臺灣山蘇花植株大小與孕性之關係
黃曜謀
1周雪美
2劉晉榮
3邱文良
1* 1 農委會林業試驗所森林生物組 2 元培科技大學生物技術學系 3 台北教育大學環境教育研究所 (收稿日期:2008.3.20,接受日期:2008.5.7) 摘 要 本研究針對臺灣北部烏來地區臺灣山蘇花,分析植株成熟度(孕性)與植株大小之相關性,評量 植株大小所引用的介量包括:叢徑大小、植株寬度、葉片數量、葉片長度及寬度。植株大小所引用 的介量彼此之間均呈直線正相關,其中以植株寬度與葉片長度之相關係數最高。當叢徑<12 cm 時、 植株寬度<79 cm、植株葉片數<9 片、平均葉長<37.8 cm 或平均葉寬<6.8 cm 時,植株不具孕性葉; 反之,當叢徑>15 cm 時、植株寬度>85 cm、植株葉片數>19 片、平均葉長>41.6 cm 或平均葉寬>7.8 cm 時,植株均具孕性葉。成熟植株與非成熟植株之間的植株大小介量均存一過渡帶。植株之孕性 葉比例(孕性葉片數/全部葉片數比例)與各種形態介量呈 S 型正相關,其中又以叢徑大小為推估台灣 山蘇花植株之孕性葉比例的最佳指標。 關鍵詞:臺灣山蘇花、孕性、物候學、蕨類植物緒 言
植物生長之孕性通常受到週遭環境變化所 調控,溫度、基質養分、水分、光線強度、日照 長度、樹冠遮蔽度等環境因子都會影響孕性葉的 形成(Wardlaw and Sharma, 1963; Conway, 1957; Dring, 1965; Sharpe, 1997; Odland, 1998, 2007)。 除外在環境因子外,植物之孕性亦同樣受到植株 內在因素所調控(Wardlaw, 1962; Odland, 1995), 例如:蕨類植物在達到完全成熟年齡時,葉片生 理開始發生改變,誘發孢子分生組織運作,於是 產 生 具 孢 子 ( 囊 群 ) 的 孕 性 葉 (Bower, 1923; Wardlaw, 1962; von Aderkas and Green, 1986)。有 關植株大小與蕨類的孕性關係研究不多(Bazzar et al., 1987; Ying and Huang, 1995; Tsai, 2005; Odland et al., 2006),其中以葉子長度最常應用在 評估孕性的指標。Odland (1995)調查發現在挪威 西部地區的 Athyrium distentifolium 長度超過 10 cm 的葉片始有生產孢子的能力,而 Thelypteris limbosperma 和 Matteuccia struthiopteris 葉片長度 則需分別超過 40 cm 及 60 cm 才有生產孢子的能 力 ; Odland (1998) 更 進 一 步 指 出 Thelypteris limbosperma 及 Athyrium distentifolium 葉片長度 不僅與能否生產孢子有關,且葉片越大,孢子囊數也越多。樹蕨植株高度曾被應用為孕性指標特 徵,例如筆筒樹樹高超過 1.75 m 方達生產孢子之 階 段 (Ying and Huang, 1995) 。 Mehltreter and Palacios-Rios (2003) 發 現 Acrostichum danaeifolium 植株孕性除了與葉片長度有關之 外,存活葉片數亦為重要指標,唯有植株葉片數 超過 10 片(含)以上者方達孕性階段。另外,Tsai (2005)發現山蘇花(Asplenium antiquum)孕性植株 的叢徑大於 22 cm,並認為植株叢徑大小為植株 孕性的良好指標。 