Areca Nut Extracts Attenuated Interferon-
γ and
Antigen-specific IgM Production in BALB/c Mice
CHIA-CHI WANG1,2, MING-CHUNG DENG3 AND TONG-RONG JAN1*
1. Department and Graduate Institute of Veterinary Medicine, School of Veterinary Medicine,
National Taiwan University, Taipei, Taiwan, R.O.C.
2. School of Pharmacy, Kaohsiung Medical University, Kaohsiung, Taiwan, R.O.C.
3. Animal Health Research Institute, Council of Agriculture, Executive Yuan, New Taipei City, Taiwan, R.O.C.
(Received: January 3, 2011; Accepted: June 23, 2011)
ABSTRACT
Areca-nut chewing has been well established as a major risk factor for oral cancer and precancerous diseases, whose pathophysi-ology has been associated with immune deterioration. Previous studies reported that areca nut extracts (ANE) affected the functionality of lymphocytes and neutrophils in vitro. We recently showed that intraperitoneal administration of ANE modulated antigen-specific immunity and promoted inflammatory reactions in ovalbumin (OVA)-sensitized mice. The objective of the present studies was to further investigate the in vivo effect of ANE on T cell-mediated immune responses. In non-sensitized mice, a single oral dose of ANE (200 mg/kg) markedly suppressed the production of interferon (IFN)-γ by splenocytes stimulated with the T cell mitogen concanavalin A, whereas the expression of interleukin (IL)-2 or IL-4 was unaltered. In OVA-sensitized mice, daily administration of ANE (200 mg/kg) for 9 days significantly suppressed the antigen-induced production of IFN-g by splenocytes and the serum level of antigen-specific IgM. Both the cellularity and the metabolic activity of splenocytes were unaffected by the ANE treatment. Collectively, these results demonstrated that oral administration of ANE modulated antigen-specific T cell responses. As IFN-g is a key cytokine involved in the activation of various immunocompetent cells, ANE-mediated suppression of IFN-γ production may be a critical mechanism contributing to the immune deterioration associated with areca-related oral diseases.
Key words: Areca nut, T-cell, interferon-γ, antigen-specific
INTRODUCTION
Areca quid (AQ) chewing is a popular habit in Taiwan, India, and several other Asian countries with approximately 600 million chewers around the world(1). AQ chewing has been well established as a major etiological factor for oral cancer and precancerous lesions, including oral leukoplakia and oral submucous fibrosis (OSF)(1,2). In addition to oral diseases, AQ chewing is also a risk factor of hepatocel-lular carcinoma(3), suggesting that AQ chewing may cause systemically deteriorating effects with a broad spectrum of outcomes.
To date, the mechanisms responsible for the broad spectrum of areca-associated toxic effects in AQ chewers are mostly unclear. Clinical studies have documented that immune dysfunction is closely associated with the patho-physiology of areca-related oral diseases(4,5). For example, daily consumption of AQ has been shown correlated with
the deregulation of humoral immunity in OSF patients(6). In addition, cell-mediated immunity has been reported compro-mised in patients with areca-associated oral diseases(5).
T helper (Th) cells play a pivotal role in the host immu-nity against tumor formation(7). In particular, the Th1 cyto-kine interferon-gamma (IFN-γ) is a key cytocyto-kine involved in both innate and acquired immunity, such as the promotion of Th1 cell differentiation, the activation of natural killer cells, cytotoxic T lymphocytes and macrophages, and the stimulation of antibody production(8). Notably, immuno-histochemical studies revealed that Th cells are the major immunocompetent cells present in OSF tissues as evidenced by an elevated ratio of CD4+ to CD8+ cells(9,10). However, the expression of IFN-γ in the OSF tissues was suppressed, indicating a down-regulation of T-cell functionality(10). Concordantly, a decreased ability of peripheral blood mono-nuclear cells isolated from OSF patients to produce IFN-γ was also observed(11). Collectively, these lines of evidence demonstrates that the reactivity of T cells, such as the expres-sion of IFN-γ, is down-regulated, which may contribute to * Author for correspondence. Tel: +886-2-33661287;
at least one week. The animal was housed in a temperature (22 ± 2°C), humidity (50 ± 20%), and light-controlled envi-ronment (12 h light/dark cycle) with free access to standard laboratory food and water.
