The FASEB Journal
•
Research Communication
LPA
1
is essential for lymphatic vessel development
in zebrafish
Shyh-Jye Lee,*,†,1 Tun-Hao Chan,*,2 Tzu-Cheng Chen,*,2 Bo-Kai Liao,‡ Pung-Pung Hwang,‡ and Hsinyu Lee*,†,1
*Institute of Zoology and†Department of Life Science, National Taiwan University, Taipei, Taiwan; and‡Institute of Cellular and Organismic Biology, Academia Sinica, Taipei, Taiwan
ABSTRACT Lysophosphatidic acid (LPA) has long been implicated in regulating vascular development via endothelial cell-expressed G protein-coupled recep-tors. However, because of a lack of notable vascular defects reported in LPA receptor knockout mouse studies, the regulation of vasculature by LPA receptors
in vivo is still uncertain. Using zebrafish as a model, we
studied the gene expression patterns and functions of an LPA receptor, LPA1, during embryonic
develop-ment, in particular, vascular formation. Whole-mount
in situ hybridization experiments revealed that
ze-brafish lpa1(zlpa1) was ubiquitously expressed early in
development, and its expression domains were later localized to the head region and the vicinity of the dorsal aorta. The expression of zlpa1surrounding the
dorsal aorta suggests its role in vasculature develop-ment. Knocking down of zLPA1 by injecting
morpho-lino (MO) oligonucleotides at 0.625–1.25 ng per em-bryo resulted in the absence of thoracic duct and edema in pericardial sac and trunk in a dose-dependent manner. These zlpa1-MO-resulted defects could be
specifically rescued by ectopic expression of zlpa1. In
addition, overexpression of vegf-c, a well-known lym-phangiogenic factor, also partially ameliorated the in-hibition of thoracic duct development. Taken together, these results demonstrate that LPA1 is necessary for
lymphatic vessel formation during embryonic develop-ment in zebrafish.—Lee, S.-J., Chan, T.-H., Chen, T.-C., Liao, B.-O., Hwang, P.-P., Lee, H. LPA1is essential for
lymphatic vessel development in zebrafish. FASEB J. 22, 3706 –3715 (2008)
Key Words: lysophosphatidic acid䡠 G protein-coupled receptor 䡠 endothelial cell 䡠 thoracic duct 䡠 morpholino
A complex vasculature network exists for the ade-quate exchange of gases, nutrients, signal molecules, and cells circulating between tissues and organs in vertebrates. This highly branched vasculature is mainly composed of blood and lymphatic vascular systems. Both types of vessels are assembled by a lining of endothelial cells (ECs). Vasculogenesis begins in the early gastrulation stage when primary axial vessels are formed, followed by angiogenesis for branching into the entire blood-conveyance network (1). After
forma-tion of the blood network, the lymphatic system devel-ops blind-ended capillaries, which converge into a larger collecting duct (the thoracic duct), ultimately merging into the anterior venous (2). The formation of the entire vasculature network depends on the precise molecular control of proliferation, migration, and as-sembly of ECs, and also EC signaling for destabilizing and stabilizing the vasculature (1). Understanding en-dothelial signaling during development in both embry-onic and adult tissues is crucial in providing insights into the regulation of development and certain disease conditions such as cancer (3).
Lysophosphatidic acid (LPA), a bioactive lipid, has diverse cellular (4) and signaling functions via its cognate cell surface G protein-coupled receptors (5, 6), and it has been implicated as having an emerging role in a variety of cancers (4). LPA receptor 1 (LPA1) of the
endothelial differentiation gene (Edg) subfamily (7) was the first receptor identified for LPA (8). The lpa1
gene is ubiquitously expressed at high levels in the colon, small intestine, placenta, brain, and heart, and moderately expressed in the pancreas, ovaries, and prostate (8). Because of its high enrichment in the neurogenic ventricular zone, the lpa1 gene was
previ-ously called ventricular zone gene-1 (vzg-1), and research mainly focused on studying its role in brain neurogen-esis. A knockout mouse study initially failed to identify a role for LPA1in the developing brain (9). However, a
study using a variant of LPA1-null mice propagated
from the original LPA1 knockout mice, the so-called
maLPA1mice, did exhibit a reduced ventricular zone,
altered neuronal markers, and increased cell death (10). Recently, we have demonstrated that LPA1 is
highly expressed in human umbilical vascular endothe-lial cells (HUVECs) (11). The high expression of LPA1
in ECs is very intriguing, as LPA has been suggested to play an important role in angiogenesis (12). In addi-tion, the LPA1 was demonstrated to link pulmonary
fibrosis to lung injury by regulating fibroblast
recruit-1Correspondence: 1 Roosevelt Rd., Section 4, Institute of
Zoology, National Taiwan University, Taipei, Taiwan 106, R.O.C. E-mail: S.-J.L., [email protected]; H.L., hsinyulee@ ntu.edu.tw
2These authors contributed equally to this work.
ment and vascular leak (13). LPA can regulate human EC proliferation (14, 15), migration (14, 15), capillary-like tube formation in vitro (16), activation of proteases (17), expression of inflammation-related genes (18), and endothelial permeability (19). It also mediates contractility, proliferation, and differentiation of vascu-lar smooth muscle cells (20 –24). Unsaturated LPA has been shown to trigger rat blood vessel remodeling in vivo (25). Moreover, knocking out a synthesizing en-zyme of LPA, autotaxin, causes lethality in mice on embryonic day 9.5, with profound vascular defects in the yolk sac (26). These results suggest that LPA is essential for blood vessel formation during develop-ment. However, none of the LPA receptor knockout studies, including those using LPA1(9), LPA2(27), and
LPA3 (28) null-mice, reported defects in the
vascula-ture except for a low incidence of cranial hemorrhage in LPA1 and LPA1/LPA2 null mice (9, 27). Whether
other recently identified LPA receptors, such as LPA4
(29) and LPA5(30), or complementary effects between
LPA receptors, might have caused the lack of detect-able vascular defects in those LPA receptor knockout mice is unclear. Thus, the role of LPA receptors in vascular development has remained an unresolved is-sue.
To study LPA receptor function, especially its role in vascular formation, we identified an lpa1 gene in ze-brafish, which is a well-established model for angiogen-esis research (31, 32). We demonstrate that knocking down LPA1translation results in inhibition of thoracic
duct formation. In addition, these defects could be fully rescued by the ectopic expression of lpa1and partially ameliorated by a lymphangiogenic factor, vegf-c. Taken together, these results suggest that LPA1functions as a
critical regulator directing lymphatic vessel develop-ment.
MATERIALS AND METHODS
Maintenance of zebrafish
Breeding wild-type (AB) and transgenic zebrafish (Tg(fli1:
EGFP)y1 from the Zebrafish Information Research Center
(Eugene, OR, USA) were cultured at 27–28°C on a 14-h light/10-h dark cycle. Embryos were collected by natural spawning, raised in 0.3⫻ Danieau’s buffer [by diluting 1⫻ Danieau’s buffer: 58 mM NaCl, 0.7 mM KCl, 0.4 mM MgSO4,
0.6 mM Ca(NO3)2, and 5.0 mM HEPES (pH 7.6), with
double-distilled water] supplemented with 50g/ml strepto-mycin and 50g/ml penicillin G at 28.5°C until observation or fixation. Embryos were staged according to Kimmel et al. (33), and stages are given as hours or days postfertilization (hpf or dpf, respectively). All animal handling procedures followed the guidelines for the use of laboratory animals at National Taiwan University, Taipei, Taiwan.
