eScholarship provides open access, scholarly publishing services to the University of California and delivers a dynamic research platform to scholars worldwide.
Peer Reviewed
Title:
Copepod diversity and composition as indicators of intrusion of the Kuroshio Branch Current into the northern Taiwan Strait in spring 2000
Author:
Hsieh, Chih-hao H, Chiu, T S, Shih, C T
Publication Date:
04-01-2004
Publication Info:
UC San Diego, Postprints
Permalink:
http://escholarship.org/uc/item/4b45g5wk
Citation:
Hsieh, Chih-hao H, Chiu, T S, & Shih, C T. (2004). Copepod diversity and composition as indicators of intrusion of the Kuroshio Branch Current into the northern Taiwan Strait in spring 2000. UC San Diego: Retrieved from: http://escholarship.org/uc/item/4b45g5wk
Abstract:
Copepod diversity and composition as indicators of the intrusion of the Kuroshio Branch Current into the northern Taiwan Strait in spring 2000. Zoological Studies 43(2): 393-403. This study was carried out during the waning of the northeastern monsoon, at which time the Kuroshio Branch Current intrudes into the northern Taiwan Strait. The Kuroshio Branch Current was characterized by high temperature and salinity in contrast to the low temperature and salinity of the China Coastal Current. A front was induced south of the Haitan Island, which resulted in a phytoplankton bloom when the China Coastal Current and Kuroshio Branch Current met. The front is marked by a temperature of 18 +/- 2 degreesC and a salinity of 33 +/- 0.5 psu. The intrusion of the Kuroshio Branch Current apparently influenced the species diversity of copepods. Species richness and diversity were higher in the area affected by the Kuroshio Branch Current than that affected by the China Coastal Current. No significant differences in the copepod abundances between the two water masses were observed. Species assemblages were distinct in the two water masses. The China Coastal Current contained only coastal/neritic cold-water species, while the Kuroshio Branch Current consisted of coastal, neritic, and oceanic warm-water species. Indicator species of each water mass were identified and are discussed. http://www.sinica.edu.tw/zool/ zoolstud/43.2/393.pdf.
Copepod Diversity and Composition as Indicators of Intrusion of the
Kuroshio Branch Current into the Northern Taiwan Strait in Spring 2000
Chih-Hao Hsieh1,*, Tai-Sheng Chiu1and Chang-tai Shih2,3,4
1Department of Zoology, National Taiwan University, Taipei, Taiwan 106, R.O.C. 2Taiwan Fisheries Research Institute, Keelung, Taiwan 202, R.O.C.
3Canadian Museum of Nature, Ottawa, Canada K1P-6P4
4Department of Marine Resources, National Sun Yat-sen University, Kaohsiung, Taiwan 804, R.O.C.
(Accepted February 13, 2004)
Chih-Hao Hsieh, Tai-Sheng Chiu and Chang-tai Shih (2004) Copepod diversity and composition as
indica-tors of the intrusion of the Kuroshio Branch Current into the northern Taiwan Strait in spring 2000. Zoological
Studies 43(2): 393-403. This study was carried out during the waning of the northeastern monsoon, at which
time the Kuroshio Branch Current intrudes into the northern Taiwan Strait. The Kuroshio Branch Current was characterized by high temperature and salinity in contrast to the low temperature and salinity of the China Coastal Current. A front was induced south of the Haitan Island, which resulted in a phytoplankton bloom when the China Coastal Current and Kuroshio Branch Current met. The front is marked by a temperature of 18 ± 2
°
C and a salinity of 33 ± 0.5 psu. The intrusion of the Kuroshio Branch Current apparently influenced the species diversity of copepods. Species richness and diversity were higher in the area affected by the Kuroshio Branch Current than that affected by the China Coastal Current. No significant differences in the copepod abundances between the two water masses were observed. Species assemblages were distinct in the two water masses. The China Coastal Current contained only coastal/neritic cold-water species, while the Kuroshio Branch Current consisted of coastal, neritic, and oceanic warm-water species. Indicator species of each water mass were identified and are discussed. http://www.sinica.edu.tw/zool/zoolstud/43.2/393.pdfKey words: China Coastal Current, Kuroshio Branch Current, Indicator, Species diversity, Species
composi-tion.
T
he Taiwan Strait is a shallow channel con-necting the East China Sea and South China Sea and serves as an important pathway for faunal exchange between these two waters in the margin of the western North Pacific. It is also a main route for migratory fishes and thus an important fishing ground. Three main currents, the China Coastal Current (CCC), Kuroshio Branch Current (KBC), and South China Sea Surface Current (SCSSC), all driven by the monsoon system, affect the hydrographic conditions of the Strait (Jan et al. 2002). When the northeastern mon-soon prevails during winter (Nov. to Mar.), the CCC moves southward into the Strait; meanwhile, the monsoon also forces the KBC to penetrate into the Strait through the Penghu Channel (Wang andChern 1988). Stagnated by the CCC, the KBC is blocked south of the Changyun Ridge when the northeastern monsoon is strong. Only when the northeastern monsoon wanes in the spring, does the KBC flow over the Changyun Ridge or move northwestward along the local isobaths into the northern Strait. A front south of the Haitan Is. is induced when the northwardly flowing KBC encounters the retreating CCC. During summer (June to Aug.), the southwestern monsoon causes the SCSSC to move northward into the Strait in place of the KBC. The seasonal variation in circu-lation of the Taiwan Strait was described by Jan et al. (2002). The physical oceanography of the intrusion of the KBC into the Strait during the wan-ing of northeastern monsoon has been well
stud-393
*To whom correspondence and reprint requests should be addressed. Present address: Scripps Institution of Oceanography, University of California-San Diego, California, 92093-0208, USA. E-mail:[email protected]
ied (Jan et al. 1998); however, the biological effects of this circulation are not known.
