1
中 國 醫 藥 大 學
專題研究計畫成果報告
計畫名稱:榖氨酸鹽於氨基乙酸受器上之蛋白質接合
點預測計算
計畫編號:CMU 100- TC - 05
執行期限:2011 年 10 月 20 日至 2012 年 06 月 19 日
單位名稱:生物科技學系
主持人:陳柏源、鍾景光
中 華 民 國 101 年 06 月 25 日
2 Introduction
As with release of neurotransmitters, the arrival of an action potential in the presynaptic terminal of an action potential in the presynaptic terminal of glycinergic neuron initiates a cascade involving vescular fusion and the release of glycine into the synaptic cleft. Glycine thus released is free to diffuse and bind with its receptors clustered on the postsynaptic face of adjacent cell.
Ligand-gated ion channels permit cells to respond rapidly to changes in their external environment. They are particularly well known for mediating fast neurotransmission in the nervous system. The glycine receptor (GlyR) is a membrane-embedded protein that contains an integral Cl- selective pore. When glycine binds to its site on the external receptor surface, the pore opens allowing Cl- to passively diffuse across the membrane. The GlyR is a member of the pentameric ligand-gated ion channel (LGIC) family, of which the nicotinic acetylcholine receptor cation channel (nAChR) is the prototypical member. Other members of this family include the cation-permeable serotonin type 3 receptor (5-HT3R), anion-permeable GABA type A and C receptors (GABAAR and GABACR), recently
identified cation-permeable zinc and GABA receptors [1, 2], as well as invertebrate anion-permeable glutamate and histidine receptors [3]. Note that glycine also directly activates a cation-selective ion channel of the excitatory glutamate receptor family [4]. The structural and functional properties of this receptor class have recently been reviewed [5] and are not considered here.
Glycine was first proposed as an inhibitory neurotransmitter on the basis of a detailed analysis of its distribution in the spinal cord [6]. Subsequent electrophysiological studies demonstrated a strychnine-sensitive hyperpolarizing action of glycine on spinal neurons [7, 8]. This hyperpolarization was soon discovered to be mediated by an increase in Cl- conductance. The receptors responsible for these actions were subsequently purified by strychnine affinity chromatography, and the first GlyR subunit was cloned in 1987 [9].
Glycine receptors are primarily restricted to the brain-stem and spinal cord. Like GABAA
receptors, the glycine receptor is a receptor ionophore that contains a Cl- channel. It is also similar in size to the GABAA receptor and is believed to possess a quasisymmetrical
pentameric structure that surrounds a water-filled ion conduction pore. Glycine receptors, which are unrelated to the glycine binding sites present on NMDA glutamate receptors, are defined pharmacologically by strychnine, a selective antagonist and a potent competitive and limited to a few ligands. The potent convulsants picrotoxin and picrotoxinin are non-competitive inhibitors of some of these receptors and believed to interact directly with the receptor’s ion channel to block Cl-
3
Current research into the GlyR can be divided into two major strands. The first involves the investigation of the molecular mechanisms of GlyR trafficking and clustering at synapses. This area is currently the subject of intense investigation, and recent progress has been covered in several authoritative reviews [10]. The second research strand is concerned with understanding the molecular structure and function of the GlyR. Research has intensified in this area over the past few years, and the purpose of this review is to present a coherent view of recent findings. Much of our understanding of GlyR structure-function has been gained by comparison with the structurally homologous nAChR.
Glutamate receptors comprise two large families, the iontropic and the metatropic receptors. Iontropic glutamate receptors contain associated ion channels that are gated by agonist binding. Three classes of iontropic glutamate receptors, N-methyl-D-asparate (NMDA), -amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), and kainate receptors, were originally named based on the ability of these drugs to serve as selective agonists.