臺 灣 山 蘇 花 (Asplenium nidus) 廣 泛 分 布 於 亞、非洲之熱帶、亞熱帶地區,常見於臺灣全島 低海拔山區(Shieh et al., 1994),被集中大規模栽 培並作為切葉與盆花用途(Chyuan, 1999)。目前已 有許多農業試驗單位推薦簡易孢子繁殖方法,介 紹給有興趣的農友採行,能夠生產並供應栽培 者,以解決種苗不足的問題,進而減少過度野外 採集所造成原生地生態的破壞。孢子是台灣山蘇 花的主要繁殖體,為避免徒勞無功或非達到最佳 的收集狀態,應對孢子的生產條件有所知悉,Yeh (1987)曾深入研究環境因子(包括:光照強度、溫 度、養分供給)對孢子形成之影響,然而,植株大 小與成熟度的相關性仍未見報導,因此本研究將 探討臺灣山蘇花植株大小與植株孕性之關係,提
供收集此ㄧ經濟植物孢子時之參考依據。
材料與方法
研究地區位於台北縣烏來鄉四崁水地區為 亞熱帶二次演替闊葉樹森林(東經 121°33’20’’,北 緯 24°53’36’’,海拔高約 500 m),主要優勢樹種 包含相思樹(Acacia confusa)、臺灣桫欏(Alsophila spinulosa)、筆筒樹(Sphaeropteris lepifera);根據 離 研 究 區 最 近 的 中 央 氣 象 局 台 北 測 候 站 2001-2005 年氣象資料顯示,該地區氣溫變化平 緩,月平均氣溫從 16℃到 30℃;月平均降水量 從 36 mm 到 581 mm,主要降水季節集中在夏秋 兩季,1 至 10 月的月平均降水量超過 100 mm, 為重濕期,雖 11、12 月平均降水量較少(分別為 36、70 mm),但仍為潮濕期,無旱季出現。 作者們先前在研究區內觀察發現,就各單一 台灣山蘇花植株而言,孕性葉與非孕性葉的出 現,與四季的環境變化之間並無明顯相關性存 在,因此,本研究僅於 2006 年 4 月進行單次調 查。選擇測量的植株標準:(1)易於原地測量,植 株著生高度距離地面 2.1 m 之內;(2)適當涵蓋不 同大小的植株(叢徑<10 cm,8 株;叢徑 11-20 cm, 8 株;叢徑 21-30 cm,5 株;叢徑 31-40 cm,6 株;叢徑>40 cm,7 株);(3)植株健康,葉片無明 顯病蟲害。在所調查的 34 植株,分別有 5、25、 4 植株的宿主為相思樹、臺灣桫欏及筆筒樹。 植 株 相 對 大 小 分 別 以 下 列 五 項 介 量 表 示 之:(1)叢徑大小(Fig. 1)(Ellwood et al., 2002; Martin et al., 2004; Tsai, 2005);(2)植株寬度(Fig. 1) (Martin et al., 2004);(3)葉片數(葉片已完全展 開);(4)葉寬度(葉子最寬之處)及(5)長度(包含葉 柄及葉身長度),每一植株測量完整且最大之 5 片葉子。 依據 Aguraiuja (2001)之定義,成熟植株係指 具生產孢子(囊)能力之個體,非成熟植株則指未 達生產孢子(囊)能力之個體,本研究中則將未發 現有孕性葉存在之個體歸入此一類群。此外,由 於臺灣山蘇花不具明顯的二型葉(dimorphic),本 文以孕性葉(fertile)及非孕性葉(sterile)分別代表 具有或不具有孢子(囊群)的葉子。孕性葉發育初 期若受到生物或機械傷害,也可能使得孢子囊呈 現發育停滯的現象。為減免外力傷害所造成不正 常發育(Tsai, 2005),因此,本研究將受損之葉片 予以摒除不納入計算。 所有表示植株大小之形態特徵與植株孕性 圖一、臺灣山蘇花。A、植株寬度;B 叢徑。Figure 1. Asplenium nidus. A. Width of plants; B.
Rosette.