Cloning and analysis of the zebrafish lpa1gene
A reverse-transcription polymerase chain reaction (RT-PCR) was performed on total RNAs extracted from embryos at the designated times using the TRIzol reagent (Invitrogen,
Carls-bad, CA, USA), according to the manufacturer’s instructions. RNAs were subsequently treated with DNase I (Invitrogen), and first-strand complementary (c) DNAs were synthesized. cDNAs were generated from total RNAs of 24-hpf zebrafish larvae by reverse transcription using the M-MLV reverse transcription kit (Promega, Madison, WI, USA). A 1208-base pair (bp) DNA fragment containing the coding region of the zebrafish lpa1 gene (zlpa1, NM_001004502) was amplified
from zebrafish embryo cDNAs by PCR using primers with the following sequences: 5⬘-GCGAGTGATTTCTGGACCTT-TCAGC-3⬘ (forward) and 5⬘-GCCTTTTTCACAGTCTCTTCT-TGCG-3⬘ (reverse). The amplified PCR fragment was sub-cloned into a pGEM-T easy vector (Promega) for sequence verification. Amino acid sequences of LPA1 from different
species among chordates were identified from the National Center for Biotechnology Information (NCBI) database and aligned using the Vector NTI software (Invitrogen). The phylogenetic tree was constructed and drawn using the MEGA3 software (Biodesign Institute, Tempe, AZ, USA).
RT-PCR analysis
RNAs and cDNAs from embryos at designated stages and from different adult tissues were prepared as described previously. A 136-bp zlpa1 fragment was amplified with the
following primers: 5⬘-CGGAGGGTAGTTGTGGTTATAG-3⬘ (forward) and 5⬘-GGTACGAGTTGCTGTAGAGTG-3⬘ (re-verse). Amplification of a 542-bp ef1␣ fragment served as the RT-PCR internal control using the following primers: 5 ⬘-CAAGGAAGTCAGCGCATACA-3⬘ (forward) and 5⬘-TGAT-GACCTGAGCGTTGAAG-3⬘ (reverse).
Whole-mount in situ hybridization (WISH)
Embryos were grown to desired stages, fixed in 4% parafor-maldehyde in phosphate-buffered saline (PBS) overnight, and manually dechorionated using fine forceps. Embryos were then stored in 100% methanol at ⫺20°C until use. Antisense digoxigenin (DIG) -labeled RNA riboprobes were synthesized according to the manufacturer’s instructions (Roche Applied Science, Penzberg, Germany). Hybridization and detection with an alkaline phosphatase-coupled anti-DIG antibody (Roche Applied Science) were performed according to Thisse et al. (34). Photographs were taken with a Nikon Coolpix995 digital camera (Nikon, Tokyo, Japan). The image background was adjusted to the white point by the Curves tool of Adobe Photoshop (Adobe Systems, San Jose, CA, USA), and all channel output levels were set to 228. For cryosections, stained samples were immersed in PBS containing 30% sucrose overnight, and embedded in optimal cutting temper-ature (OCT) compound embedding medium (Sakura, To-kyo, Japan) at⫺20°C, and 10-m frozen cross-sections were cut with a CM 1900 rapid sectioning cryostat (Leica, Heidel-berg, Germany) and attached to poly-l-lysine-coated slides (Electron Microscopy Sciences, Ft. Washington, PA, USA). Bright-field images were acquired with a TCS-SP5 confocal laser scanning microscope with a Leica HCX PL-APO⫻63/ 1.30 glycerol objective (Leica Lasertechnik, Heidelberg, Ger-many) using a laser wavelength at 405 nm.
Morpholino oligonucleotides
Antisense morpholinos (MOs) were purchased from Open Biosystems (Huntsville, AL, USA) or custom made by Gene Tools (Philomath, OR, USA). To knockdown zlpa1 gene
activity, we used two nonoverlapping MOs targeting the boundaries flanking exon 2 to interfere with its splicing. The first splice-blocking MO was designated sMO1 (sequence:
5⬘-TGGAGCACTTACCCAATACAATCAC-3⬘), which targets the boundary between exon 2 and intron 2 (⫺13 to ⫹12). The second splice-blocking MO was designated sMO2
(se-quence: 5⬘-GGTCTGTTTCTGAAAAGTAAAGATA-3⬘), which targets the boundary between intron 1 and exon 2 (⫺16 to ⫹9). A 5-bp mismatched splice-blocking MO of sMO1 was
generated to serve as a control with the following sequence, with the mismatched pairs in lowercase letters: 5⬘-TGcAG-gACTTACCgAATAgAATgAC-3⬘. In addition, a reported ze-brafish vegf-c (zvegf-c) MO (35) (with the sequence: 5⬘-GAAAATCAAATAAGTGCATTTTAG-3⬘) was purchased from GeneTools. The MOs were dissolved in sterile double-distilled water to 1 mM, stored at⫺20°C, and further diluted to the desired working concentrations in 1⫻ Danieau’s buffer with 0.5% phenol red and kept at 4°C before being used.
Expression vector construction
The cDNAs of coding regions of zlpa1 were amplified by
RT-PCR and TA-cloned into the pcDNA3.1/V5-His-TOPO vector designed to express proteins with V5 and 6-histidine double fusions at the C-terminal under the CMV promoter (Invitrogen). The primers used were as follows: 5⬘-ATTGCG-GCCGCCATGGATGATAGACAATGCTA-3⬘ (zlpa1, forward),
5⬘-ATTCTCGAGACCACTGAATGGTCATTATGGTGA-3⬘ (zlpa1, reverse). The zvegf-c in the same expression vector as zlpa1was kindly provided by C. C. Huang (Institute of Cellular
and Organismic Biology, Academia Sinica, Taipei, Taiwan).
Microinjection procedures
Microinjections were carried out according to Lai et al. (36). Briefly, embryos at desired stages were immobilized in an injection trough on a 90-mm 2% agar plate. MOs or expres-sion vectors were prepared as described at designated con-centrations. An injection volume from 0.5–2.3 nl of different MOs, vectors, or vehicles was injected into 1-cell-stage em-bryos. After injection, embryos were recovered from the injection troughs and cultured in 0.3⫻ Danieau’s buffer until being examined.
Photographs of vascular and lymphatic vessels
Blood or lymphatic vessels of Tg(fli1:EGFP)y1 zebrafish were
observed under a DM5000B epifluorescent microscope (Leica, Wetzlar, Germany) with Leica HC PL Fluotar objectives using a fluorescein isothiocyanate (FITC) or rhodamine filter cube and imaged using a CoolSNAPfx CCD camera (Roper Scientific,
Tucson, AZ, USA). The images were acquired using the Sim-plePCI program (ver. 5.2; Compix, Cranberry Township, PA, USA) and pseudocolored using green or red color for the FITC or rhodamine images, respectively.
Statistical analysis
All experimental values are presented as mean⫾ sd and were analyzed by paired-sample Student’s t test in Microsoft Excel (Microsoft, Redmond, WA, USA).
RESULTS
Cloning, sequencing, and expression profile analysis of lpa1
To investigate the roles of LPA1during embryonic
devel-opment, we first identified an lpa1gene (NM_001004502)
from the NCBI database and isolated a 1208-bp DNA fragment by PCR amplification from zebrafish em-bryo cDNAs. This DNA fragment was subcloned, sequenced, and found to be 100% matched to the reported lpa1sequence from 141 to 1348 nucleotides of NM_001004502. It is referred as zlpa1hereafter. The gene, zlpa1, encodes a protein with 346 amino acids and a predicted molecular weight of 39.6 kDa. We aligned the translated amino acid sequences of zebrafish, mouse (NP_034466), and human (NP_001392) LPA1,
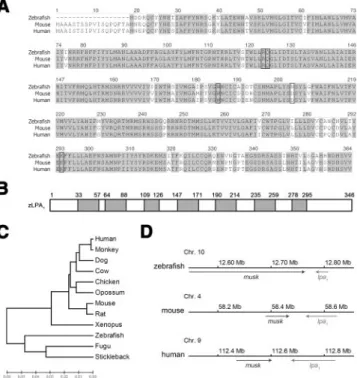
and we found that they were highly homologous with a sequence identity of 85% (Fig. 1A). A phylogenic analysis showed that zebrafish LPA1 is most closely
related to the LPA1of Fugu (Takifugu rubripes) among
chordates (Fig. 1B). To understand the chromosomal location of lpa1, we performed a syntenic analysis of these lpa1genes. The lpa1 gene is located at chromo-somes 10, 4, and 9 in the zebrafish, mouse, and human, respectively. They all have the same orientation and are positioned next to musk, a muscle-specific kinase receptor gene (Fig. 1C). Collectively, these results suggest that zlpa1 is evolutionarily related to mammalian lpa1genes.
