Antimicrobial activity of the essential oil of Litsea cubeba on cariogenic
bacteria
Tsung-Shi Yanga, Ming-Li Lioub, Tzu-Fang Huc ,Chi-Wei Pengc, and Tai-Ti Liuc*
aDepartment of Cosmeceutics, China Medical University, No. 91, Hsueh-Shih Road,
Taichung 40402, Taiwan
bDepartment of Medical Laboratory Science and Biotechnology, Yuanpei University, No.
306 Yuanpei Street, Hsinchu 30015, Taiwan
cDepartment of Food Science, Yuanpei University, No. 306 Yuanpei Street, Hsinchu
30015, Taiwan
*Author to whom correspondence should be addressed (e-mail: [email protected], Tel: 886-3-5381183) 1 2 3 4 5 7 8 9 10 11 12 13 14 15
Abstract
The antimicrobial activity of the essential oil of Litsea cubeba (LC-EO) and the antimicrobial impact (AI) of individual volatile compounds in the LC-EO on the cariogenic bacteria: Streptococcus mutans (S. mutans), Streptococcus sobrinus (S.
sobrinus), and Streptococcus sanguinis (S. sanguinis) were studied. The AI combines the
effects of minimal microbicidal concentration (MMC) and quantity of an antimicrobial substance. The AI can quantitatively characterize the relative contribution of individual components of the EO on the entire antimicrobial activity of the EO. For the LC-EO and most volatile compounds investigated, their antimicrobial activities against S. sanguinis were higher than against S. mutans or S. sobrinus. The MMCs of the LC-EO for S.
sanguinis, S. mutans, and S. sobrinus without biofilms were 375, 750, and 1500 μg mL-1,
respectively. In contrast, the MMC for each strain with the biofilms was double the above value due to the increase of the bacterial resistance. The time-killing study shows that the LC-EO at the MMC for each strain could inactivate > 90% of the corresponding bacterial population (initial inoculum of 103 or 105 CFU mL-1) within 10 min. The good
antimicrobial effects within reasonable time of food consumption render the LC-EO applicable in food systems.
Key word Index:
Antimicrobial activity, antimicrobial impact, Litsea cubeba,Streptococcus mutans, Streptococcus sobrinus, Streptococcus sanguinis
16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Introduction
Because of the abuse of antibiotics and safety concern about synthetic antimicrobial agents, consumer health consciousness is prone to natural products. The recent interest in alternative naturally derived antimicrobials has lead to a renewed scientifc interest in essential oils (EOs) of plants (1). Litsea
cubeba (LC) is a plant widely distributed in Southeastern Asia, Southern China, Japan,
and Taiwan. LC can be used as a flavoring or herbal medicine. There have been some reports about the functional effects of LC such as treatment of illnesses (2), antioxidant activity (3), and antimicrobial activity (2). As a flavoring, the fruit of LC gives a unique flavor resembling that of a mixture of pepper, ginger, and citrus. LC is used as a flavor enhancer in foods, cosmetics, and cigarettes (4).
Regarding research in antimicrobial activity of EOs, most studies have investigated the effect of EOs as a whole (2, 5, 6)or certain major components (7-9). The individual compounds in an EO are structurally different and may have different chemical properties such as volatility and oxidative stability which should affect the whole antimicrobial activity of the EO during food handling, processing, and storage. Therefore, in the previous research we presented a concept of antimicrobial impact (AI), which combines the effects of the antimicrobial activities of individual compounds (indicated by MMC) and their amounts, to quantitatively reflect the relative importance of individual components of the EO on the entire antimicrobial activity of the EO (10). The AI may vary with the different microorganisms tested due to the variation of their MMCs and must be re-determined for new target microorganisms even using the same EO. In the 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58
present study, we would like to extend the application of the LC-EO to find out whether it can be used in the inhibition of harmful oral bacteria.
Maintaining good oral health is important for attaining enjoyment and quality in food consumption. The oral health is intimately associated with oral bacteria utilizing food residues in a mouth. Streptococcus mutans (S. mutans) and Streptococcus sobrinus (S. sobrinus) have been implicated as the prime causative organisms of human dental caries (11). Streptococcus sanguinis (S. sanguinis) can create friendly environment for other streptococci to grow such as S. mutans. For example, S. sanguinis can colonize and proliferate on tooth surfaces at an early stage of dental plaque formation (12). Co-culture of S. mutans with S. sanguinis yields firm adhesion of viable cells on smooth surfaces (13).Except for dental caries, malodor is another problem in oral hygiene. It has been reported that β-galactosidase activity in saliva is associated with oral malodor (14). Accordingly, S. sanguinis may contribute to the oral malodor via the β-galactosidase activity in deglycosylation of oral mucins (acidic glycoproteins) that may be a critical initial step leading to their subsequent proteolysis and putrefaction (15). More dangerously, S. sanguinis is the leading cause of infective endocarditis (16). Therefore, S. mutans, S. sobrinus, and S. sanguinis were used in this study as representative bacteria affecting the oral health. The antimicrobial activity of LC essential oil (LC-EO) and the antimicrobial impact of its individual compounds against S. mutans,
S. sobrinus, and S. sanguinis have not been studied.
Generally, most people seldom brush their teeth after meals especially the children. Therefore, if a flavoring such as LC-EO can be naturally incorporated into foods via cooking or into food products such as candies or chewing gums as a part of formulas, it 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81
may not only provide a flavor to the foods or food products but also readily impart a beneficial effect to the public oral health. Adhesion of a microorganism to tooth enamel is considered to be the frst step in pathogenesis (17). Milk proteins have been reported to have high levels of anti-adhesive activity against S.
mutans (17, 18). Milk is a common ingredient in a variety of candies. If the LC-EO and
milk proteins can be combined together in the candies, the anti-cariogenic effect should be greatly enhanced. Therefore, the aims of this study were to investigate the antimicrobial activity of the LC-EO and its individual compounds against S. mutans, S.
sobrinus, and S. sanguinis, and the flavor acceptance of the milk-candy with the LC-EO.