進行統計分析,檢視其相關性,繪圖及統計分析 以套裝軟體(SigmaPlot 2000 for Windows)執行。
結 果
植株形態介量 直線迴歸分析顯示台灣山蘇花植株的五種 形態介量彼此之間均呈正相關,其中以植株寬度 與其他四個形態介量之間的相關係數最高(平均 值 0.90),其它依序平均葉長(平均值 0.86)、葉寬 (平均值 0.85)、叢徑(平均值 0.85),而以植株葉片 數與其他四個形態介量之間的相關係數最低(平 均值 0.76)。各形態介量之間,以植株寬與平均葉 長的相關係數最高(0.95),叢徑與植株葉片數之相 關係數最低(0.71)(Table 1)。 成熟植株與未成熟植株 調查 34 植株中,分別有 11 株及 23 株為未 成熟及成熟植株,未成熟植株形態介量平均值小 於成熟植株,且達極顯著標準(P<0.01)。叢徑小 於 12 cm 者均為未成熟植株,叢徑大於 15 cm 者 全為成熟植株,叢徑介於 12-15 cm 之間則同時存 在未成熟及成熟植株,視為「植株成熟過渡期」, 未成熟植株與成熟植株之平均叢徑分別為 7.7± 4.0 cm 及 32.6 ± 12.2 cm。植株寬度小於 79 cm 者 均為未成熟植株,植株寬度大於 85 cm 者全為成 熟植株,植株寬度介於 79-85 cm 之間則為植株成 熟過渡期,未成熟植株與成熟植株之平均植株寬 度分別為 44.5± 15.7 cm 及 155.8 ± 49.6 cm。植株 葉片數少於 9 片者均為未成熟植株,超過 19 片A
B
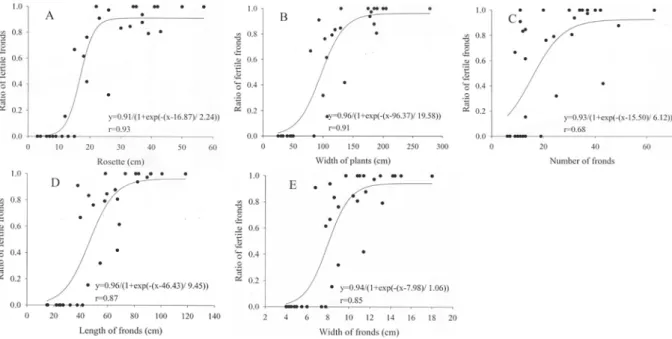
者全為成熟植株,植株葉片數介於 9-19 片之間則 為植株成熟過渡期,未成熟植株與成熟植株之平 均植株葉片數分別為 10.5 ± 3.7 片及 28.0 ± 14.3 片。平均葉長小於 37.8 cm 者均為未成熟植株, 平均葉長大於 41.6 cm 者全為成熟植株,平均葉 長介於 37.8-41.6 cm 之間則為植株成熟過渡期, 未成熟植株與成熟植株之平均葉長分別為 26.3 ± 8.2 cm 及 70.0 ± 20.7 cm。平均葉寬小於 6.8 cm 者均為未成熟植株,平均葉寬大於 7.8 cm 者全為 成熟植株,平均葉寬介於 6.8-7.8 cm 之間則為植 株成熟過渡期,未成熟植株與成熟植株之平均葉 寬分別為 5.4 ± 1.3 cm 及 11.0 ± 2.7 cm (Table 2)。 孕性葉與非孕性葉 調查 34 植株共計有 158 片完整葉子,其中 包括 62 片非孕性葉及 96 片孕性葉,非孕性葉之 葉長及葉寬小於孕性葉,且達極顯著標準。葉長 小於 36 cm 者均為非孕性葉,葉長大於 55 cm 者 全為孕性葉,葉長介於 36-55 cm 之間則同時有非 孕性葉及孕性葉,視為「葉片孕性過渡期」,非 孕性葉及孕性葉之平均葉長分別為 31.1 ± 11.6 cm 及 74.6 ± 20.4 cm。葉寬小於 6 cm 者均為非孕 性葉,葉寬大於 11 cm 者全為孕性葉,葉寬介於 6-11 cm 之間則為葉片孕性過渡期,非孕性葉及 孕性葉之平均葉寬分別為 6.3 ± 2.1 cm 及 11.5 ± 2.9 cm。然而,在葉片長寬比值(葉長/葉寬),非 孕性葉雖小於孕性葉(分別為 5.1± 1.5 與 6.7 ± 1.7),但未達顯著標準(Table 3)。 