Figure 1. Sequence analysis of the zebrafish LPA receptor, zLPA1. A) Amino acid sequence alignment of zLPA1 with its
human and mouse homologs. Identical amino acids are shaded in gray. The putative LPA binding sites are enclosed by rectan-gles. B) The zLPA1amino acid sequence was analyzed by the
HMMTOP 2.0 (http://www.enzim.hu/hmmtop/index.html). Seven predicted transmembrane domains are shown in gray boxes flanking boundary amino acid numberings. C) The phylogenic tree of LPA1homologs in chordates. The horizontal
length is proportional to the estimated time from divergence of the gene from the related family member. D) Chromosomal location of the zebrafish, mouse and human lpa1genes. The lpa1
gene is located at chromosome (Chr.) 10, 4, and 9 in the zebrafish, mouse, and human, respectively. They are all in the same orientation (left arrows) and located next to a muscle-specific kinase receptor gene, musk (right arrows).
To investigate the spatial and temporal expression patterns of zlpa1, we synthesized its antisense riboprobe and used it for the WISH analysis. The WISH analysis revealed that zlpa1 mRNAs were initially slightly ex-pressed in the entire embryo before the completion of gastrulation. During somitogenesis, the zlpa1 expres-sion domain was restricted to adaxial cells flanking the notochord, head, and tail buds (Fig. 2A, B). It was notable that 18-somite-stage embryos exhibited more-distinct individual zlpa1 expression domains restricted to adaxial cells and adjacent somites compared to 6-somite-stage embryos. At 24 hpf, zlpa1 was highly expressed in the anterior head region (including eyes, midbrain/hindbrain boundary, and brain mesoderm) and tail terminal. In addition, zlpa1expression was also observed in the dorsal aorta and vicinity (Fig. 2C). At the 31-hpf stage, the zlpa1expression domain was less prominent in the eyes and tail terminal but evident in brain mesoderm. In contrast, zlpa1 expression still remained in the dorsal aorta and vicinity (Fig. 2D). Transverse sections of the trunk region of 31-hpf em-bryos showed that zlpa1 was expressed in a portion of the dorsal aorta and vicinity, including hypochord. It was also sporadically observed in the outside peripheral muscle regions but was excluded from neural tube and notochord (Fig. 2E). Furthermore, to understand the zlpa1 developmental gene expression profile, we ana-lyzed its mRNA expression in zebrafish embryos at different stages from 1-cell to 7-dpf by RT-PCR. RT-PCR analysis showed that zlpa1 was expressed in all early embryos and larvae up to 7 dpf (Fig. 2F) and in all adult tissues examined (Fig. 2G). The zlpa1was expressed at higher levels in the brain, eyes, ovaries, testes, gills, and swim bladder (Fig. 2G).
Antisense MO interferes with zlpa1splicing and
results in pericardial and truck edema
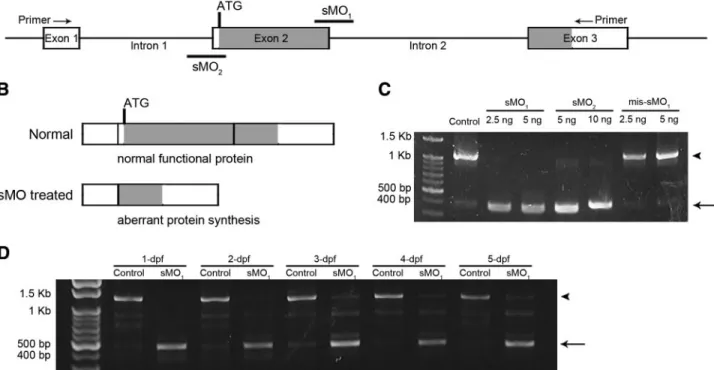
To study the roles of zLPA1during early
embryogene-sis, we used antisense MO oligonucleotides to knock-down zlpa1 gene activity. The zlpa1 gene consists of 3 exons and 2 introns, and a translation start site (ATG) is located at the 5⬘ terminal of exon 2 (Fig. 3A). Thus, we generated a splice-blocking MO (sMO1), targeting
the boundary between exon 2 and intron 2 to interfere with the splicing of exon 2. The sMO1was assumed to
skip the transcription of exon 2, which subsequently would result in aberrant protein synthesis (Fig. 3B). By the RT-PCR analysis, we demonstrated that the sMO1
caused a shortening of zlpa1transcripts from 1208 bp in untreated control embryos to 453 bp in sMO1-treated
embryos injected with 2.5 and 5 ng per embryo (Fig. 3C). The 453-bp PCR fragment was sequenced and compared to the 1208-bp fragment. Sequence compar-ison revealed that the exon 2 was not transcribed as expected (Supplemental Fig. S1). The similar effect was also observed in embryos treated with 1.25 ng per embryo. In addition, the zlpa1-splice-blocking activity of sMO1remained active in treated embryos from 1 dpf to
5 dpf (Fig. 3D). Thus, we used zlpa1-sMO1 to explore
LPA1’s function in zebrafish development.
To test the dosage response, we first injected differ-ent amounts (1.25–5 ng per embryo) of zlpa1-sMO1into
1-cell stage embryos. The zlpa1-sMO1-treated embryos
(zlpa
1morphants) showed normal morphology during
early development, but these zlpa1morphants gradually developed edema in the pericardial sac and body cavity (Fig. 4B) compared to the normal appearance of untreated zebrafish larvae (Fig. 4A) after 4 dpf.
Dose-Figure 2. Expression patterns of zlpa1. WISH
was performed in embryos at designated stages using a zlpa1antisense riboprobe. Anterior is to
the top (A, B) or left (C, D); dorsal and lateral views, respectively. A transverse section through the trunk (dorsal at the top) at the yolk exten-sion region is also presented (E) A) Six-somite stage. The gene, zlpa1, is expressed by adaxial
cells (arrow), head, and tail bud. B) Eighteen-somite stage. The zlpa1expression domain still
encompasses adaxial cells (arrow), head, and tail buds. More distinct individual zlpa1
expres-sion domains are shown in adaxial cells and vicinity. C) Twenty-four-hpf stage. The zlpa1
gene is highly expressed in the anterior head region and tail terminal. In addition, zlpa1 is
expressed in the dorsal aorta (arrowhead) and vicinity. D) Thirty-one-hpf stage. The zlpa1
ex-pression domain is localized to the brain region and diminished in the eye and tail terminal. The zlpa1 expression domain is still evident in
the dorsal aorta (arrowhead) and vicinity. E) Thirty-one-hpf stage. A transverse section re-veals that zlpa1 RNAs are condensed in part of
the dorsal aorta (white arrowhead). F, G) Ex-pression of zlpa1was also examined by
amplify-ing a 136-bp lpa1 fragment (top gels) at different embryonic stages (F) and in adult tissues (G) by RT-PCR. A 524-bp ef1␣
dependently, these morphants also gradually showed blood flow retardation and failure in ventricle contrac-tion (data not shown). Zebrafish embryos injected with 2.5 or 5 ng zlpa1-sMO1appeared very unhealthy;
there-fore, to avoid possible complications generated by high dosages, we subsequently used 1.25 ng unless otherwise stated. This dosage resulted in milder cardiovascular defects in ⬃30% of treated embryos, while the rest of embryos appeared healthy.