Experimental
Plant material
The fruit of LC was freshly harvested from the LC plants grown in the mountain areas in Jian-Shi town, Hsinchu, Taiwan. The fruit was stored in a freezer at -20°C after the sample collection. The fruit of LC was identified by morphological features and the database present in the Department of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, China Medical University, Taichung, Taiwan. A voucher specimen
(01-11-EO-LC-01) was preserved at the Department of Food Science, Yuanpei
University, Hsinchu, Taiwan. 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103
Chemicals
Tween 20, and Tween 80 were purchased from Sigma-Aldrich Co. (St. Louis, MO). Brain Heart Infusion broth (BHI broth) (Difco®), tryptic soy broth with 0.6% yeast extract (TSBYE)(Bacto™) were obtained from Becton Dickinson & Company (San Jose, CA, USA).
Bacterial strains
Streptococcus mutans (ATCC 25175), Streptococcus sobrinus (ATCC 14757), and Streptococcus sanguinis (ATCC 49295) were acquired from Bioresource Collection and
Research Center (BCRC) at the Food Industry Research and Development Institute, Hsinchu, Taiwan.
Isolation of essential oil
An amount of 100 g of LC fruit in fresh weight was ground and placed in a glass flask. Double de-ionized water was added into the flask to make a final volume of 1000 ml. The sample was subjected to hydrodistillation using a Clevenger-type apparatus. The vapor mixture of water-EO produced in the flask passed through a condenser and then the distillate was collected. The EO in the upper layer of the distillate was obtained and dried over anhydrous sodium sulfate.
104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125
Analysis of essential oil
The volume of 1μl of EO was injected into a gas chromatograph (GC, model 6890N, Agilent Technologies, Palo Alto, CA) equipped with a quadruple mass analyzer (MS, Agilent 5975). A capillary column (DB-WAX, 60 m x 0.25 mm x 0.25 μm, J &W Scientific, Folsom, CA) was used with helium as a carrier gas at a constant flow of 1 ml/min. The oven temperature was programmed as the following: initial holding at 40°C for 1 min, then 140 °C for 2 min at 5°C/ min, and 200 °C for 30 min at 3°C/ min. The MS operating parameters were the following: electron impact (EI) mode for molecular ionization with a voltage of 70 eV; ion source temperature, 200°C; total ion scan mode with a scan rate of 4.37 scans/s and mass scan range of 29-350 m/z (19). The volatiles
were identified by comparing their mass spectra with those in the library of MS data system (Wiley 275, G1035A, Agilent Technologies, Palo Alto, CA) and with those of standard compounds. The amounts of individual volatiles of the EO were expressed as percentages of the peak area relative to the total peak area obtained from a flame ionization detector at 250°C under the same GC conditions as above.
Antimicrobial activity assay
A stock LC-EO or volatile compound emulsion was prepared with a mixture of Tween 80 and Tween 20 (1: 1, w/w) and double de-ionized water at the LC-EO or volatile compound concentration of 24 mg/ml. The bacteria from stock cultures were inoculated 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148
in 5 ml broth and incubated at 37ºC for 48 h to increase their population. The bacterial number of the enriched cultures was controlled with dilution to optical density (OD) of 1 at 600 nm, which were further diluted with appropriate media for each strain to ca 105 colony forming units (CFU/mL) as an inoculum. The inoculum (0.5 ml) was
mixed with an emulsion with or without the EO or a volatile compound (1: 1, v/v), and added to a 12 ml test tube. The cultural medium was BHI broth for S. sanguninis and S.
sobrinus or TSBYE broth for S. mutans. The media for each strain were suggested by the
provider (BCRC) for optimal growth. The tubes were tightly capped and incubated for 48 h at 37˚C. The minimum microbicidal concentration (MMC) was determined by judging visually the bacterial growth in the tubes with a series of broth dilution, and then the existence of viable cells was verified by a plate count method. An inoculated growth medium without the tested compound was employed as a control. All experiments were performed in triplicate and the average values were reported as MMC. The MMC is defined as the lowest concentration of the EO or compound that results in no viable cells in a culture plate. The AI is calculated based on the MMC and component content. To know the ultimate antimicrobial ability of a compound, the MMC rather than minimal inhibitory concentration (MIC) was used in the calculation of the AI. The AI = [MMC (max) / MMC (component)] x component content (%), where the relative antimicrobial activity of MMC (max) is assumed to be unity [10]. The MMC (max) is the highest value of MMC used in a test; the MMC (component) is the MMC of a component that will be compared. For example, the MMC (max) is 12000 μg mL-1 in this study.
Kinetics of bacterial growth
149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171The bacterial samples were prepared as the above assay. The bacterial samples were incubated for 48 h at 37˚C and the OD at 600 nm was recorded at each time interval of sampling. The existence of viable cells was verified by a plate count method after 48 h of the incubation. The experiments were carried out in triplicate.
Treatment of established biofilms with LC-EO and volatile compounds
A bacterial suspension (0.5 ml) containing ca 105 CFU mL-1 was inoculated in a 12 ml
test tube. The tubes were tightly capped and incubated for 48 h at 37˚C to generate biofilms. After the biofilms were established, the cultural medium was decanted and the inside tube was rinsed 5 times with 1 ml phosphate-buffered saline (PBS) to remove the residual medium. The formation of the biofilms coated inside the wall of the tube was verified by staining them with 1 ml 1% crystal violet for 20 min. Then, the stained samples were washed 3 times with 1 ml PBS, and 1 ml 95% ethanol was added subsequently. The OD values of the supernatant were measured at 600 nm (20). Various amounts of the LC-EO and volatile compounds were added to the preformed biofilm samples containing S. sanguninis, S. sobrinus or S. mutans which were then incubated at 37˚C for another 48 h. The MMCs were determined as described above. The experiments were performed in triplicate.