III. ANE Administration Protocol (I) Non-Sensitized Protocol
The mice were either left untreated (naïve; NA), or administered by gavage with a single dose of ANE (200 mg/kg; 0.2 mL/mouse) and/or H2O (vehicle; VH; 0.2 mL/mouse). The mice were sacrificed 3 h post drug administration and their splenocytes were isolated for further experiments (Figure 1A).
(II) OVA-Sensitized Protocol
The mice were either left untreated (NA), or sensitized by intraperitoneal injection with a sensitization solution containing OVA plus alum (100 μg/1 mg; OVA) on day 3. The OVA-sensitized mice were administered once per day the dysfunction of cellular immunity in areca-associated oral
diseases.
AQ is generally comprised of areca nut (Areca catechu), piper betle leaf (Piper betle), and lime paste(1). As clinical reports have shown alterations in many aspects of the immune system in areca-associated oral diseases, the direct influence of areca nut extract (ANE) ingredients on immune cells is recently under intensive investigation. It has been reported that ANE inhibited the phagocytosis and bactericidal activity of neutrophils(12). The inhibition by areca ingredi-ents on T-cell activation has also been demonstrated(13). We previously showed that ANE inhibited the metabolic activity and IFN-γ production by murine splenocytes in vitro(14). In addition, ANE enhanced the production of inflammatory cytokines in peripheral blood mononuclear cells(15). Intra-peritoneal administration of ANE promoted inflammatory reactions associated with delayed-type hypersensitivity and modulated antigen-specific antibody production in oval-bumin (OVA)-sensitized mice(16). Together these studies clearly demonstrate that lymphocytes are one of the sensitive targets to ANE. However, evidence pertaining to the in vivo effect of ANE on antigen-specific immunity remains limited and to be further addressed.
The objective of the present study is to investigate the effects of oral administration of ANE on antigen-induced T cell responses. As AQ is consumed by chewing, the oral route of administration is a more relevant mode of exposure compared to the intraperitoneal route. We report here that oral administration of ANE markedly attenuates the expression of the Th1 signature cytokine IFN-γ in both non-sensitized and antigen-sensitized mice. In addition, ANE suppresses the serum production of antigen-specific immunoglobulin M (IgM), substantiating the modulatory effect of ANE on acquired immunity.
MATERIALS AND METHODS I. Reagents and Areca Nut Extract
All reagents were purchased from Sigma (St. Louis, MO, USA) unless otherwise stated. Areca nuts were extracted with water as previously described(17), and the extracts were freeze-dried as ANE. The ANE has been confirmed endo-toxin-free using a commercial Limulus amebocyte lysate assay kit (Kinetic-QCL®; Lonza Walkersville Inc., Walkers-ville, MD, USA). Fetal bovine serum (FBS) and RPMI 1640 medium were from Hyclone (Logan, UT, USA). Reagents for ELISA were from BD Biosciences (San Diego, CA, USA). II. Animals
Male BALB/c mice, 5 - 6 weeks of age were purchased from the Animal Breeding Center of the National Taiwan University Hospital (Taipei, Taiwan). On arrival, the mice were randomized, transferred to plastic cages containing a saw-dust bedding (five mice per cage) and quarantined for
Figure 1. Protocols for OVA-sensitization and ANE administration
to BALB/c mice. (A) In the non-sensitized protocol, male BALB/c mice were randomly divided into the following groups: naïve (NA), vehicle (VH; ddH2O) and ANE-treated (200 mg/kg). The mice were
administered with VH and ANE by gavage. The splenocytes were isolated 3 h after ANE administration for further experimentation. (B) In the OVA-sensitized protocol, mice were randomly divided into the following groups: naïve (NA), OVA-sensitized (OVA), vehicle-treated and OVA-sensitized (VH), and ANE-vehicle-treated (200 mg/kg) and OVA-sensitized (ANE). Except for the NA group, mice were sensi-tized with OVA plus alum (100 μg/1 mg) on day 3. ANE and VH were administered daily by gavage from day 1 - 9. All mice were sacrificed 3 h after the final drug treatment, and their serum and splenocytes were obtained for further experimentation.