zlpa1morphants exhibit defects in thoracic duct
formation
At 1.25 ng zlpa1-sMO1, 41.4 ⫾ 5.6% (n⫽78), embryos
developed edema in the pericardial and trunk regions (Fig. 4B). This zlpa1-sMO1-induced edema differs from
those frequently observed edemas in various drug-treated or mutant zebrafish that mainly occur in the pericardial region. Thus, we decided to further exam-ine this aspect. The occurrence of edema indicates an
accumulation of interstitial fluids, which might result from vascular leakage or a failure of lymphatic vessels to absorb fluids. To examine the integrity of the vascula-ture, we examined the effects of zlpa1-sMO1in embryo
vasculature using both wild-type and vasculature-fluo-rescent Tg(fli1:EGFP)y1 zebrafish (37). No obvious de-fects in vasculature were observed in morphants, except some distortions of the patterning of the dorsal longi-tudinal anastomic vessel (DLAV), intersegmenal vessels (ISVs), supraintestinal artery (SIA), and subintetinal vein (SIV) after 5 dpf that might be caused by edema (data not shown).
To observe the lymphatic vasculature, we adapted an approach developed by two independent laboratories (38, 39) to monitor the development of lymphatic vessels in the presence or absence of zlpa1-sMO1. We
successfully recorded the development of the thoracic duct, the largest lymphatic vessel connecting to the cardinal vein, in a zebrafish larva from 3 to 5 dpf (see Supplemental Fig. S2). The thoracic duct initially forms
Figure 3.Antisense MOs induce splicing aberrant of zlpa1. A) Diagram of a partial map of zlpa1genomic DNA. Exons and introns
are shown in boxes and lines, respectively, and labeled with the corresponding exon or intron number. Two nonoverlapping
zlpa1sMOs, sMO1and sMO2, were designed to target the boundaries of exon 2/intron 2 and exon 2/intron 1, respectively. B)
The sMOs presumably eliminated the transcription of exon 2, which contains the translation start site, and resulted in aberrant protein synthesis. C) RT-PCR analysis using a primer pair as indicated in A shows that both sMO1and sMO2caused the deletion
of exon 2 and resulted in 453-bp PCR fragments (arrow) compared to the intact 1208-bp fragments (arrowhead) in control untreated and 5-bp mismatched sMO1 (mis-MO1) embryos. D) The zlpa1transcripts of 1- to 5-dpf embryos treated without
(control) and with 1.25 ng sMO1per embryo was analyzed by RT-PCR as described.
Figure 4. Ectopic expression of zlpa1 rescues
the splice-blocking MO induced edema. Ze-brafish embryos were untreated (A) or in-jected with 1.25 ng zlpa1sMO1(B, C). Edema
appeared in the pericardial sac and gut
vicin-ity (arrows) in 5-dpf morphants (B). The zlpa1 sMO1-induced edema was notably reduced by coinjecting 100 pg of the
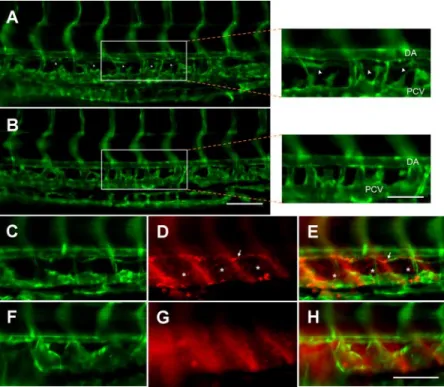
as isolated islands (Supplemental Fig. S2A, B), and the fusing of these islands gives rise to a complete vessel with lumen (Supplemental Fig. S2C, D). These obser-vations were similar to those reported by Yaniv et al. (39). In 5-dpf zlpa1morphants, thoracic ducts did not form (Fig. 5B, F) compared to those control ones with normal thoracic ducts (Fig. 5A, C). Although only some of the zlpa1morphants injected with 1.25 ng zlpa1-sMO1
exhibited mild cardiovascular defects, it was still possi-ble that the absence of thoracic ducts in those zlpa1 morphants might be a nonspecific secondary effect of cardiovascular defects. To further clarify this issue, we injected Tg(fli1:EGFP)y1 zebrafish embryos with 0.625 ng sMO1. We found no notable cardiovascular defects
in the treated embryos, and examined the integrity of their thoracic ducts. In four trials, 87.1⫾ 17.7% of 46 zlpa1morphants showed defects in thoracic duct forma-tion. Although the penetrance was lower and less consistent at this dosage, the experiment did show that the inhibition of thoracic duct formation by zlpa1-sMO1
was specific and not secondary to the cardiovascular defects. In addition, we also tested the function of the thoracic duct using an intramuscular injection of rho-damine dextran and observed that the injected rhoda-mine dextran was absorbed by the thoracic duct of the untreated zebrafish (Fig. 5D, E), but not that of the zlpa1morphants (Fig. 5G, H).
To assure the specificity of zlpa1-sMO1in inhibiting
thoracic duct formation and induction of edema, we examined the effects on zebrafish development of an sMO1 5-bp mismatched MO (mis-sMO1) and another
zlpa1 splice-blocking MO, sMO2, which targets the
boundary between intron 1 and exon 2. RT-PCR anal-ysis showed that the mis-sMO1 had no effect on zlpa1
transcript formation at up to 5 ng per embryo. In contrast, the sMO2caused missplicing of exon 2, as did
sMO1, but some residual transcripts still remained
intact, even when a higher amount of MO (10 ng/ embryo) was used (Fig. 3C). Sequence analysis of zlpa1 transcripts in those mis-sMO1 and sMO2 morphants
demonstrated that they had the same sequences as those of control and sMO1-treated embryos,
respec-tively (Supplemental Fig. S1). Using the numbers of segments between ISVs encompassed by the thoracic duct as an index, we analyzed the effects of different zlpa1 MOs on the formation of the thoracic duct. We considered a thoracic duct with⬎6 segments in length in the examined region as normal. In three indepen-dent experiments, 100.0 ⫾ 0.0% of both the control and the mis-sMO1-treated embryos contained a normal
thoracic duct. In contrast, there were significantly (P⬍0.01) lower percentages of embryos (9.1⫾3.5 and 25.7⫾6.1%) with a normal thoracic duct in the zlpa1 -sMO1- and zlpa1-sMO2-treated groups, respectively (Fig. 6A). The lower inhibitory effect of zlpa1-sMO2to block
thoracic duct formation even at a higher dosage (7.5 ng/embryo) was presumably due to its lower potency in interfering with zlpa1splicing, as described (Fig. 3C).