本研究以孕性葉片數佔全部成熟葉片數的 比例表示該植株的孕性葉比例,其數值範圍為 0.00-1.00,檢視植株形態介量與植株孕性葉比例 之關係。根據非直線迴歸分析,叢徑大小與植株 孕 性 葉 比 例 具 S 型 正 相 關 (y=0.91/(1+exp(-(x- 16.87)/ 2.24) ); r=0.93),且預測值與觀測值之間其 相關係數是所有形態介量中最高的,當叢徑小於 10 cm 以下,孕性葉比例預測值低於 0.05,當叢 徑超過 10 cm,孕性葉比例急劇上升,直至數值 約 0.90 始趨於穩定(叢徑大於 25 cm) (Fig. 2A)。 同樣地,植株寬度與植株孕性葉比例也具 S 型正 相關(y=0.96/(1+exp(-(x-96.37)/ 19.58)); r=0.91), 植株寬度小於 40 cm 以下,孕性葉比例預測值低 於 0.05,當植株寬度逐漸增大,孕性葉比例急劇 表一、台灣山蘇花植株各形態介量間直線迴歸相關係數(r 值)。
Table 1. Linear correlation coefficients (r) among five parameters of Asplenium nidus.
R W N AL AW
Rosette (R) --
Width of plants (W) 0.93** --
Number of fronds (N) 0.71** 0.81** --
Average length of fronds (AL) 0.89** 0.95** 0.73** --
Average width of fronds (AW) 0.85** 0.90** 0.79** 0.85** --
t-test, ** indicates P<0.01
表二、未成熟植株(11 株)與成熟植株(23 株)形態介量之比較(單位:cm)
Table 2. Comparison of morphological parameters of the immature (11 plants) and the mature plants (23 plants) of Asplenium nidus (unit: cm).
Immature plants Mature plants
Rosette 7.7 ± 4.0 (3-15) 32.6 ± 12.2 (12-57) Width of plants 44.5 ± 15.7 (25-85) 155.8 ± 49.6 (79-279) Number of fronds 10.5 ± 3.7 (6-19) 28.0 ± 14.3 (9-63) Average length of fronds 26.3 ± 8.2 (15-41.6) 70.0 ± 20.7 (37.8-118.4) Average width of fronds 5.4 ± 1.3 (4-7.8) 11.0 ± 2.7 (6.8-18.0) 表三、非孕性葉(62 片葉)與孕性葉(96 片葉)形態介量之比較。
Table 3. Comparison of morphological parameters of the sterile fronds (62 fronds) and the fertile fronds (96 fronds) of Asplenium nidus.