(B) Ovalbumin (OVA)-sensitized protocol
ANE and/or VH (ddH2O) Day 1 2 3 4 5 6 7 8 9 OVA/Albumin Sensitization ANE and/or VH (ddH2O)
(A) Non-sensitized protocol
Time 0 h
Sacrifice 3 h
with ANE (200 mg/kg; 0.2 mL/mouse; ANE) and/or H2O (VH; 0.2 mL/mouse) by gavage from day 1 - 9. All mice were sacrificed 3 h after the final dose of ANE, and their serum and splenocytes were isolated for further experi-ments (Figure 1B). The animal experiexperi-ments were approved by the Institutional Animal Care and Use Committee of the National Taiwan University.
IV. Spleen Index
After sacrifice, the spleen of each mouse was dissected out and weighed immediately. The spleen index was calcu-lated as the spleen weight (mg) per body weight (g).
V. Cellularity of Splenocytes
Splenocytes were stained with rat anti-mouse CD4 conjugated with fluoroscein isothiocyanate (FITC), rat anti-mouse CD8 and B220 conjugated with PE-Cy5 antibodies (BioLegend, San Diego, CA, USA) in phosphate-buffered saline (PBS) containing 2% FBS. After washing, the single cell fluorescence of 10,000 cells for each sample was measured by a flow cytometer (BD FACSCalibur, San Jose, CA, USA). Data were analyzed using the software Flowjo 5.7. VI. Measurement of The Metabolic Activity of Splenocytes
Splenocytes were cultured in RPMI 1640 medium supplemented with 100 U/mL penicillin, 100 μg/mL strep-tomycin and 5% heat-inactivated FBS. In all cases, sple-nocytes were cultured at 37°C in 5% CO2. Splenocytes (5 × 106 cells/mL) were cultured in quadruplicate in 96-well plates (100 μL/well) and stimulated with concanavalin A (ConA; 5 μg/mL) for 48 h or OVA (50 μg/mL) for 72 h. The metabolic activity of splenocytes was determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) assay. Splenocytes were cultured 4 more h in the presence of MTT (50 μg/mL) and the formed formazan was dissolved with a lysis buffer (10% SDS in N,N-demethylformamide) overnight in the dark. The plate was read on a microplate reader at 570 nm using 630 nm as the background reference.
VII. Cytokine Measurement
Splenocytes (5 × 106 cells/mL) were cultured in quadruplicate in 48-well plates (300 μL/well) and stimulated with ConA for 48 h or OVA for 72 h. After stimulation, the supernatants from each well were quantified for interleukin (IL)-2, IFN-γ and IL-4 by standard sandwich enzyme-linked immunosorbent assay (ELISA) as previously described(14). VIII. Measurement of OVA-Specific IgM
ELISA plates were coated with 0.05% OVA in coating buffer (0.1 M NaHCO3) and blocked with 1% bovine serum albumin in PBS containing 0.05% Tween-20 (PBST). After
washing with PBST, serum samples were added into wells and incubated for 2 h. After washing, radish peroxidase-conjugated anti-mouse IgM was added and incubated for 1 h. The wells were washed again and a tetramethylben-zidine solution was added for colorimetric detection on a microplate reader.