Time-kill determination
172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193An aliquot of 30 ml bacterial cultures containing ca 103 or105 CFU mL-1
with or without the LC-EO at various concentrations were grown in 100 ml serum bottles at 37 °C. Colony counts were determined at 0, 10, 15, and 20 min by taking samples (1ml) at each time interval, following a serial of 10-fold dilutions in sterile 0.85% NaCl, and plating on appropriate agar plates as described above for each strain. Colonies were counted and averaged after 48 h of the incubation at 37 °C. The experiments were conducted in triplicate.
Sensory evaluation
Sensory evaluation of the milk candy treated with the LC-EO was assessed by a panel consisting of 25 untrained panelists, who were selected from students and staff in the Yuanpei University. The milk candy with the LC-EO was prepared as the following. Trehalose (26%, w/w), a low-cariogenic sugar (21), was mixed with milk (65%), butter (5%) and water (4%), and then heated mildly and slowly with quick hand-stirring to evaporate water. When the mixture turned sticky and was nearly ready for solidification, the LC-EO (750μg g-1), which was dissolved in part of melt butter in advance, was added
and well mixed. The milk candy was placed in a mold and stored in a refrigerator for solidification. The candy with a serving size of 15 g was used for sensory evaluation. The panelists were asked to evaluate overall acceptance of the samples via aroma and taste on a hedonic scale from 1 to 9, where 1: dislike extremely, 2: dislike very much, 3: dislike 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216
moderately, 4: dislike slightly, 5: neither like nor dislike, 6: like slightly, 7: like moderately, 8: like very much, and 9: like extremely (22).
Statistical analysis
Statistical analysis of variance (ANOVA) was conducted using Statistica for Windows (StatSoft, Tulsa, OK, USA). Fisher’s LSD test was used to compare the mean values of data for signifcant difference at a 95% level.
Results and Discussion
Effects of LC-EO on growth kinetics of S. mutans, S. sobrinus, and S.
sanguinis
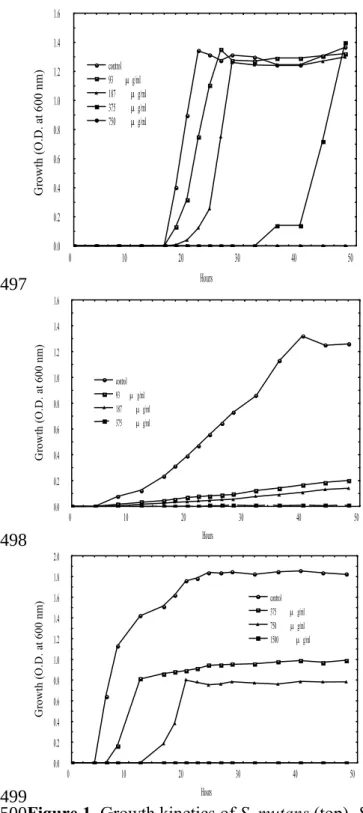
The growth kinetics of S. mutans, S. sobrinus, and S. sanguinis in the absence or presence of the LC-EO is shown in Figure 1. For S. mutans, detectable growth initiated at 18 h of the incubation and reached a maximum at 22 h. A marked antibacterial effect was observed at the LC-EO of 375 μg mL-1. The time of incipient bacterial growth was
postponed for ca 32 h at this concentration. S. mutans could be completely inactivated at the LC-EO of 750 μg mL-1.
Regarding S. sanguinis, its growth curve was different from that of S. mutans. The 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238
detectable growth of S. sanguinis without the LC-EO started at 8 h, which was earlier than that of S. mutans by 10 h. Afterward, S. sanguinis gradually grew and took 32 h to reach its maximal population. Comparatively, S. mutans only required 24 h to achieve the maximal growth. In the presence of the LC-EO at 93 μg mL-1, a noticeable inhibitory
effect was observed for S. sanguinis. Over 80% of S. sanguinis population could be inhibited by this concentration during 48 h of incubation. Moreover, the concentration of 375 μg mL-1 was required for complete inactivation of the bacteria.
As for S. sobrinus, the detectable growth of S. sobrinus without the LC-EO started at 6 h, which was earliest among the three strains and the growth reached a maximum at 24 h. The average growth rate was ca 3%/h in a time period of 6 - 24 h, which was also rapidest among the tested bacteria. About 43% of S. sobrinus survived the treatment of the LC-EO at 750 μg mL-1 after 48 h of the incubation. Complete inactivation of the S.
sobrinus required the LC-EO of 1500 μg mL-1.
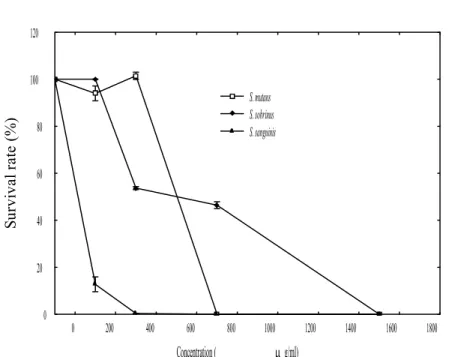
The growth of these strains in the absence or presence of the LC-EO after 48 h of the incubation was also investigated by the plate count method to verify their viability. The results are showed in Figure 2. The concentration of the LC-EO < 750 μg mL-1 could not
suppress the growth of S. mutans effectively because > 90% of the bacterial population restored their growth after 48 h of the LC-EO treatment. Regarding S. sanguinis, the survival rates of the bacteria in the presence of LC-EO at 93, 187, and 375 were 15±3, 13±2, and 0%, respectively. In addition, the survival rates of S. sobrinus with the LC-EO at 375, 750, and 1500 μg mL-1 were 53±3, 46±2, and 0%, respectively. These data were
consistent with the results obtained from the OD measurements. Accordingly, the MMCs for S. mutans, S. sobrinus, and S. sanguinis were determined as 750, 1500, and 375 μg 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261
mL-1, respectively.