To confirm that zlpa1-sMO1-induced defects were
due to loss of zlpa1, we coinjected zlpa1-sMO1with an
expressing vector containing the zlpa1coding region, which does not contain the MO-targeting site, to see whether zlpa1-sMO1-induced defects could be
res-cued. Ectopic expression of zlpa1 fully or partially rescued ⬎80% of zebrafish larvae, which might have developed morphological defects, such as edema and the lack of a swim bladder, in the presence of zlpa1-sMO1 (Fig. 4C). The morphology of zlpa1
-res-Figure 5.Knockdown of zlpa1 inhibits
forma-tion of the thoracic duct, thus preventing lymphatic uptake. Zebrafish were aligned with the dorsal to the top and anterior to the left. Epifluorescent images collected with an⫻10 objective of the lateral views of the trunk vasculature from 5-dpf Tg(fli1:EGFP)y1
ze-brafish uninjected (A) or injected with 1.25 ng of zlpa1sMO1(B). Arrowheads indicate the
thoracic duct, which is present in the control embryo (A), but absent from the zlpa1
mor-phant (B). Boxed regions of A and B viewed with an ⫻20 objective are shown at right. Rhodamine dextran at 4.6 –9.2 g was in-jected intramuscularly into 5-dpf Tg(fli1:
EGFP)y1 zebrafish. After overnight culture,
epifluorescent images collected with an⫻20 objective were photographed to reveal the vasculature (C, F) and lymphatic vessels filled with rhodamine dextran (D, G) using FITC and rhodamine filters, respectively. In addi-tion, superimposed images of C, D and F, G are shown in panels E and H, respectively. It appears that rhodamine dextran was ab-sorbed by the lymphatic vessels, including the thoracic duct (arrows) and blind-ended lym-phatic vessels (*) between intersegmental ves-sels in control zebrafish (D, E), but the uptake by the thoracic duct is not apparent in the zlpa1morphant (G, H). DA, dorsal
cued zebrafish larvae (C) was indistinguishable from that of control ones (Fig. 4A). The zlpa1-rescued zebrafish larvae contained a significantly (P⬍0.05) higher percentage (57.5⫾12.9%) of the normal tho-racic duct (see representative photograph in Supple-mental Fig. S3C) compared to those injected with zlpa1-sMO1 only (9.4⫾1.0%, Fig. 6B). A higher
per-centage (18.9⫾4.6%) of rescued larvae was found to have a intermediately long thoracic duct (2 to 6 segments) compared to those of zlpa1 morphants (9.6⫾8.4%, Fig. 6B). Intriguingly, the lymphatic de-fect-inducing effect of zlpa1-sMO1could also be
par-tially ameliorated (37.9⫾3.3 and 29.3⫾7.0% embryos with a thoracic duct ⬎6 segments and 2 to 6 seg-ments, respectively; P⬍0.05) by the ectopic expres-sion of zebrafish vegf-c (Fig. 6C, n⫽4, see representa-tive photograph in Supplemental Fig. S3D).
DISCUSSION
LPA is a bioactive lipid that is thought to regulate angio-genesis in normal and tumor tissues (12, 25, 40). How-ever, mice lacking the LPA receptors, LPA1 and LPA1/
LPA2, show no detectable defects in vasculogenesis or
angiogenesis (9, 27), rendering the regulatory mecha-nism of LPA-mediated vascular responses a puzzle. In this study, we report that zLPA1was expressed in the dorsal
aorta and vicinity, and the knockdown of zLPA1resulted
in high penetrance loss of the thoracic duct. The loss of the thoracic duct was specific to a reduction in LPA1
activity, since it was consistently observed even at low zlpa1 MO dosages and could be rescued by the ectopic expres-sion of zlpa1. In addition, overexpression of zvegf-c, a well-known lymphangiogenic factor, was also able to par-tially ameliorate the zlpa1-morphant phenotypes. This further confirms the involvement of LPA1 in mediating
lymphangiogenesis in zebrafish. To our knowledge, this is a novel function of LPA receptor, and this is the first study implicating a G-protein-coupled receptor in the regula-tion of lymphangiogenesis.
Lysophospholipids, including LPA and sphingosine 1-phosphate (S1P), have long been implicated in car-diovascular development, where S1P appears to play a more prominent role in this aspect (41). The zebrafish mutant, Miles apart, which encodes S1P2, was found to
result in cardia bifida (42). Although this is the first instance to suggest the role of S1P receptor in verte-brate development, the S1P2-null mice have no
appar-ent anatomical or physiological defects at birth (43, 44). On the other hand, S1P1-null mice exhibit strong
defects in vascular smooth muscle recruitment, embry-onic hemorrhage, and death (45). In contrast to the prominent roles of S1P receptor in cardiovascular development, the LPA receptor-null mice do not reveal notable defects in the cardiovascular system (9, 27). However, half of the LPA1knockout mice died between
late embryonic development and weaning. Although the neonatal death of those LPA1 knockout mice was
attributed to a suckling defect, it would be interesting to examine the integrity of the lymphatic system of those knockout mice. In addition, autotoxin-deficient mice die at embryonic day 9.5 showing profound vascular defect in yolk sac (26). It further supports that LPA may work via one or more of its receptors to regulate vascular development.
There are at least five mammalian LPA receptors been identified, but only two of them, LPA1 (9, 10,
27)and LPA3 (28) have known in vivo functions by
using the knockout mouse approach. More recently, a G protein-coupled receptor, P2Y5, has been shown to be a novel LPA receptor, which is involved in mainte-nance of human hair growth (46). To explore zebrafish LPA receptors, we searched the zebrafish genome database and found LPA1, LPA2(NP_001003578), LPA3
(XP_694990), and a predicted sequence of LPA4
(XP_001334713). We have cloned LPA1–3but failed to
obtain the LPA4, which only has a predicted sequence
in the database. To compare their sequence similarity
Figure 6. zlpa1 splice-blocking morpholino (sMO) inhibits
thoracic duct formation and its rescue by the ectopic expres-sion of zlpa1or zvegf-c. Tg(fli1:EGFP)
y1zebrafish embryos were
injected without (control) or with 1.25 ng zlpa1sMO1(sMO1)
(A), 1.25 ng mismatched sMO1(mis-sMO1),or 7.5 ng sMO2
(sMO2); 1.25 ng zlpa1sMO1in the absence or presence of 100
ng of the pcDNA 3.1 vector with zlpa1(sMO1⫹zlpa1) (B), or vegf-c (sMO1⫹zvegf-c) at the 1-cell stage (C), and the length of
the thoracic duct was examined at 5-dpf under an epifluores-cent microscope by counting segments between 2 interseg-mental vessels. Percentages of embryos with ⬍6 segments (gray and open regions) were compared between treatments and their respective controls. *P⬍ 0.05; **P ⬍ 0.01.
to mammalian LPA1, those zebrafish LPA1– 4sequences
were aligned with sequences of human and mouse LPA1 (Supplemental Fig. S4). Sequence alignment
showed that zebrafish LPA1 is the closest homolog to
mammalian LPA1. The sequence similarities of
ze-brafish LPA1– 4compared to human or mouse LPA1are
89, 52, 45, and 12/13%, respectively (Supplemental Fig. S4B). In addition, the phylogenetic tree analysis further reveals that zebrafish LPA1is indeed the closest
homolog of mammalian LPA1 (Supplemental Fig.
S4C). The zLPA1 contains 7 putative transmembrane
domains (Fig. 1B). In addition, Valentine et al. (47) have identified 5 critical LPA binding amino acid residues, including R3.28, Q3.29, W4.64, D5.38, and K7.36, which correspond to the zLPA1amino acid 106,
107, 168, 186, and 276 (Fig. 1A), respectively, in the LPA receptors. Those amino acid residues are con-served except D5.38, in which an aspartic acid is replaced by an asparagine, in zLPA1as indicated in Fig.
1A. With the high sequence similarity and the conser-vation of ligand-binding residues to mammalian LPA1,
the zLPA1is more likely to be an LPA receptor.