Metabotropic glutamate receptors belong to the large superfamily of G protein-couple receptors. These receptors, which are characterized by seven transmembrane domains, couple to G proteins and in turn mediate the biologic effects of receptor activation. The term metatropic was used to indicate that these receptors affect cellular biochemical metabolic processes, and do not form ion channels. However, metatropic glutamate receptors, like other G protein-coupled receptors, can exert profound effects on neuronal function through the regulation of other ion channels, second messenger cascades, and protein phosphorylation. Neuronal excitability is fundamental to neuronal function, and is primarily controlled by a fine balance between synaptic excitation and inhibition. In the mammalian central nervous system, synaptic excitation is chiefly mediated by the excitatory transmitter glutamate acting on ionotropic glutamate receptor-gated cationic channels. In the mammalian brain, synaptic inhibition is primarily mediated by the inhibitory transmitter -aminobutyric acid (GABA), acting on the GABAA receptor-gated chloride channel. In contrast, synaptic inhibition is
principally mediated by glycine acting on the GABAA receptor-gated chloride channel [11].
In contrast, synaptic inhibition is principally mediated by glycine acting on the glycine receptor (GlyR)-gated chloride channel in the brainstem and spinal cord [12, 13]. However, in addition to being a primary inhibitory transmitter, glycine can also contribute to excitatory transmission by serving as an allosteric modulator for the N-methyl-D-asparate subtype of excitatory glutamate receptor (NMDAR)[14-16]. In the present study, Dr. Yu Tian Wang’s lab (UBC, Brain Research Centre) unexpectedly found that glutamate and its several ligand analogs allosterically potentiated GlyR mediated currents by interacting with
4
glutamate-binding site localized on the receptor subunit of GlyRs, along with the previously demonstrated glycine potentiation of excitatory NMDA receptors.
In this project, we predict the glutamate binding pocket on the GlyR by using SCRATCH: a protein structure and structural feature prediction server [17] to predict the 3D protein structure of GlyR 1 subunit-N-terminal from its 1D protein sequence. By using iGEMDOCK and Accelry Discovery Studio 2.5 to evaluate the binding pocket of glutamate on GlyR 1 subunit-N-terminal, the author can evaluate its interaction forces between them in order to realize the effect of allosteric potentiation.
Method
In this study, we will use these powerful tools to perform the visualized docking interaction between the receptor protein (GlyR) and the glutamate. First, we introduce SCRATCH: a protein structure and structural feature prediction server, iGEMDOCK and Accelry Discovery Studio 2.5 to understand the functions among them.
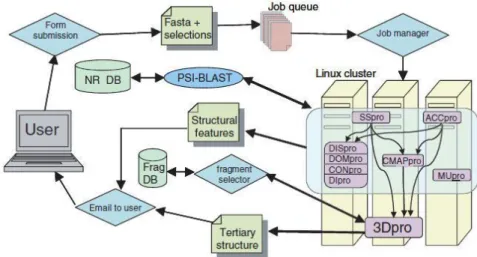
(i) SCRATCH: SCRATCH is a server for predicting protein tertiary structure and structural features. The SCRATCH software suite includes predictors for secondary structure, relative solvent accessibility, disordered regions, domains, disulfide bridges, single mutation stability, residue contacts versus average, individual residue contacts and tertiary structure. The user simply provides an amino acid sequence and selects the desired predictions, then submits to the server. Results are emailed to the user. The server is available at http://www.igb.uci.edu/servers/psss.html. Flow diagram for the SCRATCH server as illustrated in Fig.1 as below.
Fig. 1. Flow diagram for the SCRATCH server. DISpro, DOMpro, CONpro and DIpro are grouped together because they have the same inputs and their outputs are not used
5
by other predictors; however, they are standalone programs.