The CCC is a neritic water mass character-ized by low salinity, low temperature, and high nutrient due to river runoff from the mainland China (Liu et al. 2000). The KBC, which originates from the oceanic Kuroshio Current, has high tem-perature and high salinity, but counter-intuitively contains a nutrient level comparable to that of the CCC (Chung et al. 2001). It is known that zoo-plankton composition and abundance are affected by the properties of water masses (Boucher et al. 1987, Kouwenberg 1994, Lopes et al. 1999). Since our samples were collected across both the CCC and KBC, this survey allowed us to test the following hypotheses: (i) distinct species composi-tions occur in the CCC and KBC; (ii) low species diversity occurs in the CCC (a neritic water mass subject to frequent disturbances) and high diversity occurs in the KBC (an oceanic water mass with greater stability); and (iii) equal abundances of copepods occur in the CCC and KBC according to their similar nutrient contents.
Previous faunal surveys in the Taiwan Strait have been either confined to a small area or con-ducted without attention to hydrographic effects. Tzeng and Wang (1992 1993) studied the compo-sition and seasonal variation of larval fish fauna in the Tanshui River estuary off northwestern Taiwan; Chiu and Chang (1995) reported diurnal cycling in the vertical distribution of larval fishes at a fixed station in the northeastern Strait; Chiu and Chang (1994) compared the larval fish compositions dur-ing winter and sprdur-ing in the northeastern Strait; Chiu (1992) and Chiu and Chen (1997) examined the larval fish composition in relation to the hydrog-raphy in the southern Strait. The copepod fauna was reported by Tan (1967) around the Tanshui River estuary; Hsieh and Chiu (1998) compared the species assemblages and abundances of copepods in the Tanshui River estuary and adja-cent area; Zheng et al. (1982) and Chen (1992) reported copepod species in the Strait. A relatively large-scale survey including benthic, planktonic, and larval fish faunas in the Strait was conducted by Zhu et al. (1988), but was limited to the western Strait and hydrographic effects on biological varia-tion were not examined. The present study is thus a preliminary survey of copepods covering the northern Taiwan Strait and analyzes hydrographic effects on copepod assemblages. Copepods are a major component of marine zooplankton and play an important role in marine food webs (Runge 1988, Poulet and Williams 1991, Williams et al.
1994). Studies on copepods are crucial for under-standing the dynamics of marine ecosystems and are necessary for purposes of management and conservation of marine resources. The objectives of our study are to provide basic information about the copepod fauna and to determine the effects of the Kuroshio intrusion in the Taiwan.
MATERIALS AND METHODS Sampling
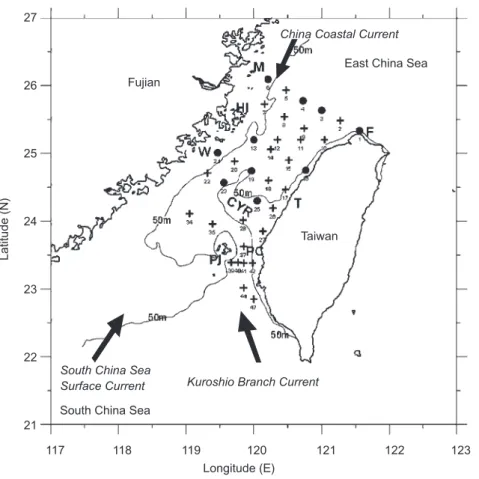
A cruise was carried out on board the Ocean Research I on 20~25 Mar. 2000 in the Taiwan Strait (Fig. 1). Copepods were sampled using a standard North Pacific zooplankton net with a mouth diameter of 45 cm and mesh size of 150 µm, and samples were preserved in seawater with 5% formaldehyde. A flowmeter was mounted at the center of the net mouth to record the volume of water filtered. The net was towed obliquely from near the bottom to the surface. Vertical profiles of temperature, salinity, fluorescence, and dissolved oxygen were recorded at each station from the bottom to the surface with a conductivity-tempera-ture-depth (CTD) profiler (Sea-Bird Electronics, Bellevue, USA) equipped with a fluorometer. The fluorescence value was not calibrated but used as a reference index of the relative phytoplankton bio-mass. During the cruise, plankton samples were collected at only 10 of 36 stations, because the ship time allocated for our study was limited.
In the laboratory, copepods were sub-sam-pled using a Folsom splitter until the sample size was reduced to 300~500 specimens. Copepods (including copepodid stages) were identified to species whenever possible. The abundance of copepods was expressed as the number of individ-uals/m3.
Data analysis
Shannon,s diversity index was used to mea-sure the species diversity of each sampling station. A correspondence analysis based on the relative abundance of the 20 most abundant species was applied to simultaneously examine the station associations and the corresponding species (Pielou 1984). For reducing the patchiness effect of zooplankton, data were square-root transformed (Krebs 1989). K-means classification based on the station scores of the correspondence analysis was applied to distinguish station groups
(Wilkinson 1990). By using K-means classifica-tion, the number of groups was decided in advance (in the current case, the number of groups was two), and an iterative algorithm was used to assign stations to the groups by maximiz-ing the variance (of input variables) between groups and minimizing the variance within groups. This procedure results in an objective classification of stations. Indicator species of each station groups were extracted by an indicator species analysis (Dufrene and Legendre 1997). The indi-cator value (Ind Val) of each species was comput-ed as follows:
Ind Val = RAkjx RFkjx 100;
where RAkj is the relative abundance of species j in group k, and RFkjis the relative frequency (pres-ence/absence) of species j in group k. The highest indicator value for a given species across groups was saved as a summary of the overall indicator
value of that species. A Monte Carlo method was used to test the significance of their indicator val-ues by randomly permuting the data and recalcu-lating Ind Val. One thousand permutations were made to compare the observed Ind Val and recal-culated Ind Val. The result was deemed significant when the observed Ind Val fell in the 5% upper tail. Only individuals identified to species were included in the above analyses. A Mann-Whitney U test was used to test for differences in copepod abun-dance, species richness, and diversity between the station groups affected by the China Coastal Current and Kuroshio Branch Current.