The different antimicrobial activities of the LC-EO against the three strains may be related to the variation in the bacterial growth kinetics. Because S. sobrinus grew early and rapidly developed its population, which was largest among the three bacteria, the relatively high bacterial population may require a higher amount of antimicrobial substance to achieve complete inactivation.
Characterization of antimicrobial activity of LC against the streptococci
without biofilms based on individual volatile compounds
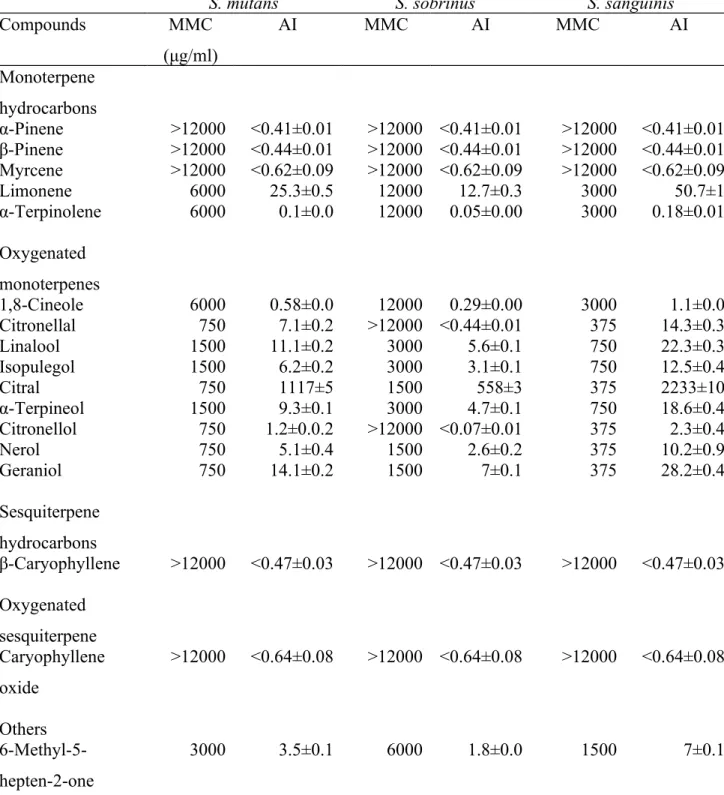
Effects of individual volatile compounds on the entire antimicrobial activity of an EO depend on the individual antimicrobial activity and their contents in the EO, which is combined as AI (10). The AI varies with the tested microorganisms due to the variation of MMCs. The identified volatile compounds of the LC-EO and their compositions are shown in Table 1. The AIs of each volatile compound in the LC-EO against these bacteria are listed in Table 2. Generally, The AIs of oxygenated monoterpenes were greater than those of monoterpene hydrocarbons, sesquiterpene hydrocarbons, or oxygenated sesquiterpene in this research. In addition, the MMCs of most compounds which showed antimicrobial activity for S. sobrinus were twice as high as that for S.
mutans. Similarly, the MMCs for S. mutans were also twice higher than that for S. sanguinis. Of monoterpene hydrocarbons, limonene and α-terpinolene had the same
MMC of 6000 μg mL-1 for S. mutans, 12000 μg mL-1 for S. sobrinus, and 3000 μg mL-1
for S. sanguinis, but the AI of limonene was greater than that of α-terpinolene. 262 263 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284
Regarding oxygenated monoterpenes, citronellal, citral, citronellol, nerol, and geraniol had the same MMC of 750 μg mL-1 for S. mutans and 375 μg mL-1 for S.
sanguinis. However, it was surprisingly found that citronellal and citronellol were nearly
ineffective against S. sobrinus. Except for the above case, these compounds displayed the strongest antimicrobial activity among the oxygenated monoterpenes. The AIs of the 5 compounds were in the descending order of citral, geraniol, citronellal, nerol, and citronellol. Comparatively, the AI of citral was markedly high because of its abundance (ca 70%) in the EO. Accordingly, citral was mainly responsible for ca 70% of total antimicrobial activity of the EO based on the ratio of its AI to the total AI. In terms of antibacterial mechanism, it is probably that citral affects the functions of cell membranes of these streptococci. Citral has been demonstrated to cause sublethal injury to the cytoplasmic and outer membrane of E. coli and L. monocytogenes (23, 24).
Linalool, isopulegol, and α-terpineol exhibited the same MMC of 1500, 3000, and 750 μg mL-1 against S. mutans, S. sobrinus, and S. sanguinis, respectively. Their AIs were
in the following order: linalool > α-terpineol > isopulegol. Linalool, α-terpineol, nerol, and geraniol are alcohol terpenoids and exhibited good antimicrobial effects against S.
mutans, S. sobrinus, and S. sanguinis in this study. As with other bactericidal alcohols,
the antibacterial mechanisms of these alcohol terpenoids may be due to protein denaturation or dehydration on the vegetative cells (25). In terms of stereochemistry, it has been suggested that cis-isomers are inactive contrary to trans-isomers (25). However, nerol (cis) and geraniol (trans) both had the same MMC for S. mutans, S. sobrinus, and S.
sanguinis in this research.
The presence of an acetate moiety in the structure appears to increase the activity of 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307
the parent compound. For example, the antimicrobial activity of geranyl acetate is greater that that of geraniol against the tested microorganisms such as Escherichia coli,
Pseudomonas aeruginosa, Staphylococcus aureus (25). In contrast, geranyl acetate was
inferior to geraniol in the inhibition of S. mutans, S. sobrinus, and S. sanguinis in this study. Because most compounds had MMCs for S. mutans greater than those for S.
sanguinis, it was surprisingly found that the MMC of geranyl acetate against S. mutans
was lower than that against S. sanguinis. In other words, geranyl acetate was more effective on S. mutans than on S. sanguinis. As for sesquiterpenes, β-caryophyllene and caryophyllene oxide had small AIs on the LC-EO due to their high MMCs and low quantities.