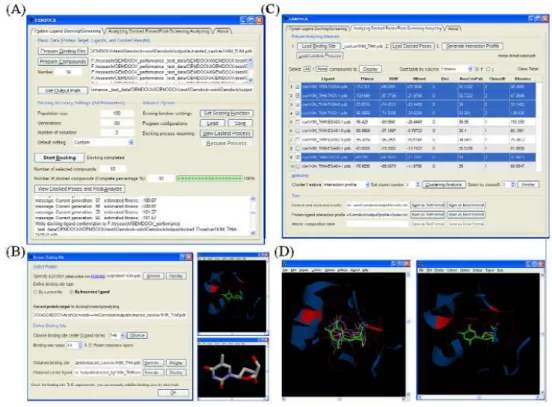
(ii) iGEMDOCK: iGEMDCOK is a suite of automated docking/screening tools. The interface of iGEMDOCK has two main tags, docking/screening tag and post-analyzing tag (shown as Fig. 2A and C, respectively). The docking/screening tag (Fig. 2A) is designed to predict how chemical molecules bind to a receptor of known 3D structure. The predicted protein-ligand poses can be further performed post-analysis in the post-analyzing tag (Fig. 2C). This can help, for example, to guide biological researchers explore better binders.
Fig. 2 The interface of iGEMDOCK
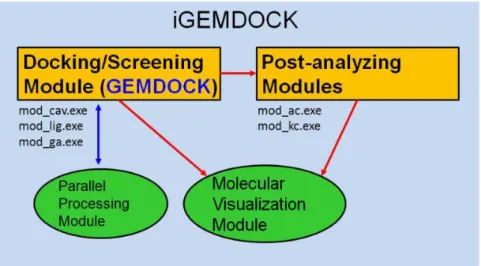
The architecture of iGEMDOCK consists of four major modules. The docking/screening and post-analyzing modules contain several components to make the screening/analyzing procedure smoothly. The predicted or clustered protein-ligand complexes can be visualized in the visualization module. The parallel processing module provides the parallel computation of screening jobs. The tools in the docking/screening and post-analyzing modules are further introduced as followed.
Components of docking/screening module
6
• mod_lig : this program generates the ligand list depending on user selected ligands.
• mod_ga : the main kernel (GEMDOCK) for docking/screening module. It works for the docking/screening process and predicts the bounded poses of protein-ligand complex.
Components of post-analyzing module
• mod_ac : atom composition program analyzes the atom composition and environment of each docked pose.
• mod_kc : k-mean cluster groups the docked poses by their interaction or atom composition descriptors (Fig. 3).
Fig. 3 Components of docking/screening module of iGEMDOCK
(iii) Accelry Discovery Studio 2.5: By offering a wealth of tools within a graphical interface, Discovery Studio makes it easier than ever to examine the properties of large and small molecules, study systems, identify leads, and optimize candidates. The science at Discovery Studio’s core has undergone years of continuous innovation based on the input of customers, internal scientists, and renowned scientific advisors. With citations in over hundreds of research publications, Discovery Studio is a trusted resource for many of the industry’s top research teams.
Discovery Studio is part of the open and scalable Accelrys Scientific Informatics Platform. The features of this powerful platform will: (1) Make better decisions by integrating, processing, and simultaneously considering diverse data types, from protein sequences and structures to molecular properties and experimental data. (2) Streamline the work environment and optimize resource utilization by integrating all the tools into the platform, from in-house code, to third-party software, to public or internal databases. (3) Process data efficiently by automating tasks and leveraging parallel computing. (4) Ensure best practices are enforced with algorithm
7
customization and workflow sharing.
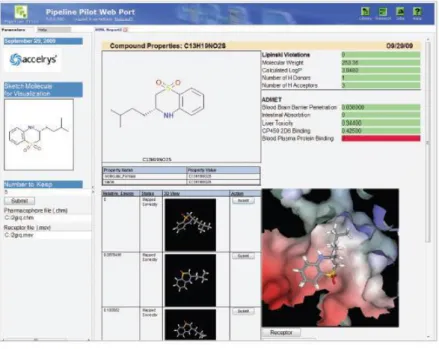
In this project, we ultilize this powerful tools to perform the visualized docking interation between the receptor protein (GlyR)and the glutamate (see Fig. 4).
