國立臺東大學應用科學系 碩士論文
指導教授:廖尉岑 博士
探討奈米銀對渦蟲運動行為模式的影響 The effects of nano-silver on the
movement patterns of planarians
研究生:張順龍 撰
中華民國 106 年 7 月
國立臺東大學應用科學系 碩士論文
指導教授:廖尉岑 博士
探討奈米銀對渦蟲運動行為模式的影響 The effects of nano-silver on the
movement patterns of planarians
研究生:張順龍 撰
中華民國 106 年 7 月
i
ii
iii
誌謝
時光荏苒,從學士到碩士,在臺東大學已經待了十年有餘,一路上 要感謝的貴人太多,無法一一答謝,深感歉意。
首先,謝謝我的家人-張爸爸、張媽媽,身為兒子的我,從大一開 始就是半年才回家一次,每年回家的天數總和沒有超過一個月,未能 待在兩老身旁盡孝道,實為不孝,幸得爸媽的支持、鼓勵與包容,讓 我得以無後顧之憂地完成學業,同時拓展我的夢想藍圖,謝謝爸爸、
謝謝媽媽!
接著要感謝我的老師-廖尉岑教授,從大二開始,我們一起在艱難 的實驗室環境中奮鬥,這句話應該不誇張,沒有貴儀的實驗室,我們 做的都是臺東在地的實驗材料,或者是低等廉價的生物體,但是實驗 的價值不在研究材料的價格,我們從豐年蝦、溫泉菌、擬球藻、黏菌、…,
一路研究到本篇內容的主角-渦蟲,或許我們的成果不多,但是在實驗 裡獲得的樂趣可不亞於拿到任何獎項,研究的本質在於設計問題去探 索真相,我會銘記老師的這句話,謝謝老師!
隱+1,這是時下年輕人的用語,無法在此時此刻公布妳的相關資訊,
相信在本篇論文完成後,老夫老妻的生活就快要開始了,謝謝妳!
iv
鏡頭轉到應用科學系,我從應科系的前身自然科學教育學系開始就 讀學士班,以前都是培育自然科的師資生,入學分數不算太高,現在 的學弟妹就不一樣了,每個專業的知識都好厲害,我真的無法比擬,
系上的老師們更猛,從教基礎學科轉型指導專業學科,因材施教的能 力可見一斑,歡迎大家來唸應用科學系,同時也謝謝應用科學系這些 年來的照顧與指教!
謝謝臺東大學附小,在我求學期間,讓我擔任了育樂營教練、代課 老師、代理教師、科展指導老師,獲得的薪資不是重點,重點是在其 中的經驗與體悟,在附小的日子,我明白了自己專業能力的不足與侷 限,也體認到自己無法往專業領域去發展,透過指導孩子的機會,我 發現自己喜歡從事推廣科學教育,漸漸地將重心轉移到科普的推展,
而碩士畢業後會繼續在臺東大學附小服務,請附小師生多多指教!
最後感謝胡焯淳教授與胡安仁老師,在百忙之中前來指導我的口試,
兩位老師所交代的那最後一哩路,我會好好走完!
「因為感謝的人太多了,那就謝天吧」-陳之藩先生《在春風裡》。 忘情理工亦不忘文學,總而言之,謝謝天!
張順龍 2017.07.01
v
探討奈米銀對渦蟲運動行為模式的影響
張 順 龍
國 立 臺 東 大 學 應 用 科 學 系
摘 要
本 研 究 主 要 是 檢 測 奈 米 銀 對 再 生 渦 蟲 的 毒 性 , 在 奈 米 銀 處 理 後,渦 蟲 的 運 動 數 據 蒐 集 並 分 析,我 們 設 計 一 個 標 準 化 圓 形 軌 道 平 台 來 定 量 及 分 析 渦 蟲 的 運 動 型 態 。
銀 奈 米 粒 子 處 理 後 的 渦 蟲 在 探 索 行 為 時 間 會 增 加 , 而 經 過 橫 切 手 術 的 渦 蟲 也 會 有 增 加 探 索 時 間 的 狀 況 。
觀 察 渦 蟲 在 圓 形 軌 道 中 的 表 現 , 對 照 組 是 以 逆 時 針 方 向 行 進 為 主,橫 切 手 術 會 導 致 順 時 針 方 向 行 進 變 多,經 過 銀 奈 米 粒 子 處 理 後 可 以 恢 復 以 逆 時 針 為 主 的 行 進 模 式 。
我 們 發 展 出 一 套 新 的 方 法 來 測 試 渦 蟲 的 運 動 行 為 , 利 用 圓 形 軌 道 讓 渦 蟲 在 有 限 的 空 間 中 進 行 運 動,這 方 法 我 們 命 名 為 pCOLMV(Planarian Circular Orbit Loco Motion Velocity),
預 期 對 再 生 醫 學 的 藥 物 開 發 有 突 破 性 的 貢 獻 。
關 鍵 詞 : 渦 蟲 、 奈 米 銀 、 運 動 行 為 模 式
vi
The effects of nano-silver on the movement patterns of planarians
Shun-Lung Chang
Abstract
The purpose of this study is to examine the toxicity of silver nanoparticles (AgNPs) in the regenerating planarians. After the AgNPs treatments, the planarian locomotion data were collected and analyzed. A standardized circular pathways platform was developed to quantify and profiling the locomotion patterns of planarians.