Antimicrobial activity of individual volatile compounds against the
streptococci with biofilms
Biofilm is a matrix-encased bacterial community and can provide a normal environment for most bacterial cells in many natural and artificial habitats. The biofilm may comprise complex associations of cells, extracellular products, and detritus either trapped within the biofilm or released from cells which have lysed as the biofilm ages. The main “cement” for all these cells and products is the mixture of polysaccharides secreted by the cells established within the biofilm (26). It cannot be generalized that the biofilm polysaccharides protect the bacterial cells within the biofilm. In fact, the protective effects of the biofilm polysaccharides often depend on conditions (26).Streptococci can form biolfims via secretion of extracellular polysaccharides. Therefore, the effect of the 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330
biofilms on the antimicrobial activity of the LC-EO and its components was also studied. The bacterial populations of S. sanguinis, S. sobrinus, and S. mutans with the established biofilms were ca 104 - 105 CFU mL-1 for the control samples after 48 h
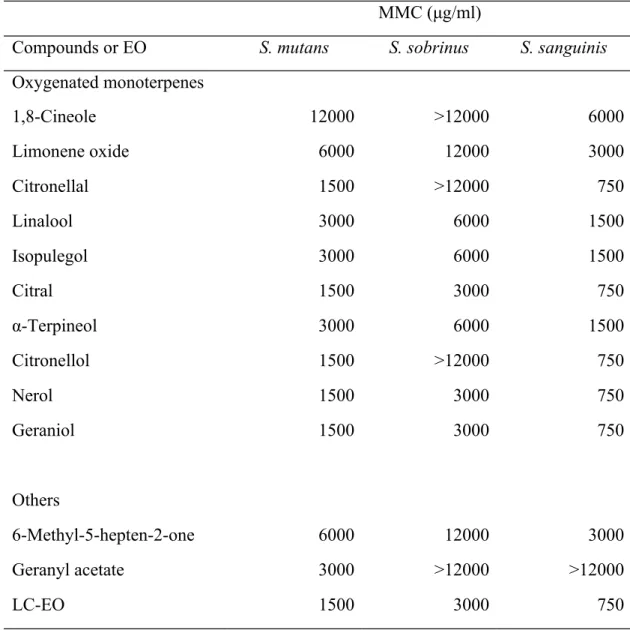
incubation at 37°C, which were similar to those in the control samples for those bacteria without the biofilms. However, the MMCs of the LC-EO and the selected volatile compounds for S. sanguinis, S. mutans, and S. sobrinus with the established biofilms were two-fold those of the bacteria without the biofilms (Table 3). These results demonstrated that the formation of the biofilms could increase the resistance of these bacteria to the LC-EO and its components. Despite the effect of the biofilms, the increase of MMCs against the bacteria with the biofilms were merely one level up as compared with that without the biofilms, which indicates that the LC-EO was still effective on inactivation of these bacteria.
Time-killing effect of LC-EO
To know the effect of the LC-EO on the cariogenic bacteria within reasonable eating time of normal food consumption, the time-killing effect of the LC-EO on S. mutans, S.
sobrinus, and S. sanguinis without the biofilms was determined. The number of mutans
streptococci > ca 105 CFU/ml is considered an infective dose to incur a risk of dental
caries [27]. Normally, the load of mutans streptococci in human mouths is lower than the infective dose. Therefore, two bacterial loads, ca 103 and105 CFU/ml, were tested in this
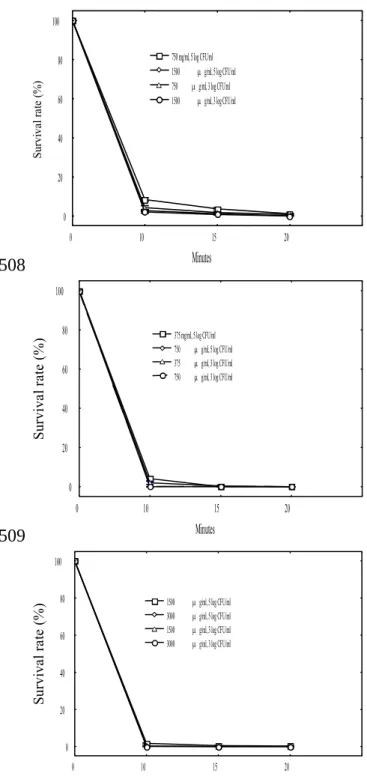
experiment. The results are shown in Figure 3. For S. mutans, the LC-EO at 750 μg mL-1
could kill ca 92% and 96% of the bacterial population with ca 105 CFU mL-1 and 103
331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353
CFU mL-1, respectively, within 10 min. Similarly, the LC-EO at 1500 μg mL-1 could
inactivate ca 98% and 100% of the population of S. sobrinus with 105 CFU mL-1 and 103
CFU mL-1, respectively. Regarding S. sanguinis, the LC-EO at 375 μg mL-1 could
eliminate ca 96% and 98% of the bacterial population at the same experimental conditions.
As a result, a lower concentration than the MMCs of the LC-EO should be sufficient to achieve a bactericidal or at least bacteriostatic effect in practice to control the bacterial population < 105 CFU mL-1. With the rapid inactivating effect, the LC-EO is suitable for
use in food systems. When the food is continuously eaten and chewed in a mouth, the released LC-EO can contact saliva and teeth, and lower the number of the cariogenic streptococci thereby reducing the risk of dental caries.
Sensory evaluation of milk candy with LC-EO
The sensory scores of the milk candies with and without the LC-EO were obtained as 7.4±1.6 and 7.1±1.8, respectively, which showed that both candies were moderately liked. Although the scores were not significantly different between the both candies (p<0.05), the average score of the milk candy with the LC-EO was slightly higher than that of the one without the LC-EO. The result manifests that the LC-EO can be compatibly used in the milk candy.