RESULTS
The two water masses have distinctive tem-perature-salinity (T-S) properties (Fig. 2). Stations 1, 3, 4, 6, and 21 characterized by low temperature and salinity were influenced by the CCC, while
sta-Fig. 1. Map showing sampling stations and isobaths in the Taiwan Strait. Crosses indicate stations with CTD data only; filled circles
indicate stations with both CTD and copepod samples; arrows indicate the main currents during the cruise; HI, Haitan Island; PI, Penghu Island; PC, Penghu Channel; CYR, Changyun ridge; F, Cape Fuguei; M, Matsu; W, Wuchiu; T, Taichung.
Latitude (N)
Longitude (E)
East China Sea
China Coastal Current
Taiwan Fujian
Kuroshio Branch Current
South China Sea 27 26 25 24 23 22 21 117 118 119 120 121 122 123
South China Sea Surface Current
tions 16, 19, 23, and 25 characterized by high tem-perature and salinity indicated intrusion of the KBC. Station 13 was at the transitional location with a broad range of temperature and salinity. The T-S curve of station 41 (no plankton sample, c.f. Fig. 1) was used to highlight the intrusion of the KBC into the Penghu Channel. Typical T-S curves1of the CCC, KBC, and SCSSC were also
plotted in figure 2 for comparison. The SCSSC is distinct from the KBC by having higher tempera-ture and lower salinity in the surface layer. This distinction provides supportive evidence that our samples were taken during the intrusion of the KBC. Since the purpose of this work was to com-pare the CCC and KBC, distinguishing the KBC and SCSSC is not discussed.
1.The typical T-S curves represent characteristics of the three
water masses (Jan, in prep.). The KBC was taken on 01/05/1991 in the central Penghu Channel, the SCSSC was taken on 08/16/1999 off the southwestern Taiwan, and the CCC was taken on 03/21/2000 in the northwestern Taiwan Strait. Note that coincidently the chosen typical CCC is the station 6 of this study.
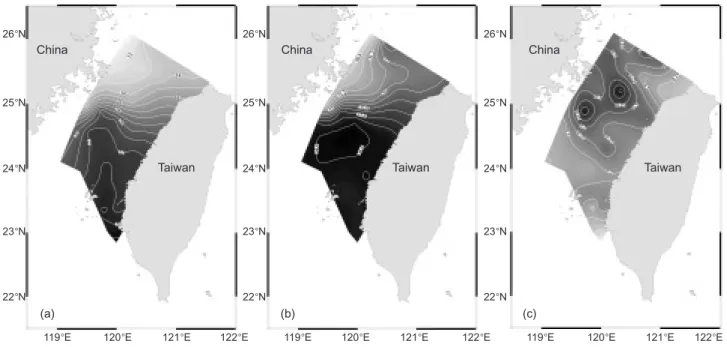
The distinctive properties of the CCC and KBC were clarified by observing surface water temperature and salinity. From contour plots of surface water temperature and salinity (Fig. 3a, b), one can observe the high-temperature and -salinity water of the KBC flowing over the Changyun Ridge (see Fig. 1) and penetrating into the northern Taiwan Strait. Dense contour lines of temperature and salinity south of the Haitan Is. indicate that a front had formed, and that the front resulted in a phytoplankton bloom represented by high fluores-cence values (Fig. 3c). Furthermore, profiles of temperature and salinity of the northern transect (Matsu-Cape Fuguei) showed a high-temperature and -salinity core at a distance 60~80 km from the western end, which indicated that the influence of the KBC had extended to the northern Strait (Fig.
4a, c). The extreme low-temperature and -salinity water along the China coast was the southward extension of the CCC and river runoff from Fujian Province (Fig. 4).
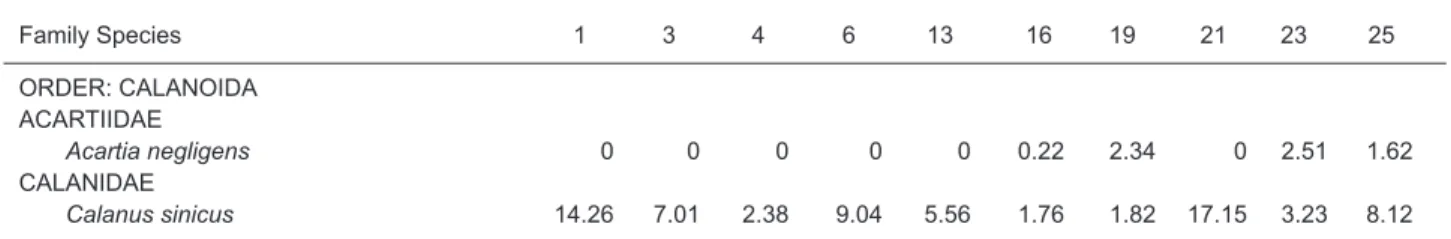
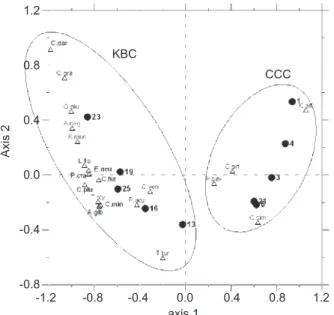
In our samples, 70 species of copepods were identified, belonging to 30 genera, 17 families, and 4 orders (only individuals identified to species were counted) (Table 1), and some clarification on cope-pod taxonomy is provided in the footnotes. Species richness and diversity were higher in the area influenced by the KBC than by the CCC (Fig. 5, Table 1). The correspondence analysis resulted in two station clusters through K-means classifica-tion, namely the CCC and KBC (Fig. 6), and the first 2 axes explained 78.11% of the variance. The CCC included stations 1, 3, 4, 6, and 21 with cor-responding species of Calanus sinicus, Corycaeus (Ditrichocorycaeus) affinis, Oithona similis, and Paracalanus parvus, while the KBC included sta-tions 13, 16, 19, 23, and 25 with corresponding species of Acartia negligens, Acrocalanus gracilis, Calocalanus gracilis, Cal. plumulosus, Cosmocalanus darwinii, Clausocalanus furcatus, Cla. minor, Euterpina acutifrons, Lucicutia flavicor-nis, Oithona plumifera, Oncaea venusta, Paracalanus aculeatus, Parvocalanus cras-sirostris, Subeucalanus mucronatus, Temora stylif-era, and T. turbinata. The indicator species of the two water masses with significant indicator values are shown in Table 2. Only one species, Corycaeus affinis, belonged to the CCC, and 16 species of the KBC were recognized as indicator species. Species with higher indicator values are more representative of the water mass, and a value of 100 indicates a perfect indicator. Two species, Calanus sinicus and Oithona similis, belonging to the CCC and one species, Clausocalanus arcuicornis, belonging to the KBC with marginal significance, are also listed in table 2. The average copepod abundance was 1368 individuals/m3in the CCC and 1296 individuals/m3