In this study, we confirmed the ubiquitous expres-sion of zlpa1in various adult tissues (Fig. 2G) compared to its mouse homolog (8). The WISH analysis demon-strated that zlpa1expression spreads through the entire
blastodisc at a low level and is then restricted to adaxial cells, head, and tail buds at somatogenesis (Fig. 2A, B). Two types of muscle precursors are reported in ze-brafish somites. Adaxial cells are located next to the notochord and form future slow-muscle fibers, whereas lateral somitic cells give rise to future fast-muscle cells (48, 49). zlpa1appears only to be expressed in nonmi-gratory adaxial cells, since it was still located at the site juxtaposed to the notochord at the 18-somite stage (Fig. 2B). This expression pattern is similar to that of engrailed 1b and the lymphatic master gene, prox1, at similar stages (50, 51). These nonmigratory adaxial cells are muscle pioneers, which are known to be future slow-muscle cells that constitute the horizontal myosep-tum (48, 49). After 24 hpf, lpa1expression was heavily condensed in the eye and brain regions (Fig. 2C, D), which is similar to that of its mammalian homologue (8). Its expression was also found in the vicinity of the dorsal aorta (Fig. 2E) where vegf-c was expressed (52) The zlpa1expression domain encompassed only part of the dorsal aorta (Fig. 2E). In contrast, the zvegf-c expression domain enclosed the entire dorsal aorta (data not shown). These partially overlapping signals of zlpa1 and zvegf-c at the dorsal aorta suggest a possible linkage between them. In addition, the zlpa1expression domain appeared to include the hypochord, a transient structure in intimate association with notochord and dorsal aorta (53). The hypochord is known to express VEGF and is essential for the development of dorsal aorta (54). The expression of zLPA1 in the hypochord and
dorsal aorta suggests a role of zLPA1 in vasculargenesis.
However, knockdown of zLPA1appeared to have limited
effects on blood vessels in zebrafish. It implies that the zLPA1 may not be essential or other factors may be
complementary to its regulation of blood vessels. The lack of noticeable morphological defects ob-served for zlpa1morphants in the early stages suggests that zPLA1 is not essential for early development,
including blood vasculature formation. In contrast, the later appearance of edema in zlpa1morphants implies a tissue fluid imbalance. The requirement of adequate fluid balance increases with growth of zebrafish larvae. Therefore, the need for a functioning thoracic duct increases later in larval development. As shown in Supplemental Fig. S2 and also in results by Yaniv et al. (39), a nearly complete thoracic duct is not formed until 5 dpf in zebrafish. The development of the thoracic duct coincides nicely to our observation that the edema of zlpa1morphants was also shown gradually after 4 or 5 dpf. Our observation that a functional thoracic duct was not formed on the treatment of zlpa1-MO (Fig. 5) further indicates that the edema was due to the lack of a thoracic duct. The blood vessels appeared normal prior to 5-dpf stage; however, we can not rule out the possibility that the vascular leakage might also contribute to the formation of edema in those zlpa1morphants.
The late appearance of zlpa1-MO phenotype raises the concern that it might not be a primary effect of MO since most MOs’ activity are known to last for only 3 or 4 days in zebrafish embryos. In this regard, our RT-PCR analysis unequivocally showed that the zlpa1-MO still retained its splice-blocking activity in 5-dpf embryos, the oldest embryos examined in this study (Fig. 3D). In addition, the splice blocking of antisense MO oligonu-cleotides is a commonly used approach to effectively knock down gene activity in zebrafish. However, one concern could be that the observed zlpa1-morphant phenotypes might be due to the dominant-negative effects of aberrantly translated proteins, but not the effects of zlpa1 gene knockdown. Although MO-gener-ated splice variants may be translMO-gener-ated, they are often degraded after the first translation by nonsense-medi-ated decay, especially if the splice modification results in a frameshift. To further address this issue, we have sequenced the PCR products generated by the splice variants of sMO1 and sMO2. Both splice variants, as
expected, had eliminated the zlpa1Exon 2, as shown in Supplemental Fig. S1. By the presence of a translation start site (ATG), we could identify 7 potential protein products with only one inframe protein that might be synthesized (Supplemental Fig. S1). With the assump-tion that those frameshift products were more likely to be degraded, we only discuss the possible effect of the inframe protein further. This possible inframe product is a 59-amino acid protein that comprises the end of the 7th transmembrane domain and the rest of C-terminal tail. By blasting this 59-amino acid sequence to the zebrafish protein database, there were only 5 hits except zLPA1(data not shown). A hypothetical protein
18 in 25 amino acids. With the low-sequence homolo-gies to other zebrafish proteins, even if this 59-amino acid protein were to produce a dominant-negative effect, it would still be specific to the zLPA1. Thus, the
possible dominant-negative effect should not affect our conclusion. Furthermore, along with the zlpa1 rescue experiments and consistent phenotypes obtained by the sMO2 (but not the 5-bp mismatched sMO1), we
conclude that the loss of thoracic duct is specifically induced by knocking down zLPA1. We currently do not
know whether the block of thoracic duct formation was a direct consequence of zLPA1 knockdown; however,
LPA has been shown to prime LPA receptor-expressing cells such as astrocytes to produce a specific secondary response to promote neuronal differentiation (55). Therefore, the loss of the thoracic duct in zlpa1 mor-phants is at least a specific secondary response by zlpa1 knockdown, if not a primary one.
The present study suggests a possible link of LPA1
with lymphangiogenesis. LPA1-mediated
lymphangio-genesis may be dependent on or independent of VEGF-C signaling. Although LPA did induce LPA1
-dependent expression of vegf-c in HUVECs in our preliminary trials (unpublished results), we failed to demonstrate consistent modulation of vegf-c expression on lpa1 knockdown or ectopic expression in zebrafish embryos by real-time PCR or WISH analyses (data not shown). These results imply that LPA and VEGF-C may work in parallel to modulate lymphatic vessel develop-ment. This finding is important because it provides a clue for examining how LPA may participate in cancer formation. LPA stimulates cell proliferation, migration, and survival. These LPA-evoked responses are hall-marks of tumor growth and metastasis (56). Thus, when significant LPA levels were found to be present in malignant effusions and its receptors were expressed in several human cancer cells, it became an emerging candidate for a cancer marker (4). LPA was also suggested to be a tumor inducer due to its capability of inducing cell motility and invasiveness, especially in both ECs (14 –16) and tumor cells (57). VEGF-C and its receptor, VEGFR-3, are expressed in lymphatic endo-thelial cells and a variety of human tumor cells. There-fore, activation of the VEGF-C/VEGFR-3 axis in con-junction of LPA1 signaling in lymphatic ECs can
facilitate metastasis by stimulating lymphangiogenesis within and around tumors (58 – 60).
In summary, we have discovered a novel function of the LPA receptor, LPA1, as being a key receptor for
mediating thoracic duct formation in zebrafish. Signal-ing for LPA1-mediated lymphangiogenesis possibly
oc-curs in parallel to that of VEGF-C. These findings can assist our understanding of the molecular control of lymphangiogenesis in vivo, as well as help in unraveling the possible role of LPA in tumor metastasis through its regulation of lymphangiogenesis.
This work was supported by a grant (NSC95–2311-B-002– 015-MY3) to S.J.L. from the National Science Council of Taiwan.