Conclusions
354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372 373 374 375 376The LC-EO has marked antimicrobial activities against the selected cariogenic streptococci and the antimicrobial activities are in the order of S. sanguinis > S. Mutans >
S. sobrinus. The good antimicrobial effects within reasonable time of food consumption
indicate that the LC-EO can be used to control these harmful oral bacteria via food systems such as the milk candy. For most of the effective volatile compounds tested, their antimicrobial activities against S. sanguinis are higher than against S. mutans or S.
sobrinus; however, geranyl acetate is more effective against S. mutans than against S. sanguinis. Citronellal and citronellol are very effective against S. mutans and S. sanguinis, whereas they are nearly ineffective against S. sobrinus. Sometimes, the LC-EO
may need a fine-tuning or even a greater modification of their original flavor notes to fit in with formulations of various products. The investigation of MMCs and AIs of individual compounds in the LC-EO can provide useful information for such purposes. For example, oxygenated monoterpenes such as citronellal, citronellol, nerol, and geraniol have comparable antimicrobial effects with citral in terms of MMC; however, their amounts are far lower than that of citral and result in low AIs in the LC-EO. Thus, these compounds may be selectively added into the EO via sensory evaluation to further enhance the entire antimicrobial activity of the EO.
377 378 379 380 381 382 383 384 385 386 387 388 389 390 391 392 393
References
1. J. Gutierrez, C. Barry-Ryan and P. Bourke, The antimicrobial efficacy of plant
essential oil combinations and interactions with food ingredients. Int. J. Food
Microbiol., 124, 91-97 (2008).
2. H. Wang and Y. Liu, Chemical composition and antibacterial activity of essential oils
from different parts of Litsea cubeba. Chem. Biodivers., 7, 229-235 (2010).
3. J. K. Hwang, E. M. Choi and J. H. Lee, Antioxidant activity of Litsea cubeba. Fitoterapia, 76, 684-686 (2005).
4. Y. Wang, Z.T. Jiang and R. Li, Complexation and molecular microcapsules of Litsea
cubeba essential oil with β-cyclodextrin and its derivatives. Eur. Food Res.
Technol., 228, 865-873 (2009).
5. B. Bozin, N. Mimica-Dukic, N. Simin and G. Anackov, Characterization of the
volatile composition of essential oils of some Lamiaceae spices and the antimicrobial and antioxidant activities of the entire oils. J. Agric. Food Chem., 54,
1822-1828 (2006).
6. D. Lopes-Lutz, D. S. Alviano, C. S. Alviano and P. P. Kolodziejczyk, Screening of
chemical composition, antimicrobial and antioxidant activities of Artemisia essential oils. Phytochemistry, 69, 1732-1738 (2008).
7. A. H. Ebrahimabadi, E. H. Ebrahimabadi, Z. Djafari-Bidgoli, F. J. Kashi, A. Mazoochi and H. Batooli, Composition and antioxidant and antimicrobial activity of the
essentialoil and extracts of Stachys inflata Benth from Iran. Food Chem., 119,
452-458 (2010). 394 395 396 397 398 399 400 401 402 403 404 405 406 407 408 409 410 411 412 413 414 415 416
8. J. Wang, H. Liu, J. Zhao, H. Gao, L. Zhou, Z. Liu, Y. Chen and P. Sui, Antimicrobial
and antioxidant activities of the root bark essential oil of Periploca sepium and its main component 2-hydroxy-4-methoxybenzaldehyde. Molecules, 15, 5807-5817
(2010).
9. R Shukla, P. Singh, B. Prakash and N. K. Dubey, Antifungal, aflatoxin inhibition and
antioxidant activity of Callistemon lanceolatus (Sm.) sweet essentialoil and its majorcomponent 1,8-cineole against fungal isolates from chickpea seeds. Food
Control., 25, 27-33 (2011).
10. T. T. Liu and T. S. Yang, Antimicrobial impact of the components of essential oil of Litsea cubeba from Taiwan and antimicrobial activity of the oil in food systems. Int. J. Food Microbiol., 156, 65-78
(2012).
11. H. Wu, M. Fan, X. Zhou, A. Mo, Z. Bian, Q. Zhang and Z. Chen, Detection of
Streptococcus mutans and Streptococcus sobrinus on the permanent first molars of the Mosuo people in China. Caries Res., 37, 374-380 (2003).
12.. C. G. Bloomquist, B. E. Reilly and W. F. Liljemark, Adherence, accumulation, and
cell division of a natural adherent bacterial population. J. Bacteriol. 178,
1172-1177 (1996).
13. M. Tamesada, S. Kawabata, T. Fujiwara and S. Hamada, Synergistic effects of
streptococcal glucosyltransferases on adhesive biofilm formation. J. Dent. Res., 83,
874-879 (2004).
14. N. Sterer, R. Bar-Ness Greenstein and M. Rosenberg, β-Galactosidase activity in
saliva is associated with oral malodor. J. Dent. Res., 81, 182-185 (2002).
417 418 419 420 421 422 423 424 425 426 427 428 429 430 431 432 433 434 435 436 437 438 439
15. J. S. Van der Hoeven and P. M. Camp, Synergistic degradation of mucin by
Streptococcus oralis and Streptococcus sanguis in mixed chemostat cultures. J.
Dent. Res., 70, 1041-1044 (1991).
16. J. E. Callahan and C. L. Munro, T Kitten, The Streptococcus sanguinis competence
regulon is not required for infective endocarditis virulence in a rabbit model. PloS
ONE 6: e26403 (2011).
17. R. M. Halpin, M. M. O’Connor, A. McMahon, C. Boughton, E. D. O’Riordan, M. O’Sullivan and D.B. Brady, Inhibition of adhesion of
Streptococcus mutans to hydroxylapatite by commercial dairy powders and individual milk proteins. Eur. Food Res. Technol., 227,
1499–1506 (2008).