Sterile fronds Fertile fronds Average fronds length (cm) 31.1 ± 11.6 (11-55) 74.6 ± 20.4 (36-128) Average fronds width (cm) 6.3 ± 2.1 (3-11) 11.5 ± 2.9 (6-19) Frond length / frond width 5.1± 1.5 (2.1-10.3) 6.7 ± 1.7 (3.7-12.9)
圖二、台灣山蘇花五種形態介量與植株孢子葉比例之關係。A、叢徑;B、植株寬度;C、植株葉片數;D、平 均葉長;E、平均葉寬。
Figure 2. The correlation between fertile frond ratio and five parameters of Asplenium nidus. A. Rosette; B. Width of
plant; C. Number of fronds; D. Average length of fronds; E. Average width of fronds. 上升,直至植株寬度達 155 cm 以上,孕性葉比 例始趨於穩定(數值約 0.95) (Fig. 2B)。植株葉片 數少於 9 片者均為未成熟植株,當植株葉片數超 過 9 片,孕性葉比例急劇上升(y=0.93/(1+exp(-(x- 15.50)/ 6.12)), r=0.68),直至數值約 0.9(植株葉片 數大於 25 片)始趨於穩定(Fig. 2C),不過預測值 與觀測值之間其相關係數是所有形態介量中最 低的。平均葉長小於 37.8 cm 者均為未成熟植 株,當植株平均葉超過 37.8 cm,孕性葉比例同 樣 呈 急 劇 上 升 (y=0.96/(1+exp(-(x-46.43)/ 9.45)), r=0.87),直至數值約 0.95(植株平均葉長大於 80 cm)始趨於穩定(Fig. 2D)。平均葉寬與植株孕性葉 比例具 S 型正相關(y=0.94/(1+exp(-(x-7.98)/ 1.06)), r=0.85),植株平均葉寬小於 5 cm 以下,孕性葉 比例預測值低於 0.05,當植株平均葉寬逐漸增 大,孕性葉比例呈急劇上升,直至植株平均葉寬 達 12 cm 以上,孕性葉比例始趨於穩定(數值約 0.95) (Fig. 2E)。
討 論
台灣山蘇花從幼孢子體第一片葉開始至成熟孕 性葉,均為單葉,葉型幾乎無變異,因此須尋求 除葉型以外,更多判斷年齡或成熟度的形態特 徵。本研究參考 Tsai (2005)測量山蘇花大小主要 形態介量叢徑之外,另外再加入植株寬度、植株 葉片數、葉長及葉寬等四種介量。結果顯示:用 來描述台灣山蘇花植株大小的五種形態介量彼 此之間均呈直線正相關,其中以植株寬度與其他 四 個 形 態 介 量 之 間 的 相 關 係 數 最 高 ( 平 均 值 0.90),即使葉片數與其他四個形態介量之間的相 關係數最低,其平均值也有 0.76。若將台灣山蘇 花各種介量間兩兩作直線迴歸分析,植株寬與平 均葉長的相關係數最高(0.95),植株寬約為平均葉 長的四倍。台灣東北部福山地區山蘇花叢徑大小 與植株葉片數之間有正相關,其相關係數值偏低 (r=0.4),Tsai (2005)認為係取樣數量過少(20 株) 所造成樣本值的變異較大;除了取樣偏低之外, 光 線 的 多 寡 也 可 能 是 其 中 之 ㄧ , 如 Cyathea caracasana,在開闊棲地植株葉片生產量約為森 林裡植株的 2~3 倍(Arens, 2001)。或許是本研究 之生育地環境(包括光線強度)較福山地區來的均 勻,使得台灣山蘇花叢徑大小與植株葉片數之相 關係數(r=0.76)較福山地區山蘇花為高;當然,並 不排除種間差異的可能性。 蕨類植物到達一定的年齡後便有產生孕性 葉的能力(Bower, 1923; Wardlaw, 1962),一般蕨 類需要 1 至 5 年時間方達到成熟階段(White, 1971)。蕨類植物年齡或成熟度通常可由外部形態 或/及植株大小來加以判識,例如: Platyceriumbifurcatum 年齡可由植株葉片種類來加以推斷, 幼株長出片狀葉,兩年後開始產生基部葉,唯有 年 齡 超 過 3-4 年 才 生 成 孕 性 葉 (Pemberton, 2003)。