REFERENCES
1. Adams, R. H., and Alitalo, K. (2007) Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 8, 464 – 478
2. Oliver, G. (2004) Lymphatic vasculature development. Nat. Rev.
4,35– 45
3. Cleaver, O., and Melton, D. A. (2003) Endothelial signaling during development. Nat. Med. 9, 661– 668
4. Mills, G. B., and Moolenaar, W. H. (2003) The emerging role of lysophosphatidic acid in cancer. Nat. Rev. Cancer 3, 582–591 5. Anliker, B., and Chun, J. (2004) Cell surface receptors in
lysophospholipid signaling. Semin. Cell Dev. Biol. 15, 457– 465 6. Meyer zu Heringdorf, D., and Jakobs, K. H. (2007)
Lysophospho-lipid receptors: signalling, pharmacology and regulation by lyso-phospholipid metabolism. Biochim. Biophys. Acta 1768, 923–940 7. Moolenaar, W. H. (1999) Bioactive lysophospholipids and their
G protein-coupled receptors. Exp. Cell Res. 253, 230 –238 8. Hecht, J. H., Weiner, J. A., Post, S. R., and Chun, J. (1996)
Ventricular zone gene-1 (vzg-1) encodes a lysophosphatidic acid receptor expressed in neurogenic regions of the developing cerebral cortex. J. Cell Biol. 135, 1071–1083
9. Contos, J. J., Fukushima, N., Weiner, J. A., Kaushal, D., and Chun, J. (2000) Requirement for the lpA1 lysophosphatidic acid receptor gene in normal suckling behavior. Proc. Natl. Acad. Sci.
U. S. A. 97, 13384 –13389
10. Estivill-Torrus, G., Llebrez-Zayas, P., Matas-Rico, E., Santin, L., Pedraza, C., De Diego, I., Del Arco, I., Fernandez-Llebrez, P., Chun, J., and De Fonseca, F. R. (2008) Absence of LPA1 signaling results in defective cortical development. Cereb. Cortex
18,938 –950
11. Lin, C. I., Chen, C. N., Lin, P. W., Chang, K. J., Hsieh, F. J., and Lee, H. (2007) Lysophosphatidic acid regulates inflammation-related genes in human endothelial cells through LPA1and
LPA3. Biochem. Biophys. Res. Commun. 363, 1001–1008.
12. English, D., Kovala, A. T., Welch, Z., Harvey, K. A., Siddiqui, R. A., Brindley, D. N., and Garcia, J. G. (1999) Induction of endothelial cell chemotaxis by sphingosine 1-phosphate and stabilization of endothelial monolayer barrier function by lyso-phosphatidic acid, potential mediators of hematopoietic angio-genesis. J. Hematother. Stem Cell Res. 8, 627– 634
13. Tager, A. M., Lacamera, P., Shea, B. S., Campanella, G. S., Selman, M., Zhao, Z., Polosukhin, V., Wain, J., Karimi-Shah, B. A., Kim, N. D., Hart, W. K., Pardo, A., Blackwell, T. S., Xu, Y., Chun, J., and Luster, A. D. (2008) The lysophosphatidic acid receptor LPA1
links pulmonary fibrosis to lung injury by mediating fibroblast recruitment and vascular leak. Nat. Med. 14, 45–54
14. Lee, H., Goetzl, E. J., and An, S. (2000) Lysophosphatidic acid and sphingosine 1-phosphate stimulate endothelial cell wound healing. Am. J. Physiol. Cell Physiol. 278, C612–C618
15. Panetti, T. S. (2002) Differential effects of sphingosine 1-phos-phate and lysophosphatidic acid on endothelial cells. Biochim.
Biophys. Acta 1582, 190 –196
16. Langlois, S., Gingras, D., and Beliveau, R. (2004) Membrane type 1-matrix metalloproteinase (MT1-MMP) cooperates with sphingosine 1-phosphate to induce endothelial cell migration and morphogenic differentiation. Blood 103, 3020 –3028 17. Wu, W. T., Chen, C. N., Lin, C. I., Chen, J. H., and Lee, H. (2005)
Lysophospholipids enhance matrix metalloproteinase-2 expres-sion in human endothelial cells. Endocrinology 146, 3387–3400 18. Lin, C. I., Chen, C. N., Chen, J. H., and Lee, H. (2006)
Lysophospholipids increase IL-8 and MCP-1 expressions in human umbilical cord vein endothelial cells through an IL-1-dependent mechanism. J. Cell. Biochem. 99, 1216 –1232 19. Schulze, C., Smales, C., Rubin, L. L., and Staddon, J. M. (1997)
Lysophosphatidic acid increases tight junction permeability in cultured brain endothelial cells. J. Neurochem. 68, 991–1000 20. Cerutis, D. R., Nogami, M., Anderson, J. L., Churchill, J. D.,
Romberger, D. J., Rennard, S. I., and Toews, M. L. (1997) Lysophosphatidic acid and EGF stimulate mitogenesis in human airway smooth muscle cells. Am. J. Physiol. 273, L10 –L15 21. Hayashi, K., Takahashi, M., Nishida, W., Yoshida, K., Ohkawa, Y.,
Kitabatake, A., Aoki, J., Arai, H., and Sobue, K. (2001) Pheno-typic modulation of vascular smooth muscle cells induced by unsaturated lysophosphatidic acids. Circ. Res. 89, 251–258
22. Toews, M. L., Ustinova, E. E., and Schultz, H. D. (1997) Lysophosphatidic acid enhances contractility of isolated airway smooth muscle. J. Appl. Physiol. 83, 1216 –1222
23. Tokumura, A., Iimori, M., Nishioka, Y., Kitahara, M., Sakashita, M., and Tanaka, S. (1994) Lysophosphatidic acids induce pro-liferation of cultured vascular smooth muscle cells from rat aorta. Am. J. Physiol.Cell Physiol 267, C204 –C210
24. Tokumura, A., Yotsumoto, T., Masuda, Y., and Tanaka, S. (1995) Vasopressor effect of lysophosphatidic acid on spontaneously hypertensive rats and Wistar-Kyoto rats. Res. Commun. Molec.
Pathol. Pharmacol. 90, 96 –102
25. Yoshida, K., Nishida, W., Hayashi, K., Ohkawa, Y., Ogawa, A., Aoki, J., Arai, H., and Sobue, K. (2003) Vascular remodeling induced by naturally occurring unsaturated lysophosphatidic acid in vivo. Circulation 108, 1746 –1752
26. Van Meeteren, L. A., Ruurs, P., Stortelers, C., Bouwman, P., van Rooijen, M. A., Pradere, J. P., Pettit, T. R., Wakelam, M. J., Saulnier-Blache, J. S., Mummery, C. L., Moolenaar, W. H., and Jonkers, J. (2006) Autotaxin, a secreted lysophospholipase D, is essential for blood vessel formation during development. Mol.
Cell. Biol. 26, 5015–5022
27. Contos, J. J., Ishii, I., Fukushima, N., Kingsbury, M. A., Ye, X., Kawamura, S., Brown, J. H., and Chun, J. (2002) Characterization of lpa2(Edg4) and lpa1/lpa2 (Edg2/Edg4) lysophosphatidic acid receptor knockout mice: signaling deficits without obvious pheno-typic abnormality attributable to lpa2. Mol. Cell. Biol. 22, 6921– 6929 28. Ye, X., Hama, K., Contos, J. J., Anliker, B., Inoue, A., Skinner, M. K., Suzuki, H., Amano, T., Kennedy, G., Arai, H., Aoki, J., and Chun, J. (2005) LPA3-mediated lysophosphatidic acid signalling
in embryo implantation and spacing. Nature 435, 104 –108 29. Noguchi, K., Ishii, S., and Shimizu, T. (2003) Identification of
p2y9/GPR23 as a novel G protein-coupled receptor for lyso-phosphatidic acid, structurally distant from the Edg family.