The standardized circular pathways platform was designed to force planarian moving forward, however, experiments showed that the platform still allow planarian moving clockwise or
counterclockwise. We defined the “looking around” behavior as explore time (ET). The explore time was increased both the AgNPs treatments groups and planarian cutting group.
vii
The performance of planarians in circular pathways shown that the moving direction of the control group was counterclockwise , however, the planarian cutting group changed the direction clockwise. After the AgNPs treatments, the planarian moving direction was restored to counterclockwise-oriented mode .
We developed a novel approach for the locomotion testing of planarian using circular orbit in a limited space. The method, we named it “Planarian Circular Orbit LocoMotion Velocity (pCOLMV)”, can be quite useful in biomedical research,
especially in the drug development for regeneration medicine.
Keywords: planarian、nano-silver、movement patterns
1
目次
i. 口試委員審定書ii. 授權書 iii. 誌謝辭 v. 中文摘要 vi. 英文摘要
圖目次...4
表目次...4
第一章 前言 壹、渦蟲...5
貳、奈米銀粒子應用 ...6
一、傷口敷料 ...6
二、抗菌劑 ...7
參、奈米銀合成...7
肆、奈米銀對渦蟲的毒性...7
伍、研究目的...8
一、奈米銀對渦蟲的毒性...9
二、渦蟲運動速度...9
三、渦蟲圓形軌道運動速度...9
2
第二章 材料與方法
壹、藥品與試劑...10
一、藥品...10
二、奈米銀溶液配製...10
三、渦蟲來源...10
四、渦蟲準備工作...11
貳、儀器...13
參、實驗方法...13
一、奈米銀的合成與對渦蟲的毒性...13
二、渦蟲的預備實驗...16
三、運動行為資料蒐集...18
第三章、結果與討論 壹、渦蟲進行移動前的探索時間...20
一、橫切手術會使探索時間增加...20
二、奈米銀溶液處理會使探索時間增加...20
三、奈米銀溶液處理的影響比橫切手術明顯...21
四、奈米銀溶液處理與橫切手術有加成的效果...21
3
貳、渦蟲在方向上的表現...22
一、渦蟲移動時以逆時針方向為主...22
二、橫切手術會造成渦蟲失去方向感...23
三、奈米銀粒子可以促進渦蟲恢復方向感...23
參、渦蟲實驗的一些新發現和設計...24
一、雙頭渦蟲...24
二、紅色渦蟲...24
三、3D 列印圓形軌道...25
第四章 結論與未來展望...26
第五章 參考文獻...27
附錄...33
4
圖目次
圖 1 渦蟲實驗所需器材...12
圖 2 在實驗室飼養渦蟲情形...12
圖 3 本研究所使用的奈米銀...13
圖 4 用 100%奈米銀處理後,渦蟲的型態變化...15
圖 5 色光實驗組...16
圖 6 pLMV 實驗...17
圖 7 圓形軌道實驗...17
圖 8 探索時間...21
圖 9 雙頭渦蟲在顯微鏡底下的影像...24
圖 10 顯微鏡下,渦蟲體色改變的樣子...24
圖 11 3D 列印圓形軌道實驗...25
表目次 表 1 渦蟲在順時針方向移動與逆時針方向移動之比例...22
5
第一章 前言
壹、渦蟲渦蟲是一種神奇的生物,不僅具備中樞神經系統,也和所有脊椎 動物一樣有雙側對稱性和三軸[1-3]。大多數研究用的渦蟲體長約一公 分,不占空間又好操作,很早便成為實驗室中一個常用的模式動物。