RESULTS
I. ANE Attenuated IFN-γ Production by Splenocytes Stimulated with ConA
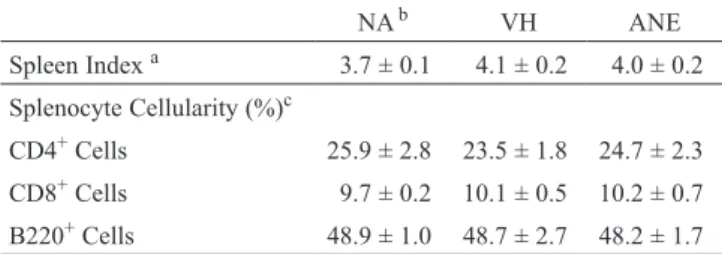
T cell-derived cytokines participate in both humoral and cell-mediated immune responses. We therefore exam-ined the effect of ANE on the expression of IL-2, a pivotal cytokine for T-cell clonal expansion, and IL-4 and IFN-γ that are signature cytokines produced by Th1 and Th2 cells, respectively, in non-sensitized mice. The dose (200 mg/kg) of ANE was used according to previous reports showing the in vivo effect of ANE to inhibit mast cell degranulation(18), and to induce the sister chromatin exchange of bone marrow chromosomes(19). Mice were sacrificed 3 h post a single dose of ANE administration (Figure 1A), and their splenocytes were stimulated with the T-cell mitogen concanavalin A (ConA; 5 μg/mL) for 48 h to induced cytokine expression. The production of IFN-γ was significantly attenuated by the ANE treatment (Figure 2A), whereas IL-2 and IL-4 were unaffected (Figure 2B and C). The effect of ANE on cyto-kine production may be due to alterations in T cell cellularity and/or activity. Thus, the influence of ANE on the spleen index, cellularity and the metabolic activity of splenocytes was examined. The ANE treatment exerted no effect either on the spleen index, cellularity (Table 1) or the metabolic activity of splenocytes activated by ConA (Figure 2D). II. ANE Attenuated Antigen-Specific IFN-γ Production
As ANE differentially suppressed ConA-induced IFN-γ production by splenocytes of non-sensitized mice,
Table 1. Effect of ANE on spleen index and splenocyte cellularity
NA b VH ANE Spleen Index a 3.7 ± 0.1 4.1 ± 0.2 4.0 ± 0.2 Splenocyte Cellularity (%)c CD4+ Cells 25.9 ± 2.8 23.5 ± 1.8 24.7 ± 2.3 CD8+ Cells 9.7 ± 0.2 10.1 ± 0.5 10.2 ± 0.7 B220+ Cells 48.9 ± 1.0 48.7 ± 2.7 48.2 ± 1.7 a Spleen index was calculated as the spleen weight (mg) per body
weight (g). Data are expressed as mean ± SE of four samples and are representative of three independent experiments.
b NA: untreated; VH: vehicle-treated and OVA-sensitized and
chal-lenged and ANE: ANE-treated and OVA-sensitized and chalchal-lenged.
c The percentage of CD4+, CD8+ and B220+ cells was determined
by flow cytometry. Data are expressed as mean ± SE of triplicate samples pooled from three independent experiments.
we thus investigated whether ANE affected antigen-specific IFN-γ production. Splenocytes of OVA-sensitized mice were re-stimulated with OVA (50 μg/mL) in culture for 72 h to induce OVA-specific cytokine production. The OVA re-stimulation successfully induced IFN-γ production (Figure 3; OVA re-stimulation vs. no stimulation), which was significantly attenuated by repeated administration of ANE (Figure 3; ANE vs. VH). None of the metabolic activity of splenocytes, the spleen index and cellularity was affected by ANE treatment (data not shown).
III. ANE Attenuated OVA-Specific IgM Production
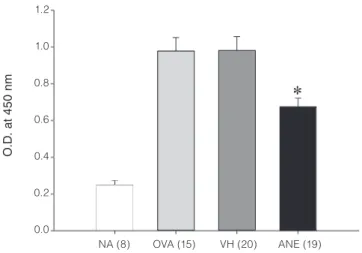
Antibody production is one of the hallmark responses in the acquired immunity. Cytokines derived from Th cells play a pivotal role in the activation of antibody-producing B cells. As ANE administration attenuated OVA-specific IFN-γ production, we therefore examined the effect of ANE on the serum level of OVA-specific IgM, the major isotype of immunoglobulin produced during the primary antibody
response. Our data showed a marked increase in the serum OVA-specific IgM in OVA-sensitized mice indicating a successful induction of antibody response, which was significantly attenuated by repeated administration of ANE (Figure 4).