J. Biol. Chem. 278, 25600 –25606
30. Lee, C. W., Rivera, R., Gardell, S., Dubin, A. E., and Chun, J. (2006) GPR92 as a new G12/13- and Gq-coupled lysophospha-tidic acid receptor that increases cAMP, LPA5. J. Biol. Chem. 281,
23589 –23597
31. Weinstein, B. M. (2002) Plumbing the mysteries of vascular development using the zebrafish. Sem. Cell Dev. Biol. 13, 515–522 32. Ny, A., Autiero, M., and Carmeliet, P. (2006) Zebrafish and
Xenopus tadpoles: small animal models to study angiogenesis
and lymphangiogenesis. Exp. Cell Res. 312, 684 – 693
33. Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., and Schilling, T. F. (1995) Stages of embryonic development of the zebrafish. Dev. Dyn. 203, 253–310
34. Thisse, C., Thisse, B., Schilling, T. F., and Postlethwait, J. H. (1993) Structure of the zebrafish snail1 gene and its expression in wild-type, spadetail and no tail mutant embryos. Development
119,1203–1215
35. Ober, E. A., Olofsson, B., Makinen, T., Jin, S. W., Shoji, W., Koh, G. Y., Alitalo, K., and Stainier, D. Y. (2004) Vegfc is required for vascular development and endoderm morphogenesis in ze-brafish. EMBO Rep. 5, 78 – 84
36. Lai, S. L., Chang, C. N., Wang, P. J., and Lee, S. J. (2005) Rho mediates cytokinesis and epiboly via ROCK in zebrafish. Mol.
Reprod. Dev. 71, 186 –196
37. Lawson, N. D., Scheer, N., Pham, V. N., Kim, C. H., Chitnis, A. B., Campos-Ortega, J. A., and Weinstein, B. M. (2001) Notch signaling is required for arterial-venous differentiation during embryonic vascular development. Development 128, 3675–3683 38. Kuchler, A. M., Gjini, E., Peterson-Maduro, J., Cancilla, B.,
Wolburg, H., and Schulte-Merker, S. (2006) Development of the zebrafish lymphatic system requires VEGFC signaling. Curr. Biol.
16,1244 –1248
39. Yaniv, K., Isogai, S., Castranova, D., Dye, L., Hitomi, J., and Weinstein, B. M. (2006) Live imaging of lymphatic development in the zebrafish. Nat. Med. 12, 711–716
40. Hu, Y. L., Tee, M. K., Goetzl, E. J., Auersperg, N., Mills, G. B., Ferrara, N., and Jaffe, R. B. (2001) Lysophosphatidic acid induction of vascular endothelial growth factor expression in human ovarian cancer cells. J. Natl. Cancer Inst. 93, 762–768 41. Birgbauer, E., and Chun, J. (2006) New developments in the
biological functions of lysophospholipids. Cell. Mol. Life Sci. 63, 2695–2701
42. Kupperman, E., An, S., Osborne, N., Waldron, S., and Stainier, D. Y. (2000) A sphingosine-1-phosphate receptor regulates cell
migration during vertebrate heart development. Nature 406, 192–195
43. Ishii, I., Ye, X., Friedman, B., Kawamura, S., Contos, J. J., Kingsbury, M. A., Yang, A. H., Zhang, G., Brown, J. H., and Chun, J. (2002) Marked perinatal lethality and cellular signaling deficits in mice null for the two sphingosine 1-phosphate (S1P) receptors, S1P2/LPB2/EDG-5 and S1P3/LPB3/EDG-3. J. Biol. Chem. 277, 25152–25159
44. MacLennan, A. J., Carney, P. R., Zhu, W. J., Chaves, A. H., Garcia, J., Grimes, J. R., Anderson, K. J., Roper, S. N., and Lee, N. (2001) An essential role for the H218/AGR16/Edg-5/LPB2
sphingosine 1-phosphate receptor in neuronal excitability. Eur.
J. Neurosci. 14, 203–209
45. Liu, Y., Wada, R., Yamashita, T., Mi, Y., Deng, C. X., Hobson, J. P., Rosenfeldt, H. M., Nava, V. E., Chae, S. S., Lee, M. J., Liu, C. H., Hla, T., Spiegel, S., and Proia, R. L. (2000) Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, is essen-tial for vascular maturation. J. Clin. Invest. 106, 951–961 46. Pasternack, S. M., von Kugelgen, I., Aboud, K. A., Lee, Y. A.,
Ruschendorf, F., Voss, K., Hillmer, A. M., Molderings, G. J., Franz, T., Ramirez, A., Nurnberg, P., Nothen, M. M., and Betz, R. C. (2008) G protein-coupled receptor P2Y5 and its ligand LPA are involved in maintenance of human hair growth. Nat.
Genet. 40, 329 –334
47. Valentine, W. J., Fells, J. I., Perygin, D. H., Mujahid, S., Yokoyama, K., Fujiwara, Y., Tsukahara, R., Van Brocklyn, J. R., Parrill, A. L., and Tigyi, G. (2008) Subtype-specific residues involved in ligand activation of the endothelial differentiation gene family lysophosphatidic acid receptors. J. Biol. Chem. 283, 12175–12187
48. Devoto, S. H., Melancon, E., Eisen, J. S., and Westerfield, M. (1996) Identification of separate slow and fast muscle precursor cells in vivo, prior to somite formation. Development 122, 3371–3380 49. Stickney, H. L., Barresi, M. J., and Devoto, S. H. (2000) Somite
development in zebrafish. Dev. Dyn. 219, 287–303
50. Thisse, B., and Thisse, C. (2004) Fast release clones: a high throughput expression analysis. Retrieved October 19, 2007, from http://zfin.org; ZFIN ID: ZDB-PUB-040907-1
51. Thisse, B., and Thisse, C. (2005) High throughput expression analysis of ZF-models consortium clones. Retrieved October 19, 2007, from http://zfin.org; ZFIN ID: ZDB-PUB-051025-1 52. Covassin, L. D., Villefranc, J. A., Kacergis, M. C., Weinstein,
B. M., and Lawson, N. D. (2006) Distinct genetic interactions between multiple Vegf receptors are required for development of different blood vessel types in zebrafish. Proc. Natl. Acad. Sci.
U. S. A. 103, 6554 – 6559
53. Eriksson, J., and Lofberg, J. (2000) Development of the hypo-chord and dorsal aorta in the zebrafish embryo (Danio rerio). J.
Morphol. 244, 167–176
54. Cleaver, O., and Krieg, P. A. (1998) VEGF mediates angioblast migration during development of the dorsal aorta in Xenopus.
Development 125, 3905–3914
55. Spohr, T. C., Choi, J. W., Gardell, S. E., Herr, D., Rehen, S. K., Gomes, F. C., and Chun, J. (2008) LPA receptor-dependent secondary effects via astrocytes promote neuronal differentia-tion. J. Biol. Chem. 283, 7470 –7479
56. Hanahan, D., and Weinberg, R. A. (2000) The hallmarks of cancer. Cell 100, 57–70
57. Imamura, F., Horai, T., Mukai, M., Shinkai, K., Sawada, M., and Akedo, H. (1993) Induction of in vitro tumor cell invasion of cellular monolayers by lysophosphatidic acid or phospholipase D. Biochem. Biophys. Res. Commun. 193, 497–503
58. Akagi, K., Ikeda, Y., Miyazaki, M., Abe, T., Kinoshita, J., Mae-hara, Y., and Sugimachi, K. (2000) Vascular endothelial growth factor-C (VEGF-C) expression in human colorectal cancer tis-sues. Brit. J. Cancer 83, 887– 891
59. Niki, T., Iba, S., Tokunou, M., Yamada, T., Matsuno, Y., and Hirohashi, S. (2000) Expression of vascular endothelial growth factors A, B, C, and D and their relationships to lymph node status in lung adenocarcinoma. Clin. Cancer Res. 6, 2431–2439 60. Su, J. L., Yen, C. J., Chen, P. S., Chuang, S. E., Hong, C. C., Kuo,
I. H., Chen, H. Y., Hung, M. C., and Kuo, M. L. (2007) The role of the VEGF-C/VEGFR-3 axis in cancer progression. Brit. J.
Cancer 96, 541–545
Received for publication January 29, 2008. Accepted for publication June 12, 2008.