Fig. 4: Access to complex science from a user-friendly, web-based interface makes it easy for medicinal chemists to visualize and explore the complex relationship between biological and chemical experimental data using both 2D and 3D data.
(iv) HADOCK v.2.0 have experimented previously with the inclusion of water in the NMR structure calculation of a protein-non-specific DNA complex: in that case, an extensive set of NOEs together
Procedure:
Step 1: SCRATCH:
It is well known that combining predictors usually improves prediction accuracy. In this work, we use SCRATCH to perform protein 3D structure prediction. Current methods for predicting secondary structure typically combine multiple neural networks, sometimes several hundreds of them, trained more or less independently. Combination of different systems rather than networks has also been used. At the alignment level, the ability to produce profiles that include increasingly remote homologs using PSI-BLAST has also contributed to performance improvement. Divergent evolutionary profiles contain not only enough
8
information to substantially improve prediction accuracy but even to correctly predict long stretches of identical residues observed in alternative secondary structure states depending on nonlocal conditions. An example is a method automatically identifying structural switches, and thus finding a remarkable connection between predicted secondary structure and aspects of function. Finally, at the algorithmic level, new bidirectional recurrent neural network architectures in combination with BLAST profiles to produce a first-generation secondary structure predictor SSpro 1.0.
Step 2: iGEMDOCK
The detail of iGEMDOCK for the protein–ligand docking is as following descriptions. iGEMDOCK, an automatic docking tool, is able to generate all experimental variables and serve as a flexible or hybrid-solution docking tool. We designed a new rotamer-based mutation operator for reducing the search space of ligand structure conformations, and used a differential evolution operator for reducing the disadvantages of Gaussian and Cauchy mutations. First, they specified the coordinates of ligand and protein atoms, the ligand binding area, atom formal charge, and atom types. Crystal coordinates of the ligand and protein atoms were taken from the Protein Data Bank, and were separated into different files. The size and location of the ligand binding site was determined by considering the protein atoms located 10 Å from each ligand atom when preparing the proteins. iGEMDOCK then automatically determined the center of the receptor and the search cube of a binding site according to the maximum and minimum of coordinates of these selected protein atoms.
Step 3. HADDOCK v.2.0
ADDOCK v.2.0 is a solvated docking approach that explicit accounts for the presence of water in protein-protein complexes. The solvated docking protocol is based on the concept of the first encounter complex in which a water layer is present in-between the molecules. Docking is performed from solvated biomolecules and waters are removed in a biased Monte Carlo procedure based on water mediated contact propensities obtained from an analysis of high-resolution crystal structure.
Results and Discussion
In the mammalian brain stem and spinal cord, Synapse is a mechanism of action that is mediated by excitatory and inhibitory excitability grain acid, it acts on the ions - grain acid receptor-gated cation channels and inhibits glycine roleglycine receptor (GlyR)-gated chloride channel.
9
The literature indicates that the possible role of grain acid and several analog ligand in the glycine receptor-gated chloride ion currents of spinal neurons,it would increase the open rate of single ion channel without any known ion receptor activation or metabotropic glutamate receptors. From past studies and found that the grain acid may act on the glycine receptor (GlyR)-gated chloride channel.
Therefore, this study was applicant Accelry Discovery Studio 3.0 software to calculate and predict the protein junction position of grain acid glycine receptor and the force nature. It is going to facilitate verification grains acid may act on the glycine receptor (GlyR) gated chloride the molecular mechanisms of ion channels.