DISCUSSION
Although immune dysfunction has been reported to be associated with the pathophysiology of areca-related oral diseases(5), evidence pertaining to the in vivo effect of areca nut constituents on antigen-specific immunity is scarce. The present study demonstrated that oral administration of ANE markedly attenuated the production of IFN-γ by splenocytes in both non-sensitized and antigen-sensitized mice, which is in line with our previous report showing the suppression of IFN-g production by splenocytes directly exposed to ANE in vitro(14). As one of the major sources for IFN-g is Th1 cells, these results suggest that ANE differentially affects the
Figure 2. Attenuation of IFN-g production by ANE administration in non-sensitized mice. Mice were treated with ANE as described in Figure
1A. Splenocytes (5 × 106 cells/mL) were stimulated with concanavalin A (ConA; 5 mg/mL) for 48 h. (A-C) The concentration of IFN-g, IL-2
and IL-4 in the supernatants was measured by ELISA. (D) The metabolic activity of viable cells was determined using the MTT assay. Data are expressed as the mean ± SE of quadruplicate cultures. Results are representative of six independent experiments. *p < 0.05 compared to the VH
group. 160 140 120 100 80 60 40 20 0 NA VH ANE IL -4 (p g /m L) 140 120 100 80 60 40 20 0 NA VH ANE IF N -γ ( ng /m L) 1.4 1.2 1.0 0.8 0.6 0.4 0.2 0.0 NA VH ANE O .D . (5 70 /6 50 nm ) 6 5 4 3 2 1 0 NA VH ANE IL -2 (n g /m L) No Stimulation ConA (A) (C) (B) (D) No Stimulation ConA No Stimulation ConA No Stimulation ConA
functions of Th1 cells. Furthermore, we showed a marked attenuation of the serum production of antigen-specific IgM in OVA-sensitized mice treated with ANE. On the base of these results, we postulated that the attenuation of the acquired immunity by ANE may contribute to the clinical observed immune dysfunction in patients with areca-related oral diseases(5).
Immune responses mediated by the Th1 signature cyto-kines IFN-g are critical for the host defense against microbes and tumor cells(20,21). IFN-γ can activate the killing activity
of natural killer cells, cytotoxic T cells and macrophages, and promote the production of opsonizing and complement-fixing antibodies that facilitate the phagocytosis of foreign antigens(20,22). Lacking IFN-γ in knockout mice increased the incidence of lymphoma, clearly indicating its pivotal role in cancer immunosurveillance(21). Interestingly, T lympho-cytes have been identified as the major immunocompetent cells in the subepithelial connective tissue of OSF patients(9), and the expression of IFN-γ in the oral cavity of OSF patient was suppressed compared to normal controls(23). Further-more, the ability of peripheral blood mononuclear cells to produce IFN-γ was diminished in patients with oral leuko-plakia, OSF, or head and neck cancers(11,24,25). Given the crit-ical role of IFN-g in immunosurveillance, down-regulation of the capability of Th1 cells to express IFN-g could be a critical mechanism in the pathophysiology of areca-related oral diseases. The disturbances of Th1 cell functions may be caused by immunosuppressive mediators released by tumor cells(26). Alternatively, as shown in the present studies, contribution from the immunomodulatory effect caused by areca constituents may be another critical mechanism to suppress the functionality of T cells.
An increased production of autoantibodies and circu-lating immune complexes has been documented in patients with areca-related oral diseases(27,28). Using the OVA model, we showed that the production of antigen-specific IgM was attenuated by oral administration of ANE, which is consistent with our previous report showing a similar effect by intra-peritoneal administration of ANE(16). It is currently unclear whether ANE attenuated IgM via a direct effect on B cells or an indirect mechanism mediated by cytokine suppression. This issue requires further investigations.