Table 1. Copepod species composition (%), species richness, diversity, and abundance, and environmental
information at each sampling station. (only individuals identified to species were included in the measure-ment of species richness and diversity)
Family Species 1 3 4 6 13 16 19 21 23 25 ORDER: CALANOIDA ACARTIIDAE Acartia negligens 0 0 0 0 0 0.22 2.34 0 2.51 1.62 CALANIDAE Calanus sinicus 14.26 7.01 2.38 9.04 5.56 1.76 1.82 17.15 3.23 8.12
Table 1. (Cont.) Family Species 1 3 4 6 13 16 19 21 23 25 Canthocalanus pauper 0 0 0 0 0.38 0.22 1.04 0 0.72 0.32 Cosmocalanus darwinii 0 0 0 0 0 0 0 0 2.87 0.65 Mesocalanus tenuicornis 0 0 0 0 0 0 0.26 0 0 0 Nannocalanus minor 0 0.19 0 0 0 0.22 0.26 0 0 0.32 Undinula vulgaris 0 0 0 0 0 0 0.26 0 0.72 0 CANDACIIDAE Candaciidae copepodid 0 0 0 0 0 0 0.26 0 0 0.32 CLAUSOCALANIDAE Clausocalanus arcuicornis 0 0.19 0 0 0 0.44 0.52 0 0.36 0.32 Clausocalanus furcatus 0 0 0 0 3.26 0 1.56 0 2.87 2.27 Clausocalanus mastigophorus 0 0 0 0 0 0.44 0 0 0.36 0 Clausocalanus minor 0 0 0 0 1.34 1.32 2.34 0 1.08 2.92 EUCALANIDAE Pareucalanus attenuatus 0 0 0 0 0 0 0 0 0.36 0.32 Rhincalanus rostrifrons1 0 0 0 0 0 0 0 0 0.72 0 Subeucalanus crassus 0 0 0 0 0 0 0 0 0.36 0.32 Subeucalanus mucronatus 0 0 0 0 0 0.88 1.04 0 1.79 0.65 Eucalanidae copepodid 0 0 0 0 0.38 0 0 0 0.72 3.57 EUCHAETIDAE Euchaeta concinna 0 0 0 0 0 0 0.26 0 0 0 Euchaeta plana 0 0.19 0 0.6 0 0.22 0 0 0 0 Euchaeta rimana2 0 0 0 0 0.19 0 1.04 0 1.08 0 Euchaeta spinosa 0 0 0 0 0 0 0 0 0 0.32 Euchaetidae copepodid 0.4 0.76 0.95 0.6 1.53 4.18 4.95 0 5.73 0.65 LUCICUTIIDAE Lucicutia flavicornis 0 0 0 0 0 1.1 0.78 0 0.72 0.32 Lucicutia gaussae 0 0 0 0 0 0 0 0 0.36 0 METRIDINIDAE Pleuromamma abdominalis 0 0 0 0 0 0 0 0 0 0.32 Pleuromamma gracilis 0 0 0 0 0 0.22 0 0 0 0 PARACALANIDAE Acrocalanus gibber 0 0 0 0 0.77 1.54 2.6 0 0.72 2.6 Acrocalanus gracilis 0 0 0 0 0 0 0 0 0 0.32 Calocalanus contractus 0 0 0 0 0 0 0.78 0 0 0 Calocalanus gracilis 0 0 0 0 0 0.22 1.04 0 2.87 0 Calocalanus monospinus 0 0 0 0 0 0 0 0 0.72 0 Calocalanus pavoninus 0 0 0 0 0 0 0 0 1.08 0 Calocalanus plumulosus 0 0 0 0 0 0.66 0.26 0 0.36 1.62 Calocalanus styliremis 0 0 0 0 0 0 0.26 0 1.08 0 Paracalanus aculeatus 0 0.19 0 0 3.45 6.81 5.99 7.56 7.17 7.47 Paracalanus parvus s.l. 29.32 55.3 59.62 64.76 42.53 70.33 40.89 32.56 24.73 41.56 Parvocalanus crassirostris 0 0 0 0 0.38 1.1 2.6 0 2.15 2.6 Parvocalanus sp. 0 0 0 0 0 0 0 0 0 0.32 PONTELLIDAE Pontellidae copepodid 0 0 0 0 0.19 0 0.26 0 0 0.65 SCOLECITRICHIDAE Scolecithricella longispinosa 0 0 0 0 0 0.22 0.26 0 0 0 Scolecithricella sp. 0 0 0 0.3 0 0 0 0 0 0 Scolecithrix danae 0 0 0 0 0 0.22 0.26 0 1.08 0.32 TEMORIDAE Temora discaudata 0 0 0 0 0 0 0 0 0.36 0 Temora stylifera3 0 0 0 0 1.34 0.44 1.3 0 1.08 4.55 Temora turbinata 0 0 0 0 0.57 0.22 0.78 0.87 0 0.32 ORDER: CYCLOPOIDA OITHONIDAE Oithona atlantica 0 0 0 0 0 0 0.26 0 0.36 0 Oithona attenuata 0 0 0 0 0.38 0 0 0 0 0
Table 1. (Cont.) Family Species 1 3 4 6 13 16 19 21 23 25 Oithona brevicornis 0 0 0 0 0.19 0 0 0 0 0 Oithona decipiens 0 0 0 0 0 0 0.26 0 0.36 0.65 Oithona fallax 0 0 0 0 0 0 0 0 0 1.3 Oithona fragilis 0 0 0 0 0 0 0 0 0.36 0 Oithona plumifera 0 0 0 0 0.19 0.22 6.77 0 12.9 2.92 Oithona similis 1.61 18.75 8.08 18.37 14.94 1.32 1.82 24.71 0 1.62 Oithona simplex 0 0 0 0 0.19 0 0 0 0 0 ORDER: HARPACTICOIDA CLYTEMNESTRIDAE Clytemnestra rostrata 0 0 0.24 0.3 0 0 0.26 0 0 0.32 EUTERPINIDAE Euterpina acutifrons 0 0 0 0 0.57 0 2.6 0 1.08 2.27 MIRACIIDAE Macrosetella gracilis 0 0 0.24 0 0.57 0 0 0 0 0 ORDER:POECILOSTOMATOIDA CORYCAEIDAE