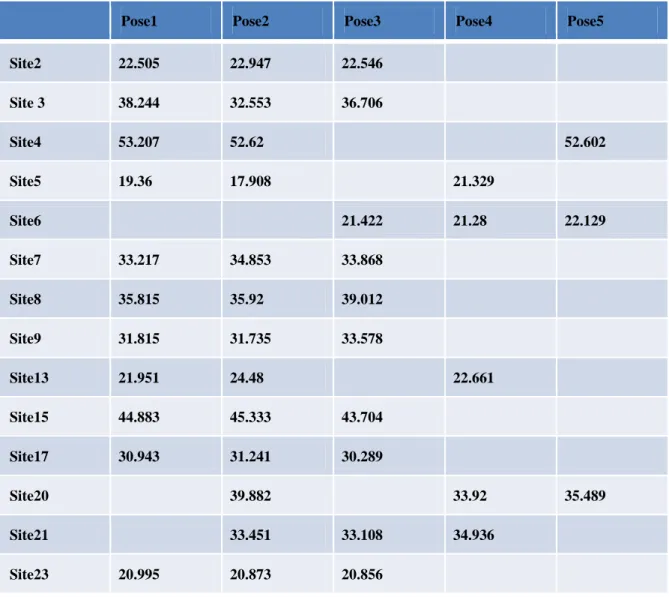
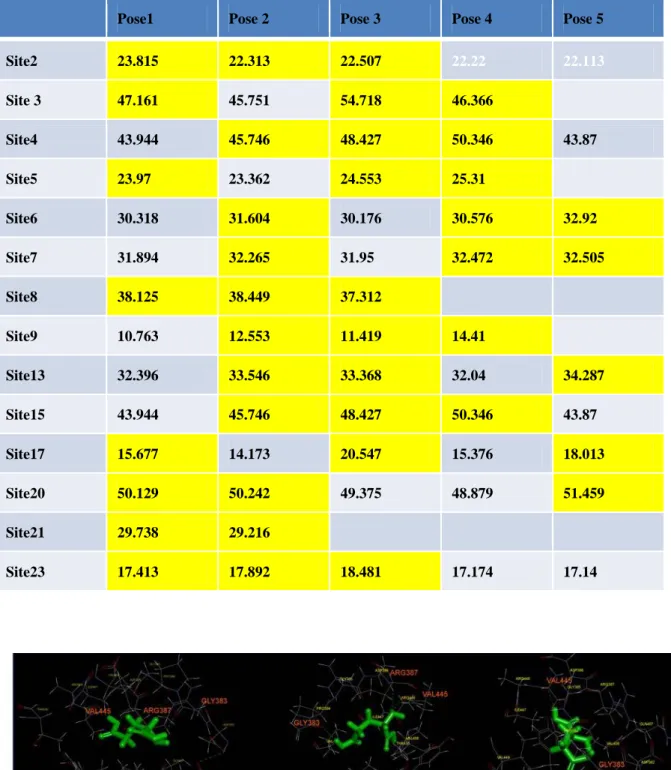
We searched Gly alpha 1 of the Protein Data Bank and found the experiment-related protein (Code: 1T3E) using Accelry Discovery Studio 3.0 do the docking simulation (Table 1 and Table 2).
Table 1. Anhydrous environment
Pose1 Pose2 Pose3 Pose4 Pose5
Site2 22.505 22.947 22.546 Site 3 38.244 32.553 36.706 Site4 53.207 52.62 52.602 Site5 19.36 17.908 21.329 Site6 21.422 21.28 22.129 Site7 33.217 34.853 33.868 Site8 35.815 35.92 39.012 Site9 31.815 31.735 33.578 Site13 21.951 24.48 22.661 Site15 44.883 45.333 43.704 Site17 30.943 31.241 30.289 Site20 39.882 33.92 35.489 Site21 33.451 33.108 34.936 Site23 20.995 20.873 20.856
15
Table 2. Hydrous environment
Pose1 Pose 2 Pose 3 Pose 4 Pose 5 Site2 23.815 22.313 22.507 22.22 22.113 Site 3 47.161 45.751 54.718 46.366 Site4 43.944 45.746 48.427 50.346 43.87 Site5 23.97 23.362 24.553 25.31 Site6 30.318 31.604 30.176 30.576 32.92 Site7 31.894 32.265 31.95 32.472 32.505 Site8 38.125 38.449 37.312 Site9 10.763 12.553 11.419 14.41 Site13 32.396 33.546 33.368 32.04 34.287 Site15 43.944 45.746 48.427 50.346 43.87 Site17 15.677 14.173 20.547 15.376 18.013 Site20 50.129 50.242 49.375 48.879 51.459 Site21 29.738 29.216 Site23 17.413 17.892 18.481 17.174 17.14
20
In the calculation of anhydrous environment, it is convenient to calculate because there is no interference of the water molecules. If you really want to understand the combined effect of proteins and drugs in the in vivo environment better, you will need water molecules to do most of the environmental factors, and then use the program to simulate proteins and drugs with the affinity of the status that is affected by water molecules.But by adding water molecules influence and calculate docking of protein and drug, will may affect the docking scores. So the docking scores that were obtained may be more worst than the original.