Arecoline at the dose between 5 - 20 mg/kg has been shown to affect splenic cellularity and antigen-specific immune responses(30,31). However, the amount of arecoline contained in our ANE is < 10 mg/mg of ANE (data not shown). Hence, the dose of arecoline corresponding to 200 mg of ANE/kg is < 2 mg/kg that is much lower than the reported effective dose range of arecoline. Moreover, our data showed that ANE administration did not affect splenic cellularity. Therefore, it is apparent that arecoline may not play a major role in the present studies. In addition to alkaloids, areca nuts contain a significant amount of polyphenols, in particular procyanidins(29). A growing body of evidence suggests that procyanidins exhibit a broad spectrum of biological effects, such as immunomodulation(32-34). For example, highly oligo-meric procyanidins purified from Jatoba herb ameliorated encephalomyelitis and collagen-induced arthritis in mice via suppression of Th1 immunity, including suppression of IFN-γ production and inhibition of macrophage maturation(33,34) Therefore, procyanidins might be the active constituents contributing to the immunosuppressive effect of ANE.
In summary, the present studies demonstrated that oral administration of ANE attenuated the production of IFN-γ and OVA-specific IgM. These results indicate that areca nuts contain constituents that can be orally absorbed and produce immunomodulatory effects. Areca-mediated
Figure 3. Attenuation of antigen-induced IFN-g production by ANE
administration in OVA-sensitized mice. Mice were treated with ANE as described in Figure 1B. Splenocytes (5 × 106 cells/mL) were
stim-ulated with OVA (50 mg/mL) for 72 h, and the concentration of OVA-specific IFN-γ in the supernatants was measured by ELISA. Data are expressed as the mean ± SE of quadruplicate cultures. Results are representative of four independent experiments. *p < 0.05 compared to the matched VH group.
30 25 20 15 10 5 0 NA OVA VH ANE IF N -γ ( ng /m L) No Stimulation OVA (50 µg/mL)
Figure 4. Attenuation of the serum production of OVA-specific IgM
by ANE administration. Mice were treated with ANE as described in Figure 1B. The serum titer of OVA-specific IgM was determined by ELISA. Data are expressed as the mean ± SE of pooled data from four independent experiments. The number of individual sample for each group is indicated in the parenthesis.*p < 0.05 compared to the matched VH group. 1.2 1.0 0.8 0.6 0.4 0.2 0.0 NA (8) OVA (15) VH (20) ANE (19) O .D . at 4 50 nm
down-regulation of IFN-γ production may be a potential mechanism involved in the compromised acquired immunity associated with areca-related oral diseases.
ACKNOWLEDGMENTS
This work was supported by grant NSC98-2320-B-002-036-MY3 from the National Science Council, and 99AS-9.2.5-BQ-B1(2) from the Council of Agriculture, Executive Yuan, Taiwan.
REFERENCES
1. Nair, U., Bartsch, H. and Nair, J. 2004. Alert for an epidemic of oral cancer due to use of the betel quid substitutes gutkha and pan masala: a review of agents and causative mechanisms. Mutagenesis 19: 251-262. 2. IARC. 2004. Betel-quid and areca-nut chewing and
some areca-nut derived nitrosamines. IARC Monogr. Eval. Carcinog. Risks. Hum. 85: 1-334.
3. Tsai, J. F., Chuang, L. Y., Jeng, J. E. M., Ho, S., Hsieh, M. Y., Lin, Z. Y. and Wang, L. Y. 2001. Betel quid chewing as a risk factor for hepatocellular carcinoma: a case-control study. Br. J. Cancer 84: 709-713.
4. Chang, L. Y., Wan, H. C., Lai, Y. L., Liu, T. Y. and Hung, S. L. 2006. Enhancing effects of areca nut extracts on the production of interleukin-6 and interleukin-8 by peripheral blood mononuclear cells. J. Periodontol. 77: 1969-1977.