Corycaeus (Agetus) flaccus 0 0 0 0 0 0 0.26 0 0 0
Corycaeus (Corycaeus) speciosus 0 0 0 0 0 0 0 0 1.08 0
Corycaeus (Ditrichocorycaeus) affinis 54.02 16.67 28.27 3.92 2.3 0 1.3 16.86 0 0
Corycaeus (Ditrichocorycaeus) dahli 0 0 0 0 0.77 0 0 0 0.36 0.32
Corycaeus (Ditrichocorycaeus) erythraeus 0 0 0 0 0 0 0.52 0 0 0
Corycaeus (Ditrichocorycaeus) subtilis 0 0 0 0 0 0 0 0 0.72 0
Corycaeus (Onychocorycaeus) pacificus 0 0 0 0 0.96 0.22 0 0 0 0.32
Corycaeus (Urochocorycaeus) furcifer 0 0 0 0 0 0 0 0 0.36 0
Farranula gibbula 0 0 0 0 0.38 0 0 0 2.15 0 Farranula rostrata 0 0 0 0 0.19 0 0 0 0 0 Corycaeidae copepodid 0 0 0 0 8.81 2.2 1.56 0 1.79 1.62 ONCAEIDAE Oncaea media 0 0.19 0 0 0.19 0 0 0 1.08 0 Oncaea mediterranea 0 0 0 0 0.96 0.22 0.78 0 0.72 0 Oncaea venusta 0.4 0.38 0 2.11 6.32 2.64 5.73 0.29 6.09 2.6 Oncaea sp1 0 0 0 0 0.19 0 0 0 0 0 Oncaea sp2 0 0 0.24 0 0 0 0 0 0 0 Triconia conifera4 0 0.19 0 0 0 0 0.78 0 0 0 Triconia minuta 0 0 0 0 0 0.22 0 0 0 0 Triconia similis 0 0 0 0 0 0 0.78 0 0 0 SAPPHIRINIDAE Copilia mirabilis 0 0 0 0 0 0 0 0 0 0.32 Sapphirina gemma 0 0 0 0 0 0 0 0 0.36 0 Sapphirina ovatolanceolata 0 0 0 0 0 0 0 0 0.36 0 No. of species 5 11 6 7 26 27 39 7 43 33 Shannon,s diversity 1.059 1.22 0.986 1.067 1.933 1.212 2.443 1.567 2.878 2.295 Density (ind./m3) 1119 1153 1719 855 1246 2123 1159 1497 1351 960 Depth (m) 70 80 72 41 56 50 46 35 60 31
Sea surface temperature (
°
C) 14.71 13.56 13.66 10.55 13.67 18.70 21.31 13.08 22.52 21.84 Sea surface salinity (psu) 32.63 32.11 32.27 29.56 31.22 33.43 34.35 30.42 34.42 34.40 1. Bowman (1971) recognized the two forms of Rhincalanus cornutus, f. typica from the Indian and Pacific oceans, and f. atlanticafrom the Atlantic Ocean as two species, based on the difference in the structure of the 5th legs as well as the relative length of dis-tal segments of the 1st antenna of female. He retained the species name for the Atlantic form and reinsdis-talled the Indopacific form as Rhincalanus rostrifrons, which was already recognized by Dana (1853).
2. Bradford (1974) found the so-called Euchaeta marina.
3. According to Fleminger and Hulsemann (1973), Temora stylifera is endemic to the Atlantic Ocean. However, the specimens of T.
stylifera in the current study are consistent with those described in Chen and Zhang (1965). Whether T. stylifera also distribute in
the Pacific Ocean or the species described in Chen and Zhang (1965) is another unknown new species is a question that required further study.
in the KBC. There was no significant difference in abundance between these two water masses (U = 10.000, p = 0.602). The species richness and diversity were higher in the KBC than those in the CCC (U = 0.000, p = 0.009 and U = 2.000, p = 0.028 respectively).
DISCUSSION
Intrusions of the Kuroshio Branch Current (KBC) into the northern Taiwan Strait during the waning of the northeastern monsoon have been described by Jan et al. (1998 2002). According to their observations from 1985 to 1992 and simula-tion studies, they found a recurring pattern of the KBC intruding into the retracting China Coastal Current (CCC) during spring, which was generally defined as occurring in Apr. to May. Although the present study was conducted in late Mar., the observed hydrography is consistent with the results of Jan et al. (2002), i.e., the presence of the high-temperature and -salinity KBC intruding into the low-temperature and -salinity CCC (Figs. 3, 4). The front induced by the interaction of the KBC and CCC is marked by the temperature of around 18 ± 2
°
C and salinity of around 33 ± 0.5 psu(Fig. 3a, b). The temperature and salinity proper-ties showed a strong contrast between the CCC
and KBC (Figs. 2, 3a, b). The hydrographic pat-tern was clear, but the temperature and salinity values should be treated with caution since long-term variations in oceanographic properties in the Taiwan Strait are still under examination (Jan,
Fig. 2. Temperature-salinity diagram of copepod sampling
sta-tions. Station 41 represents the temperature-salinity curve in the Penghu Channel; KBC, Kuroshio Branch Current; SCSSC, South China Sea Surface Current; CCC, China Coastal Current.