18. E. A. Warner, A.D. Kanekanian and A.T. Andrews, Bioactivity of milk
proteins: 1. Anticariogenicity of whey proteins. Int. J. Dairy
Technol., 54, 151-153 (2001).
19. T. S. Yang, T. T. Liu and T. F. Hu, Effects of lecithin and pectin on riboflavin
photosensitized oxidation of orange oil in a multilayered oil-in-water emulsion. J.
Agric. Food Chem., 59, 9344-9350 (2011).
20. G. A. O'Toole and R. Kolter, Initiation of biofilm formation in Pseudomonas
fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a genetic analysis. Mol. Microbiol., 28, 449-461 (1998).
21. T. Neta, K Takada andM. Hirasawa, Low-cariogenicity of trehalose as a substrate. J. Dent., 28, 571-576 (2000). 440 441 442 443 444 445 446 447 448 449 450 451 452 453 454 455 456 457 458 459 460 461
22. M. Meilgaard, G..V. Civille and B.T. Carr, Sensory Evaluation Techniques, 2nd edn. CRC Press, Boca Raton, FL (1991).
23. M. Somolinos, D. García, R. Pagán and B. Mackey, Relationship between sublethal
injury and microbial inactivation by the combination of high hydrostatic pressure and citral or tert-butyl hydroquinone. Appl. Environ. Microbiol. 74, 7570-7577
(2008).
24. M. Somolinos, D. García, S. Condón, B. Mackey and R. Pagan, Inactivation of
Escherichia coli by citral. J. Appl. Microbiol., 108, 1928-1939 (2010).
25. H. J. D. Dorman and S. G. Deans, Antimicrobial agents from plants: antibacterial
activity of plant volatile oils. J. Appl. Microbiol., 88, 308-316 (2000).
26. I. W. Sutherland, Biofilm exopolysaccharides: a strong and sticky framework. Microbiol., 147, 3-9 (2001).
27. H. V. Jordan, R. Laraway, R. Snirch and M. Marmel, A simplified diagnostic system
for cultural detection and enumeration of Streptococcus mutans. J. Dent. Res., 66,
57-61 (1987).
28. S. Arctander, Perfume and flavor chemicals (aroma chemicals). Montclair, NJ., (1969). 462 463 464 465 466 467 468 469 470 471 472 473 474 475 476 477 478 479 480 481 482 483 484
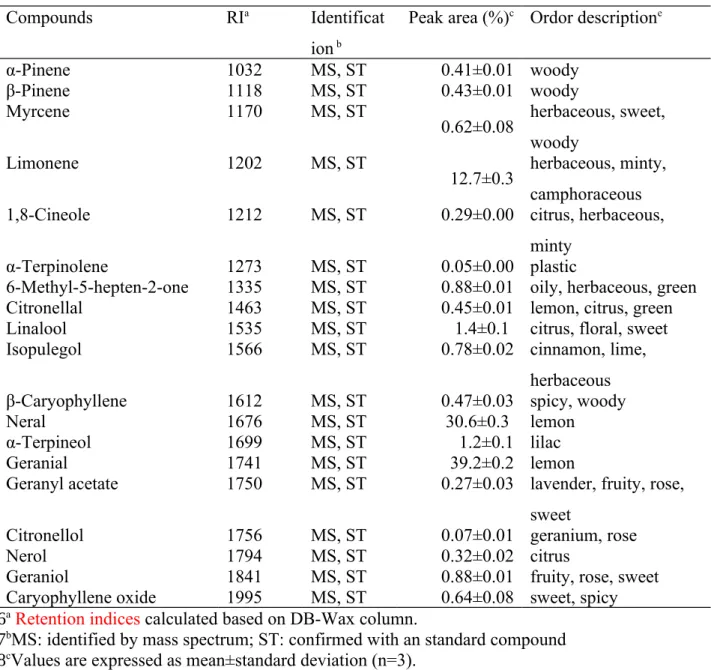
Table 1. Identification and composition of volatile compounds in the LC-EO
Compounds RIa Identificat
ion b
Peak area (%)c Ordor descriptione
α-Pinene 1032 MS, ST 0.41±0.01 woody β-Pinene 1118 MS, ST 0.43±0.01 woody Myrcene 1170 MS, ST 0.62±0.08 herbaceous, sweet, woody Limonene 1202 MS, ST 12.7±0.3 herbaceous, minty, camphoraceous
1,8-Cineole 1212 MS, ST 0.29±0.00 citrus, herbaceous,
minty
α-Terpinolene 1273 MS, ST 0.05±0.00 plastic
6-Methyl-5-hepten-2-one 1335 MS, ST 0.88±0.01 oily, herbaceous, green
Citronellal 1463 MS, ST 0.45±0.01 lemon, citrus, green
Linalool 1535 MS, ST 1.4±0.1 citrus, floral, sweet
Isopulegol 1566 MS, ST 0.78±0.02 cinnamon, lime,
herbaceous
β-Caryophyllene 1612 MS, ST 0.47±0.03 spicy, woody
Neral 1676 MS, ST 30.6±0.3 lemon
α-Terpineol 1699 MS, ST 1.2±0.1 lilac
Geranial 1741 MS, ST 39.2±0.2 lemon
Geranyl acetate 1750 MS, ST 0.27±0.03 lavender, fruity, rose, sweet
Citronellol 1756 MS, ST 0.07±0.01 geranium, rose
Nerol 1794 MS, ST 0.32±0.02 citrus
Geraniol 1841 MS, ST 0.88±0.01 fruity, rose, sweet
Caryophyllene oxide 1995 MS, ST 0.64±0.08 sweet, spicy
aRetention indices calculated based on DB-Wax column.
bMS: identified by mass spectrum; ST: confirmed with an standard compound cValues are expressed as mean±standard deviation (n=3).