5. Chang, M. C., Chiang, C. P., Lin, C. L., Lee, J. J., Hahn, L. J. and Jeng, J. H. 2005. Cell-mediated immunity and head and neck cancer: with special emphasis on betel quid chewing habit. Oral Oncol. 41: 757-775.
6. Rajendran, R., Sugathan, C. K., Remani, P., Ankathil, R. and Vijayakumar, T. 1986. Cell mediated and humoral immune responses in oral submucous fibrosis. Cancer 58: 2628-2631.
7. O’Garra, A. 1998. Cytokines induce the development of functionally heterogeneous T helper cell subsets. Immu-nity. 8: 275-283.
8. Billiau, A. and Matthys, P. 2009. Interferon-gamma: a historical perspective. Cytokine Growth Factor Rev. 20: 97-113.
9. Chiang, C. P., Wu, H. Y., Liu, B. Y., Wang, J. T. and Kuo, M. Y. 2002. Quantitative analysis of immunocompetent cells in oral submucous fibrosis in Taiwan. Oral Oncol. 38: 56-63.
10. Haque, M. F., Harris, M., Meghji, S. and Speight, P. M. 1997. An immunohistochemical study of oral submu-cous fibrosis. J. Oral Pathol. Med. 26: 75-82.
11. Haque, M. F., Meghji, S., Khitab, U. and Harris, M. 2000. Oral submucous fibrosis patients have altered levels of cytokine production. J. Oral Pathol. Med. 29: 123-128.
12. Hung, S. L., Cheng, Y. Y., Peng, J. L., Chang, L. Y., Liu,
T. Y. and Chen, Y. T. 2005. Inhibitory effects of areca nut extracts on phagocytosis of Actinobacillus actino-mycetemcomitans ATCC 33384 by neutrophils. J. Peri-odontol. 76: 373-379.
13. Chang, M. C., Wu, H. L., Lee, J. J., Lee, P. H., Chang, H. H., Hahn, L. J., Lin, B. R., Chen, Y. J. and Jeng, J. H. 2004. The induction of prostaglandin E2 production, interleukin-6 production, cell cycle arrest, and cytotox-icity in primary oral keratinocytes and KB cancer cells by areca nut ingredients is differentially regulated by MEK/ERK activation. J. Biol. Chem. 279: 50676-50683. 14. Wang, C. C., Liu, T. Y., Wey, S. P., Wang, F. I. and Jan,
T. R. 2007. Areca nut extract suppresses T-cell activation and interferon-gamma production via the induction of oxidative stress. Food Chem. Toxicol. 45: 1410-1418. 15. Chang, L. Y., Wan, H. C., Lai, Y. L., Kuo, Y. F., Liu, T.
Y., Chen, Y. T. and Hung, S. L. 2009. Areca nut extracts increased expression of inflammatory cytokines, tumor necrosis factor-alpha, interleukin-1beta, interleukin-6 and interleukin-8, in peripheral blood mononuclear cells. J. Periodontal. Res. 44: 175-183.
16. Wang, C. C., Lin, H. L., Wey, S. P. and Jan, T. R. 2011. Areca-nut extract modulates antigen-specific immunity and augments inflammation in ovalbumin-sensitized mice. Immunopharmacol. Immunotoxicol. 33: 315-322. 17. Liu, T. Y., Chen, C. L. and Chi, C. W. 1996. Oxidative
damage to DNA induced by areca nut extract. Mutat. Res. 367: 25-31.
18. Lee, J. H., Chang, S. H., Park, Y. S., Her, E., Lee, H. Y., Park, J. W., Han, J. W., Kim, Y. M. and Choi, W. S. 2004. In-vitro and in-vivo anti-allergic actions of Arecae semen. J. Pharm. Pharmacol. 56: 927-933.