Table 2. Indicator species of China Coastal Water and Kuroshio Branch Water. (Ind Val, indicator value;
p-value, probability of Type-I error through Monte-Carlo test; *, species with only marginal significance)
China Coastal water Kuroshio Branch Water
Species Ind Val p-value Species Ind Val p-value
Corycaeus (Ditrichocorycaeus) affinis 97 0.004 Acrocalanus gibber 100 0.004
Canthocalanus paupur 100 0.004
*Calanus sinicus 70.4 0.073 Clausocalanus minor 100 0.004
*Oithona similis 74.9 0.082 Oithona plumifera 100 0.004
Parvocalanus crassirostris 100 0.004 Temora stylifera 100 0.004 Oncaea venusta 90 0.004 Paracalanus aculeatus 83 0.032 Acartia negligens 80 0.038 Calocalanus plumulosus 80 0.038 Clausocalanus furcatus 80 0.045 Euterpina acutifrons 80 0.045 Lucicutia flavicornis 80 0.038 Oncaea mediterranea 80 0.044 Scolecithrix danae 80 0.038 Subeucalanus mucronatus 80 0.038 *Clausocalanus arcuicornis 70 0.081 Salinity (psu) 29 31 33 35 30 28 26 24 22 20 18 16 14 12 10 SCSSC KBC CCC Temperature (
°
C)pers. comm.).
No significant differences in copepod abun-dance between the areas affected by the CCC and KBC were detected, which is consistent with the studies of nutrient fluxes (Liu et al. 2000, Chung et al. 2001). The front induced south of the Haitan Island resulted in a phytoplankton bloom (Fig. 3c), but no corresponding spatial pattern of zooplank-ton was observed. However, we have a limited sample size and do not have in situ rate measure-ments of phytoplankton and zooplankton. Therefore, no attempt was made to explain the trophic links among nutrients, phytoplankton, and zooplankton.
The species richness and diversity of cope-pods were higher in the area affected by the KBC than that by the CCC (Fig. 5). The Kuroshio is an oceanic current with a more-stable environment for plankton compared with China coastal water, which is subject to disturbance and nutrient enrich-ment from the land. The fluctuating coastal envi-ronment might be the reason why we only saw a few coastal and neritic species in the CCC (Table 1). The study by Shih and Chiu (1998) in the water north of Taiwan also showed a similar trend that copepod diversity decreases from the Kuroshio area toward the China coastal area. A similar coastal-oceanic gradient of copepod diver-sity was also observed in the waters off Rio de Janeiro, southeast Brazil (Lopes et al. 1999). On the other hand, the intrusion of the KBC brought warm-water oceanic species into the Taiwan Strait,
added to the coastal species that can tolerate high temperature and salinity, and resulted in a com-bined assemblage with a higher diversity. Enrichment in zooplankton diversity caused by mixing the Kuroshio assemblage with the coastal assemblage was also observed in the coastal area south of Japan (Noda et al. 1998). A long-term study is required to check if the lower diversity in the CCC and higher diversity in the KBC are invari-ant biological properties of these two water mass-es. It will be interesting to observe the long-term dynamics of the copepod diversity in the frontal area where the CCC meets the KBC.
The presence of the two station clusters, namely the CCC and KBC, based on the copepod compositions is consistent with the results derived from hydrographic data. According to the records of Chen and Zhang (1965) and Chen et al. (1974), among the four corresponding species of the CCC, Calanus sinicus is abundant in the coastal and neritic waters of the Yellow Sea and East China Sea, Corycaeus (Ditrichocorycaeus) affinis and Oithona similis are abundant in the coastal waters of the Yellow Sea, and Paracalanus parvus is abundant in coastal waters off China. Of the 16 corresponding species of the KBC, Acartia negli-gens, Calocalanus gracilis, Calocalanus plumulo-sus, Cosmocalanus darwinii, Lucicutia flavicornis, and Subeucalanus mucronatus are oceanic species; Acrocalanus gibber, Clausocalanus furca-tus, Clausocalanus minor, Oithona plumifera, Oncaea venusta, Paracalanus aculeatus, T.
stylif-Fig. 3. Contour plots of surface water (a) temperature, (b) salinity, and (c) fluorescence.
China China China
Taiwan (a) (b) (c) Taiwan Taiwan 26°N 25°N 24°N 23°N 22°N 26°N 25°N 24°N 23°N 22°N 26°N 25°N 24°N 23°N 22°N 119°E 120°E 121°E 122°E 119°E 120°E 121°E 122°E 119°E 120°E 121°E 122°E
era, and T. turbinata are neritic species; and Euterpina acutifrons and Parvocalanus cras-sirostris are coastal species. They are all confined to warm waters (Chen and Zhang 1965, Chen et al. 1974, Chen 1992). From our study, we believe that Calanus sinicus, Corycaeus affinis, and Oithona similis followed the CCC into the Taiwan Strait from the north, while warm-water oceanic species accompanied the KBC into the Strait from the south and mixed with coastal and neritic species in the northern Strait. Paracalanus parvus was the most abundant species and had a broad distribution in this study.
In conclusion, the CCC contained only coastal/neritic cold-water species, but the KBC consisted of coastal, neritic, and oceanic warm-water species. The copepod community in the KBC is a mixture of the Kuroshio assemblage and coastal/neritic assemblage in the Taiwan Strait, which reflects the high diversity mentioned above. Using plankton assemblages to characterize the hydrography is important for understanding marine ecosystems. For example, Boucher et al. (1987) showed the spatial distribution of zooplankton in relation to the hydrography in the Ligurian Sea
front; Kouwenberg (1994) examined the copepod distribution in relation to the spatial structure in the northwestern Mediterranean; Gowen et al. (1998) used plankton distribution to characterize the frontal structure in the Irish sea; and Lopes et al. (1999) studied the distribution pattern of copepods related to the advection of South Atlantic Central Water southeast of Brazil.