e As reported in the reference[26]
485
486 487 488 489
Table 2. Minimal microbicidal concentration (MMC) and antimicrobial impact (AI) of individual compounds in the LC-EO on S. mutans, S. sobrinus, and S. sanguinis without the biofilms
S. mutans S. sobrinus S. sanguinis
Compounds MMC (μg/ml) AI MMC AI MMC AI Monoterpene hydrocarbons α-Pinene >12000 <0.41±0.01 >12000 <0.41±0.01 >12000 <0.41±0.01 β-Pinene >12000 <0.44±0.01 >12000 <0.44±0.01 >12000 <0.44±0.01 Myrcene >12000 <0.62±0.09 >12000 <0.62±0.09 >12000 <0.62±0.09 Limonene 6000 25.3±0.5 12000 12.7±0.3 3000 50.7±1 α-Terpinolene 6000 0.1±0.0 12000 0.05±0.00 3000 0.18±0.01 Oxygenated monoterpenes 1,8-Cineole 6000 0.58±0.0 12000 0.29±0.00 3000 1.1±0.0 Citronellal 750 7.1±0.2 >12000 <0.44±0.01 375 14.3±0.3 Linalool 1500 11.1±0.2 3000 5.6±0.1 750 22.3±0.3 Isopulegol 1500 6.2±0.2 3000 3.1±0.1 750 12.5±0.4 Citral 750 1117±5 1500 558±3 375 2233±10 α-Terpineol 1500 9.3±0.1 3000 4.7±0.1 750 18.6±0.4 Citronellol 750 1.2±0.0.2 >12000 <0.07±0.01 375 2.3±0.4 Nerol 750 5.1±0.4 1500 2.6±0.2 375 10.2±0.9 Geraniol 750 14.1±0.2 1500 7±0.1 375 28.2±0.4 Sesquiterpene hydrocarbons β-Caryophyllene >12000 <0.47±0.03 >12000 <0.47±0.03 >12000 <0.47±0.03 Oxygenated sesquiterpene Caryophyllene oxide >12000 <0.64±0.08 >12000 <0.64±0.08 >12000 <0.64±0.08 Others 6-Methyl-5-hepten-2-one 3000 3.5±0.1 6000 1.8±0.0 1500 7±0.1 490 491 492
Table 3. Minimal microbicidal concentration of the LC-EO and selected volatile compounds in the LC-EO on S. mutans, S. sobrinus, and S. sanguinis with the biofilms
MMC (μg/ml)
Compounds or EO S. mutans S. sobrinus S. sanguinis
Oxygenated monoterpenes 1,8-Cineole 12000 >12000 6000 Limonene oxide 6000 12000 3000 Citronellal 1500 >12000 750 Linalool 3000 6000 1500 Isopulegol 3000 6000 1500 Citral 1500 3000 750 α-Terpineol 3000 6000 1500 Citronellol 1500 >12000 750 Nerol 1500 3000 750 Geraniol 1500 3000 750 Others 6-Methyl-5-hepten-2-one 6000 12000 3000 Geranyl acetate 3000 >12000 >12000 LC-EO 1500 3000 750 493 494 495 496
0 10 20 30 40 50 Hours 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 G ro w th ( O .D . a t 6 00 n m ) control 93 g/ml 187 g/ml 375 g/ml 750 g/ml 0 10 20 30 40 50 Hours 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 G ro w th ( O .D . a t 6 00 n m ) control 93 g/ml 187 g/ml 375 g/ml 0 10 20 30 40 50 Hours 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 G ro w th ( O .D . a t 6 00 n m ) control 375 g/ml 750 g/ml 1500 g/ml
Figure 1. Growth kinetics of S. mutans (top), S. sanguinis (middle), and S. sobrinus
(bottom) in the absence or presence of increasing concentrations of the LC-EO. 497
498
499 500 501
0 200 400 600 800 1000 1200 1400 1600 1800 Concentration ( g/ml) 0 20 40 60 80 100 120 Su rv iv al r at e (% ) S. mutans S. sobrinus S. sanguinis
Figure 2. Effects of the LC-EO on the inactivation of S. mutans, S. sobrinus, and S. sanguinis after incubation at 37ºC for 48 h based on the plate counts of cultures.
502 503 504 505 506 507
0 10 15 20 Minutes 0 20 40 60 80 100 Su rv iv al r at e (% ) 750 mg/ml, 5 log CFU/ml 1500 g/ml, 5 log CFU/ml 750 g/ml, 3 log CFU/ml 1500 g/ml, 3 log CFU/ml 0 10 15 20 Minutes 0 20 40 60 80 100 S ur vi va l r at e (% ) 375 mg/ml, 5 log CFU/ml 750 g/ml, 5 log CFU/ml 375 g/ml, 3 log CFU/ml 750 g/ml, 3 log CFU/ml 0 10 15 20 Minutes 0 20 40 60 80 100 Su rv iv al r at e (% ) 1500 g/ml, 5 log CFU/ml 3000 g/ml, 5 log CFU/ml 1500 g/ml, 3 log CFU/ml 3000 g/ml, 3 log CFU/ml
Figure 3. Survival rates of S. mutans (top), S. sanguinis (middle), and S. sobrinus
(bottom) with two bacterial loads (5 log CFU/ml and 3 log CFU/ml) in the absence or presence of the LC-EO at various concentrations.
508 509 510 511 512 513 514 515 516
Figure captions
Figure 1. Growth kinetics of S. mutans (top), S. sanguinis (middle), and S. sobrinus
(bottom) in the absence or presence of increasing concentrations of the LC-EO.
Figure 2. Effects of the LC-EO on the inactivation of S. mutans, S. sobrinus, and S. sanguinis after incubation at 37ºC for 48 h based on the plate counts of cultures.
Figure 3. Survival rates of S. mutans (top), S. sanguinis (middle), and S. sobrinus
(bottom) with two bacterial loads (5 log CFU/ml and 3 log CFU/ml) in the absence or presence of the LC-EO at various concentrations.
517 518 519 520 521 522 523 524 525 526 527 528