19. Mukherjee, A. and Giri, A. K. 1991. Sister chromatid exchange induced by pan masala’(a betel quid ingredient) in male mice in vivo. Food Chem. Toxicol. 29: 401-403. 20. Schroder, K., Hertzog, P. J., Ravasi, T. and Hume, D. A.
2004. Interferon-gamma: an overview of signals, mech-anisms and functions. J. Leukoc. Biol. 75: 163-189. 21. Street, S. E. A., Trapani, J. A., MacGregor, D. and
Smyth, M. J. 2002. Suppression of lymphoma and epithelial malignancies effected by interferon γ. J. Exp. Med. 196: 129-134.
22. Janeway, C. Travers, A., P., Walport, M. and Capra, J. D. 1996. Immunobiology: the immune system in health and disease. 6th ed. pp.26. Garland Pub. New York, U.S.A. 23. Haque, M. F., Harris, M., Meghji, S. and Barrett, A.
W. 1998. Immunolocalization of cytokines and growth factors in oral submucous fibrosis. Cytokine 10: 713-719. 24. Pillai, M. R., Balaram, P., Kannan, S., Sudha, L., Nalina-kumari, K. R., Hareendran, N. K. and Nair, M. K. 1990. Interferon activation of latent natural killer cells and alteration in kinetics of target cell lysis: clinical implica-tions for oral precancerous lesions. Oral Surg. Oral Med. Oral Pathol. 70: 458-461.
25. Heimdal, J. H., Aarstad, H. J., Klementsen, B. and Olofsson, J. 1998. Disease stage related in vitro respon-siveness of peripheral blood T-lymphocytes in patients
with head and neck carcinoma. Acta. Otolaryngol. 118: 887-891.
26. Kim, R., Emi, M., Tanabe, K. and Arihiro, K. 2006. Tumor-driven evolution of immunosuppressive networks during malignant progression. Cancer Res. 66: 5527-5536.
27. Balaram, P., Pillai, M. R. and Abraham, T. 1987. Immu-nology of premalignant and malignant conditions of the oral cavity. II. Circulating immune complexes. J. Oral Pathol. 16: 389-391.
28. Chiang, C. P., Hsieh, R. P., Chen, T. H., Chang, Y. F., Liu, B. Y., Wang, J. T., Sun, A. and Kuo, M. Y. 2002. High incidence of autoantibodies in Taiwanese patients with oral submucous fibrosis. J. Oral Pathol. Med. 31: 402-409.
29. Ranadive, K. J., Gothoskar, S. V., Rao, A. R., Tezab-walla, B. U. and Ambaye, R. Y. 1976. Experimental studies on betel nut and tobacco carcinogenicity. Int. J. Cancer 17: 469-476.
30. Dasgupta, R., Saha, I., Pal, S., Bhattacharyya, A., Sa, G., Nag, T. C., Das, T. and Maiti, B. R. 2006. Immunosup-pression, hepatotoxicity and depression of antioxidant status by arecoline in albino mice. Toxicology. 227: 94-104.
31. Selvan, R. S., Selvakumaran, M. and Rao, A. R. 1991. Influence of arecoline on immune system: II. Suppres-sion of thymus-dependent immune responses and param-eter of non-specific resistance after short-term exposure. Immunopharmacol. Immunotoxicol. 13: 281-309. 32. Williamson, G. and Manach, C. 2005. Bioavailability
and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 81: 243S-255S. 33. Miyake, M., Ide, K., Sasaki, K., Matsukura, Y., Shijima,
K. and Fujiwara, D. 2008. Oral administration of highly oligomeric procyanidins of Jatoba reduces the severity of collagen-induced arthritis. Biosci. Biotechnol. Biochem. 72: 1781-1788.
34. Miyake, M., Sasaki, K., Ide, K., Matsukura, Y., Shijima, K. and Fujiwara, D. 2006. Highly oligomeric procyani-dins ameliorate experimental autoimmune encephalo-myelitis via suppression of Th1 immunity. J. Immunol. 176: 5797-5804.