Besides showing distinctive community struc-tures in the CCC and KBC, we provide indicator species of each water mass. Among the 16 good indicators for the KBC, three are not included in the 20 most abundant species. These three, Canthocalanus pauper, Oncaea mediterranea, and Scolecithrix danae, are also warm-water species according to the records of Chen and Zhang (1965) and Chen et al. (1974). We have to emphasize that the copepod community in the KBC differs from that of the main Kuroshio Current by having a mixture of Kuroshio oceanic species and coastal/neritic warm-water species. By extracting the indicator species, we hope to pro-vide useful information for monitoring Taiwan Strait ecosystems. Some species that are not abundant but appear frequently could be important targets
Fig. 4. Vertical sections of temperature of the (a) northern (Matsu-Cape Fuguei) and (b) central (Wuchiu-Taichung) transects and of
salinity of the (c) northern (Matsu-Cape Fuguei) and (d) central (Wuchiu-Taichung) transects. Triangles indicate sampling stations. Relative distance (km) 120 100 80 60 40 20 0 140 160 120 100 80 60 40 20 0 140 160 Relative distance (km) (a) (b) (c) (d) Relative distance (km) 100 120 80 60 40 20 0 120 100 80 60 40 20 0 Relative distance (km) 0 -20 -40 -60 -80 -100 0 -20 -40 -60 -80 -100 0 -20 -40 -60 -80 -100 0 -20 -40 -60 -80 -100 Depth (m) Depth (m) Depth (m) Depth (m)
for long-term observation. The use of biological indicators to label water masses is as effective as physical or chemical methods in many cases, and it also provides useful information for observing ecosystem dynamics, especially when plankters are sensitive to environmental changes (Haury and Pieper 1988). Also, variations in zooplankton com-position and abundance are widely used as prox-ies to observe long-term changes in marine ecosystems (Rebstock 2001 2002, Beaugrand et
al. 2002, Lindley and Reid 2002). Our future plan is to continually monitor this ecosystem in an attempt to understand seasonal and inter-annual variations.
Acknowledgments: We thank the crew of the
Ocean Research I for their help in the collection of plankton and oceanographic data. We also thank Dr. S. Jan of the Institute of Physical Ocean-ography, National Sun Yat-Sen Univ., Kaohsiung, Taiwan for sharing his cruise time with us and pro-viding insightful advise on the hydrography of the Taiwan Strait and Dr. M. D. Ohman for his com-ments on this manuscript. Thanks are also due to the constructive criticism of this manuscript by the three anonymous reviewers. We would like to extend our acknowledgement to Ms. K.Z. Chang and Ms. C.C. Chen in the Economic Fish Labora-tory, Department of Zoology, National Taiwan Univ., Taipei, Taiwan for their aid in the laboratory. This work was partially financed by two research grants from the National Research Council, R.O.C. (NSC89-2611-M002-008 to TSC and NSC89-2811-B056-0002 to CtS).
REFERENCES
Beaugrand G, PC Reid, F Ibañez, JA Lindley, M Edwards. 2002. Reorganization of North Atlantic marine copepod biodiversity and climate. Science 296: 1692-1694. Böttger-Schnack R. 1999. Taxonomy of Oncaeidae
(Co-pepoda, Poecilostomatoida) from the Red Sea. I. 11 species of Triconia gen. nov. and a redescription of T.
similis (Sars) from Norwegian waters. Mitt. Hamb. Zool.
Mus. Inst. 96: 37-128.
Boucher J, F Ibañez, L Prieur. 1987. Daily and seasonal varia-tions in the spatial distribution of zooplankton populavaria-tions in relation to the physical structure in the Ligurian Sea front. J. Mar. Res. 45: 133-173.
Bowman TE. 1971. The distribution of calanoid copepods off the southeastern United States between Cape Hatteras and southern Florida. Smiths. Contr. Zool. 96: 1-58. Bradford JM. 1974. Euchaeta marina (Prestandrea)
(Copepoda, Calanoida) and two closely related new species from the Pacific. Pac. Sci. 28: 159-169.
Chen QC. 1992. Zooplankton of China seas (1). Beijing: Science Press.
Chen QC, SZ Zhang. 1965. The planktonic copepods of the Yellow Sea and the East China Sea. 1. Calanoida. Stud. Mar. Sinica 7: 20-133. (in Chinese with English abstract) Chen QC, SZ Zhang, CS Zhu. 1974. On planktonic copepods
of the Yellow Sea and the East China Sea. 2. Cyclopoida and Harpacticoida. Stud. Mar. Sinica 9: 27-100. (in Chinese with English abstract)
Chiu TS. 1992. An exploration on ichthyoplankton assemblage in the water off southwestern Taiwan, with an inference on the role of Kuroshio on the distribution of ichthyoplankton. J. Fish. Soc. Taiwan 19: 1-12.
Fig. 5. Scatter plots of copepod species (a) richness (number
of species) and (b) diversity (Shannon,s index) at each sam-pling station. (a) China 40 20 10 5 Taiwan 26°N 25°N 24°N 23°N 22°N 119°E 120°E 121°E 122°E (b) China 3 2 1 Taiwan 26°N 25°N 24°N 23°N 22°N 119°E 120°E 121°E 122°E
Fig. 6. Ordination diagram derived from correspondence
analysis based on the relative abundance of the 20 most abun-dant species of copepods, showing station associations and corresponding species. The ellipses indicate two station clus-ters determined by K-means classification. The first letter of the genus and the first three letters of the species name were used as abbreviations in the plot.
Axis 2 1.2 0.8 0.4 0.0 -0.4 -0.8 axis 1 KBC CCC -1.2 -0.8 -0.4 0.0 0.4 0.8 1.2
Chiu TS, KZ Chang. 1994. Comparison of ichthyoplankton fauna in northern Taiwan Strait during winter and spring. Acta Zool. Taiwanica 5: 23-31.