Lomagramma guianensis 幼株與成熟植株 亦可由葉片形態來鑑別,幼株葉片一回羽裂,成 熟植株則生產一回羽狀複葉(Gay, 1993)。墨西哥 灣 Acrostichum danaeifolium 的成熟植株需達到 10 片 葉 片 以 上 (Mehltreter and Palacios-Rios, 2003)。Thelypteris angustifolia 植株平均葉片數 3 (最小值-最大值:1-8),成熟(具孕性葉)植株平均 葉片數(4.1)較未成熟(無孕性葉)植株(2.8)來得多 (Sharpe, 1997)。樹蕨類以樹高當作成熟指標是極 為普遍的作法,例如:在 Fiji 的熱帶性樹蕨 Cyathea hornei 植株在 1 m 以上與葉片長度在 90 cm 以上才有孕性葉出現(Ash, 1987);筆筒樹的樹 高大於 1.75 m 後具有產生孕性葉的能力(Ying and Huang, 1995);上述之蕨類是否達成熟階段, 係由少數形態特徵作二分法切割,然而本研究結 果顯示成熟植株與非成熟植株之間的植株大小 介量均存一過渡帶;Polypodium scouleri 也有類 似的現象,當植株重量介於 0.5- 5.0 kg、葉片長 度 30 cm- 70 cm 或羽片數 11-23 片時,均為此種 蕨類成熟植株與非成熟植株之間的過渡帶(Sillett and Bailey, 2003)。 臺灣山蘇花之孕性葉比例範圍從 0.00 至 1.00,比例愈高代表孕性葉片出現頻率愈高,孕 性葉比例與所分析的五個形態介量均呈 S 型正相 關,換言之,隨植株長大成熟長出孕性葉後,孕 性葉比例會隨之急速增加,高至一比例後始趨於 固定值(0.90-0.95),其中又以叢徑大小之迴歸預 測值與觀測值最為高度相關(r=0.93),其它依序為 植株寬度(r=0.91)、平均葉長(r=0.87)、植株平均 葉寬(r=0.85)、植株葉片數(r=0.68)。台灣山蘇花 叢徑較大者,植株中絕大多數的葉片為孕性葉, 但仍舊有極少數葉片以非孕性葉的形式存在,可 能與外界物理傷害或光照強度有關;當山蘇花之 孕性葉在萌芽期如受到枯枝擊損機械性損傷,會 喪 失 本 來應有 的 繁 殖能力 轉 換 成營養 葉 (Tsai, 2005),本研究已事先排除受損之葉片不計,因 此物理傷害應非限制孕性葉片形成的主因素;另 外,Harvey and Caponetti (1972, 1973)指出在樹冠 鬱閉遮蔽情況下,光照量不足導致光合作用下 降,當葉片內蔗糖濃度不足時,孢子囊發育及孢 子形成將嚴重受阻,無法形成孕性葉。Yeh and Wang (2000) 也 曾 指 出 密 葉 鐵 線 蕨 (Adiantum raddianum)於低光照下生長並無植株形成孢子, 隨著光照強度的增加,孕性葉片數隨之呈直線增 加。至於臺灣山蘇花成熟植株之非孕性葉的形 成,是否受光照量、被其它葉片壓迫、或尚有其 它原因所影響,仍有待進一步確認。 Sato (2005)曾採用十個形態介量評估日本地 區 Polystichum tripteron 發育階段,發現其中以葉 片中肋分叉數對孕性葉與非孕性葉的鑑別能力 最為有效,是一有效研究該物種生活史工具。本 研究結果顯示台灣山蘇花非孕性葉之葉長及葉 寬均小於孕性葉,但兩者之間並無明確的區分界 線,而是存有一過渡期,至於葉片長寬比值方 面,非孕性葉雖小於孕性葉,但未達顯著標準, 不適宜當作判斷葉片是否具孕性的指標。儘管, 本研究發現台灣山蘇花未成熟植株的五種形態 介量平均值小於成熟植株者,且達極顯著標準, 不 過 成 熟 植 株 與 未 成 熟 植 株 之 間 存 著 一 過 渡 期,因此直接由形態介量判斷植株是否達到成熟 階段須格外謹慎。另外,台灣山蘇花植株孕性葉 比例與叢徑大小、植株寬度、葉片數、葉寬度及 葉長度均呈 S 型極顯著相關,其中以叢徑大小為 介量之迴歸式預測值與觀測值相關係數最高。
參考文獻
Aguraiuja R. 2001. Population status of five Hawaiian endemic fern taxa within the genus Diellia (Aspleniaceae). CBM:s Skriftserie 3: 7-24.