References
1. Beckstead MJ, Weiner JL, Eger EI II, Gong DH, and Mihic SJ. Glycine and -aminobutyric acidA receptor function is enhanced by inhaled drugs of abuse. Mol Pharmacol 57: 1199–1205, 2000.
2. Davies PA, Wang W, Hales TG, and Kirkness EF. A novel class of ligand-gated ion channel is activated by Zn2+. J Biol Chem 278: 712–717, 2003.
3. Gisselmann G, Pusch H, Hovemann BT, and Hatt H. Two cDNAs coding for histamine-gated ion channels in D. melanogaster. Nat Neurosci 5: 11–12, 2002.
4. Chatterton JE, Awobuluyi M, Premkumar LS, Takahashi H, Talantova M, Shin Y, Cui J, Tu S, Sevarino KA, Nakanishi N, Tong G, Lipton SA, and Zhang D. Excitatory glycine receptors containing the NR3 family of NMDA receptor subunits. Nature 415: 793–798, 2002.
5. Dingledine R, Borges K, Bowie D, and Traynelis SF. The glutamate receptor ion channels.
Pharmacol Rev 51: 7–61, 1999.
6. Beg AA and Jorgensen EM. EXP-1 is an excitatory GABA-gated cation channel. Nat
Neurosci 6: 1145–1152, 2003.
21
1502–1503, 1967.
8. Werman R, Davidoff RA, and Aprison MH. Inhibition of motoneurones by iontophoresis of glycine. Nature 214: 681–683, 1967.
9. Grenningloh G, Rienitz A, Schmitt B, Methfessel C, Zensen M, Beyreuther K, Gundelfinger ED, and Betz H. The strychninebinding subunit of the glycine receptor shows homology with nicotinic acetylcholine receptors. Nature 328: 215–220, 1987. 10. Kneussel M and Betz H. Clustering of inhibitory neurotransmitter receptors at developing
postsynaptic sites: the membrane activation model. Trends Neurosci 23: 429–435, 2000. 11. Mody, I., De Koninck, Y., Otis, T. S. and Soltesz, I. Bridging the cleft at GABA synapses
in the brain. Trends Neurosci 17: 517-525, 1994.
12. Betz, H. and Laube, B. Glycine receptors: recent insights into their structural organization and functional diversity. J Neurochem 97: 1600-1610, 2006.
13. Lynch, J. W. Molecular structure and function of the glycine receptor chloride channel.
Physiol Rev 84: 1051-1095, 2004.
14. Johnson, J. W. and Ascher, P. Glycine potentiates the NMDA response in cultured mouse brain neurons. Nature 325: 529-531, 1987.
15. Kleckner, N. W. and Dingledine, R. Requirement for glycine in activation of NMDA-receptors expressed in Xenopus oocytes. Science 241: 835-837, 1988.
16. Dingledine, R., Kleckner, N. W. and McBain, C. J. The glycine coagonist site of the NMDA receptor. Adv Exp Med Biol 268: 17-26, 1990.
17. Cheng, J. Randall, A. Z. Sweredoski, M. J. and Baldi, P. SCRATCH: a protein structure and structural feature prediction server. Nucleic Acids Research, 33: W72–W76, 2005.