Chiu TS, KZ Chang. 1995. Diurnal cycling in vertical distribu-tion of ichthyoplankton at a fixed stadistribu-tion off northwestern Taiwan. Zool. Stud. 34: 183-192.
Chiu TS, CS Chen. 1997. Distribution of ichthyoplankton in the waters off southwestern Taiwan. Proc. Nat. Sci. Council (ROC) 21: 79-89.
Chung SW, S Jan, KK Liu. 2001. Nutrient fluxes through the Taiwan Strait in spring and summer 1999. J. Oceanogr.
57: 47-53.
Dana JD. 1853. Crustacea. Part II. In United States Exploring Expedition during the years 1838, 1839, 1840, 1841, 1842 under the Command of Charles Wilkes, U.S.N. 13: 689-1618.
Dufrene M, P Legendre. 1997. Species assemblages and indi-cator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67: 345-366.
Fleminger A, K Hulsemann. 1973. Relationship of Indian Ocean epiplanktonic Calanoids to the world oceans. In Zeitzschel B, ed. The biology of the Indian Ocean. New York: Springer-Verlag Press.
Gowen RJ, R Raine, M Dickey-Collas, M White. 1998. Plankton distributions in relation to physical oceanograph-ic features on the southern Malin Shelf, August 1996. ICES J. Mar. Sci. 55: 1095-1111.
Haury LR, RE Pieper. 1988. Zooplankton: scales of biological and physical events. In DF Soule, GS Kleppel, eds. Marine organisms as indictors. New York: Springer-Verlag Press.
Hsieh CH, TS Chiu. 1998. Copepod abundance and species composition of Tanshui River estuary and adjacent waters. Acta Zool. Taiwanica 9: 1-9.
Jan S, CS Chern, J Wang. 1998. A numerical study of cur-rents in the Taiwan Strait during winter. Terre. Atmosph. Oceanic Sci. 9: 615-642.
Jan S, Wang J, CS Chern, SY Chao. 2002. Seasonal variation of the circulation in the Taiwan Strait. J. Mar. Syst. 35: 249-268.
Kouwenberg JHM. 1994. Copepod distribution in relation to seasonal hydrographics and spatial structure in the north-western Mediterranean (Golfe du Lion). Estuar. Coast. Shelf. S. 38: 69-90.
Krebs CJ. 1989. Ecological methodology. New York: Harper and Row.
Lindley JA , PC Reid. 2002. Variations in the abundance of
Centropages typicus and Calanus helgolandicus in the
North Sea: deviations from close relationships with tem-perature. Mar. Biol. 141: 153-165.
Liu KK, TY Tang, GC Cong, LY Chen, FK Shiah. 2000. Cross-shelf and along-Cross-shelf nutrient fluxes derived from flow fields and chemical hydrography observed in the southern East China Sea off northern Taiwan. Cont. Shelf Res. 20: 493-523.
Lopes, RM, FP Brandini, SA Gaeta. 1999. Distribution pat-terns of epipelagic copepods off Rio de Janeiro (SE Brazil) in summer 1991/1992 and winter 1992. Hydrobiologia 411: 161-174.
Noda, M, I Ikeda, S Ueno, H Hashimoto, K Gushima. 1998. Enrichment of coastal zooplankton communities by drift-ing zooplankton patches from the Kuroshio front. Mar. Ecol.-Prog. Ser. 176: 55-65.
Pielou EC. 1984. The interpretation of ecological data. New York: J Wiley.
Poulet SA, R Williams. 1991. Characteristics and properties of copepods affecting the recruitment of fish larvae. Bull. Plankton Soc. Jpn. Spec. 271-290.
Rebstock GA. 2001. Long-term stability of species composi-tion in calanoid copepods off southern California. Mar. Ecol.-Prog. Ser. 215: 213-224.
Rebstock GA. 2002. Climatic regime shifts and decadal-scale variability in calanoid copepod populations off southern California. Glob. Change Biol. 8: 71-89.
Runge JA. 1988. Should we expect a relationship between pri-mary production and fisheries? The role of copepod dynamics as a filter of trophic variability. Hydrobiologia
167/168: 61-71.
Shih Ct, TS Chiu. 1998. Copepod diversity in the water mass-es of the southern East China Sea north of Taiwan. J. Marine Syst. 15: 533-542.
Tan TH. 1967. On the distribution of copepods in waters sur-rounding Taiwan. Rep. Inst. Fish. Biol. Min. Econ. Aff. Natl. Taiwan Univ. 2: 14-20.
Tzeng WN, YT Wang. 1992. Structure, composition and sea-sonal dynamics of the larval and juvenile fish community in the mangrove estuary of Tanshui River, Taiwan. Mar. Biol. 113: 481-490.
Tzeng WN, YT Wang. 1993. Hydrography and distribution dynamics of larval and juvenile fishes in the coastal waters of the Tanshui River estuary, Taiwan, with refer-ence to estuarine larval transport. Mar. Biol. 116: 205-217.
Wang J, CS Chern. 1988. On the Kuroshio branch in the Taiwan Strait during wintertime. Prog. Oceanogr. 21: 469-491.
Wilkinson L. 1990. SYSTAT: the system for statistics. Evaston, IL: Systat, Inc.
Williams R, DVP Conway, HG Hunt. 1994. The role of cope-pods in the planktonic ecosystems of mixed and stratified waters of the European shelf seas. Hydrobiologia
292/293: 521-230.
Zheng Z, SJ Li, S Li, BY Chen. 1982. On the distribution of planktonic copepods in Taiwan strait. Taiwan Strait 1: 69-79. (in Chinese with English abstract)
Zhu CS, JZ Wu, YS Lin, M Su, CJ Huang, QB Hu. 1988. The species composition, distribution and abundance of plank-ton. In Fujian Institute of Oceanography, eds. A compre-hensive oceanographic survey of the central and northern part of Taiwan Strait. Beijing: Science Press. (in Chinese)