Arens NC. 2001. Variation in performance of the tree fern Cyathea caracasana (Cyatheaceae) across a successional mosaic in an Andean cloud forest. Amer. J. Bot. 88: 545-551. Ash J. 1987. Demography of Cyathea hornei
(Cyatheaceae), a tropical tree-fern in Fiji. Aust. J. Bot. 35: 331-342.
Bazzar FA, Chiarello NR, Coley PD, and Pitelka LF. 1987. Allocating resources to reproduction and defence. BioScience 37: 58-67.
Bower FO. 1923. The ferns. Volume I. Cambridge University Press, Cambridge.
Chyuan JH. 1999. In-vitro culture of leaf primordium of Asplenium nidus. Bull. Hualien Dist. Agri. Impro. Stat. 17: 53-62. (in Chinese)
Conway E. 1957. Spore production in bracken. J. Ecol. 45: 273-284.
Dring MJ. 1965. The influence of shaded conditions on the fertility of bracken. Brit. Fern Gaz. 9:
222-227.
Ellwood MD, Jones DT, and Foster WA. 2002. Canopy ferns in lowland dipterocarp forest support a prolific abundance of ants, termites, and other invertebrates. Biotropica 34(4): 575-583.
Gay H. 1993. The architecture of a dimorphic clonal fern, Lomagramma guianensis (Aublet) Ching (Dryopteridaceae). Bot. J. Linn. Soc. 111: 343-358.
Harvey WH, and Caponetti JD. 1972. In vitro studies on the induction of sporogenous tissue on leaves of cinnamon fern. I. Environmental factors. Can. J. Bot. 50: 2673-2682.
Harvey WH, and Caponetti JD. 1973. In vitro studies on the induction of sporogenous tissue on leaves of cinnamon fern. II. Some aspects of carbohydrate metabolism. Can. J. Bot. 51: 341-349.
Martin CE, Lin TC, Hsu CC, Lin SH, Lin KC, Hsia YJ, and Chiou WL. 2004. Ecophysiology and Plant Size in a Tropical Epiphytic Fern, Asplenium nidus, in Taiwan. Intel. J. Plant Sci. 165: 65–72.
Mehltreter K. and Palacios-Rios M. 2003. Phenological studies of Acrostichum danaeifolium (Pteridaceae, Pteridophyta) at a mangrove site on the Gulf of Mexico. J. Trop. Ecol. 19: 155-162.
Odland A. 1995. Frond development and phenology
of Thelypteris limbosperma, Athyrium
distentifolium, and Matteuccia struthiopteris in western Norway. Nord. J. Bot. 15: 225-236. Odland A. 1998. Size and reproduction of
Thelypteris limbosperma and Athyrium distentifolium along environmental gradients in western Norway. Nord. J. Bot. 18: 311-321. Odland A. 2007. Geographical variation in frond size, rootstock density, and sexual reproduce- tion in Matteuccia struthiopteris populations in Norway. Popul. Ecol. 49: 229-240.
Odland A, Naujalis JR, and Stapulionyte. 2006. Variation in the structure of Metteuccia struthiopteris populations in Lithuania. Biologia 1: 83-90.
Pemberton RW. 2003. The common staghorn fern, Platycerium bifurcatum, naturalizes in southern Florida. Amer. Fern J. 93: 203-206. Sato T. 2005. Quantitative expression of fern leaf
development and fertility in Polystichum tripteron (Aspidiaceae). Plant Syst. Ecol. 150:
191-200.
