 1.8 + Tariquidar 65.3 ± 2.8 23.8 ± 2.4 11.0 ± 0.9 + Nilotinib 69.7 ± 6.0 20.3 ± 5.6 10.1 ± 0.8 + Volasertib 5.5 ± 2.3 13.1 ± 4.3 81.4 ± 6.3 + Volasertib + tariquidar 3.2 ± 1.6 10.7 ± 3.0 86.2 ± 4.6 + Volasertib + nilotinib 8.3 ± 4.6 18.2 ± 2.0 73.5 ± 6.6 KB-V-1 Control 62.8 ± 3.8 24.5 ± 2.0 12.7 ± 2.2 + Tariquidar 65.8 ± 3.9 24.1 ± 3.3 10.2 ± 0.9 + Nilotinib 65.5 ± 2.7 23.9 ± 2.5 10.6 ± 1.1 + Volasertib 57.5 ± 1.0 25.6 ± 3.3 16.9 ± 2.5 + Volasertib + tariquidar 4.0 ± 1.1 15.0 ± 3.5 80.5 ± 3.9 + Volasertib + nilotinib 15.6 ± 1.0 28.1 ± 2.7 56.4 ± 2.9 The values are mean ± SD calculated from three independent experiments.
Table 3. Effect of ABCB1 Modulators on the Chemosensitivity of Volasertib in ABCB1-Overexpressing Cell Lines.
Cell line Transporter overexpressed IC50 (nM) † Volasertib Volasertib + tariquidar (1 μM) Volasertib + nilotinib (0.5 μM) KB-3-1 - 1.5 ± 0.3 1.9 ± 0.4 2.4 ± 0.6 KB-V-1 ABCB1 241.6 ± 38.6 2.9 ± 0.5*** 32.5 ± 8.7*** HEK293 - 2.8 ± 0.8 1.1 ± 0.3* 1.6± 0.4 MDR19 ABCB1 93.1 ± 12.3 1.5 ± 0.5*** 19.2 ± 3.5*** † IC50 values are the mean ± SD calculated from dose-response curves obtained from at least three independent experiments using a cytotoxicity assay as described in the
Table 4. Chemosensitizing Effect of Volasertib on ABCB1-Mediated Drug Resistance in KB Human Epidermal Carcinoma Cells.
Treatment Concentration (nM) IC50 (nM) † KB-3-1 KB-V-1 Doxorubicin - 62.6 ± 20.6 (1) ‡ 2721.3 ± 228.7 (44) + Volasertib 1 86.1 ± 25.6 (1) 2928.5 ± 325.5 (47) + Volasertib 2 51.2 ± 16.3 (1) 3046.8 ± 575.1 (49) + Tariquidar 1000 49.4 ± 13.4 (1) 66.6 ± 10.4 (1)*** Colchicine - 5.0 ± 2.0 (1) 699.9 ± 64.4 (140) + Volasertib 1 4.9 ± 2.0 (1) 751.6 ± 92.2 (150) + Volasertib 2 4.0 ± 1.4 (1) 778.3 ± 93.9 (156) + Tariquidar 1000 4.8 ± 1.7 (1) 7.9 ± 1.9 (2)*** Paclitaxel - 2.9 ± 1.0 (1) 1675.5 ± 135.7 (578) + Volasertib 1 2.5 ± 0.8 (1) 1916.6 ± 161.9 (661) + Volasertib 2 1.6 ± 0.4 (1) 1929.4 ± 168.1 (665) + Tariquidar 1000 3.0 ± 0.9 (1) 2.3 ± 0.6 (1)*** † IC50 values are the mean ± SD calculated from dose-response curves obtained from three independent experiments using a cytotoxicity assay as described in the
Experimental Section. * P < 0.05; ** P < 0.01; *** P < 0.001. ‡R.R.: relative resistance values were calculated by dividing the IC50 values of ABCB1-expressing KB-V-1 cells by the IC50 values of parental KB-3-1 cells in the absence of volasertib or tariquidar.
For Table of Contents Only