Sharpe JM. 1997. Leaf growth and demography of the rheophytic fern Thelypteris angustifolia ( Willdenow ) Proctor in a Puerto Rican rainforest. Plant Ecol. 130: 203-212.
Shieh WC, DeVol CE, and Kuo CM. 1994. Aspleniaceae. In: Editorial Committee of the Flora of Taiwan (eds.) Flora of Taiwan I (2nd edition), Sandos Chromograph Printing Company, Taipei, pp.449-464.
Sillett SC, and Bailey MG.. 2003. Effects of tree crown structure on biomass of the epiphytic fern Polypodium scouleri (Polypodiaceae) in redwood forests. Amer. J. Bot. 90: 255-261. Tsai PH. 2005. The phenology of Asplenium
antiquum Makino in the Fushan experimental forest. Master thesis, School of Forestry and Resource conservation, National Taiwan University, Taipei, Taiwan. (in Chinese).
von Aderkas P, and Green PEJ. 1986. Leaf development of the ostrich fern Metteuccia struthiopteris (L.) Todaro. Bot. J. Linn. Soc. 93: 307-321.
Wardlaw CW. 1962. The sporogenous meristems of ferns: A morphogenetic commentary. Phyto- morphology 12: 394-408.
Wardlaw CW, and Sharma DN. 1963. Experimental and analytical studies of pteridophytes. XL. Factors in the formation and distribution of sori in leptosporangiate ferns. Ann. Bot. 27: 101-121.
White RA. 1971. Experimental and developmental studies of the fern sporophyte. Bot. Rev. 37: 509-540.
Yeh DM. 1987. The growth habits of Nephrolepis exaltata and Asplenium nidus and effects of temperature, mineral nutrition and growing medium on growth. Mater thesis, Institute of Horticulture, National Taiwan University, Taipei, Taiwan. (in Chinese).
Yeh DM, and Wang HM. 2000. Effects of irradiance on growth net photosynthesis and indoor performance of the shade-adapted plant, maidenhair fern. J. Hort. Sci. Biotech. 75: 293-298. (in Chinese).
Ying SS, and Huang YM. 1995. Phenological study on Sphaeropteris lepifera at Su-Au area. Mem.Coll. Agri. Natl. Taiwan Univ. 35(4): 451-64. (in Chinese).
The Correlation of Plant Size and Fertility of Asplenium nidus L.
(Aspleniaceae; Pteridophyta)
Yao-Moan Huang
1, Hsueh-Mei Chou
2, Chin-Jung Liu
3, Wen-Liang Chiou
1*1
Division of Forest Biology, Taiwan Forestry Research Institute Taipei, Taiwan
2
Department of Biotechnology, Yuanpei University Shin-chu, Taiwan
3
Graduate School of Environmental Education, Taipei Municipal University of Education Taipei, Taiwan
(Received: 20 March 2008, accepted: 7 May 2008)
ABSTRACT
This study focuses on the correlation between plant size and fertility of Asplenium nidus sampled from Wulai, northern Taiwan. Morphological parameters used to assess plant size include rosette size, plant width, frond number, length and width of fronds. There are linear positive correlations between these five parameters. The correlation coefficient of plant width and frond length is the highest among them all. Plants with rosette size < 12cm, plant width < 79 cm, frond number < 9, average frond length < 37.8 cm, or average frond width < 6.8 cm do not produce fertile fronds. On the contrary, plants with rosette size > 15 cm, plant width > 85 cm, frond number > 19, average frond length > 41.6 cm, or average frond width > 7.8 cm produce fertile fronds. A transitional stage exists between all five morphological parameters of mature and immature plants. The ratio of fertile fronds (number of fertile fronds / number of all fronds) of each plant is S-formed positive correlation with its morphological parameters, with rosette size being the best indicator among them in estimating the ratio of fertile fronds.