從達爾文到二十世紀,有各式各樣的渦蟲研究[1-12],有的研究運動 [1]、有的研究毒性[2]、有的切割以後研究其再生能力[3-4]、有的研 究電對其細胞生長的影響等等[5],或是研究渦蟲的基本的特性(趨光 性、趨化性等等),進而研發渦蟲在科技上應用的可能性[6-12]。
近年來由於幹細胞與組織工程的研究興起,從分子到組織或從離 子到神經,渦蟲以絕佳的模式生物態勢,挺立於再生醫學的研究與應 用上[13-21],研究的面向十分廣泛。有些研究著重在渦蟲的運動速度 (planarian locomotion velocity ---pLMV),渦蟲運動速度(pLMV)是 一種很容易量化的行為反應,主要是在 Raffa 教授實驗室所開發出來,
應用在相當多的藥物研究上,甚至包括複方的處理[22-31]。pLMV 是基 於計數的格線交叉,經過處理的渦蟲單獨放入一個透明塑膠培養皿,
在格線版進行簡單的疊加計數,雖然步驟容易卻是十分有效的方法,
該方法被發現影響數值的原因是渦蟲對劑量依存性(與濃度有關)
[25-28]。
6
有一部份研究是在說明生物的探索時間[33-35],對生物而言,當 進入一個陌生的環境時,會對該環境有探索的行為,進而獲取訊息、
了解狀況,方能前行。而腦部掌控記憶、認知、判斷,無論經過化學 性或物理性的刺激,這些刺激對腦部的影響可以反應在探索行為上 [36-38]。
在型態觀察的部分,我們習慣用英文字母的形狀去定義生物體的 型態,在渦蟲接受某些刺激(例如:用塑膠吸管去吸牠)或致死性的刺 激時,會產生 C 型態(C Shape)[38]。
貳、奈米銀粒子應用 一、傷口敷料
奈米銀粒子在醫療領域被開發出許多應用,像是在床單、枕套、
醫療用止血紗布中加入奈米銀,除了可以防止細菌滋生和疾病傳染,
更可以降低病人在手術前後被病菌感染的風險[39-40]。
此外,將奈米銀加入人工皮膚敷料中,可以保護燒燙傷病患的 傷口,對長期需要敷料的病患而言,能夠有效避免細菌感染的情況。
近年來,相關研究已經發展到人體的植入性材料。例如,在固 定人工關節的骨水泥中加入奈米銀來取代抗生素,可以抑制細菌的 生長,避免手術後感染[41-42]。
7
二、抗菌劑
自古以來,銀做為毒物的偵測及抗菌劑,透過氧化破壞微生物 蛋白質氫硫官能基,而達到抗菌的效果。不同於其他抗菌劑,且奈 米銀具有無刺激性、無過敏性、不受酸鹼值影響等優點[39,42]。
參、奈米銀合成
許多奈米材料具備獨特的表面電漿共振(SPR)現象[43-45],金 屬奈米顆粒(尤其是銀和金),藉著有效的散射和吸收光的共振,隨著 形狀及顆粒大小的不同,呈現不同的色變範圍,例如奈米銀在可見光 光譜 400nm 至 700nm 波長之間[46-47],如何利用金屬奈米這種肉眼就 可以檢測的色變性質,開發新式生物化學感測器,是一個熱門的研究 方向[46-49]。
肆、奈米銀對渦蟲的毒性
利用模式生物來測試新開發奈米材料在生物體內的毒性,也是近 年來受到重視的研究方向[50-53]。本實驗室過去以多重模式生物,從 溫泉菌、擬球藻到斑馬魚,探討奈米金屬的毒性。現在進行的是鹵蟲 和渦蟲的奈米毒性測試[54],鹵蟲(Artemia)又稱為豐年蝦,其無節幼 蟲是極佳的餵食養殖魚蝦貝類的活體餌料。現在常用來當奈米粒子的 細胞毒性的評估模式生物[3]。鹵蟲和渦蟲(Planarian)在實驗上都有 不易固定,難以觀察的問題[52]。
8
其實結合透明無毒的聚二甲基矽氧烷(Polydimethylsiloxane,
PDMS),以薄膜的型態包裹模式生物,利用其獨特的光學性質來進行分 析診斷,是一個有效的策略。我們可以調控奈米顆粒製劑,使其具有 吸收和散射的光的特定波長,以產生客製化的複合材料,符合我們的 需求。
不同的色光誘導,或是以不同的溫度,或是不同的化學物質處理,
都會讓奈米銀產生色變,選擇其中具有高再現性的過程,可以發展低 成本、快速的偵測分析平台。在我們實驗室,結合微型光譜儀及自製 組件,已經完成一套利用奈米銀色變為基礎所建置的整合偵測平台,
提供一個高效能,快速,而且成本低的分析方法。
四種情況依探索時間進行排序:切 50%>沒切 50%>切 0%>沒切 0%。
透過排序,我們可以發現,不論有沒有橫切手術,奈米銀溶液處理後 的渦蟲在平均探索時間排在前兩名,分別是 6.6 秒和 6.3 秒,這樣的 結果顯示,奈米銀溶液對渦蟲的運動行為影響比橫切手術更加明顯。
9
伍、研究目的
一、奈米銀對渦蟲的毒性
從文獻中,可以知道奈米銀應用在傷口敷料上有長期抗菌的效 果,而在其他渦蟲的文獻中,目前只有極少數文獻對於奈米銀與渦 蟲之間的研究[54],這讓我們想要檢測奈米銀對渦蟲的毒性,了解 奈米銀對渦蟲再生的影響。
二、渦蟲運動速度(Planarian LocoMotion Velocity,pLMV) 無論是在文獻探討或是一般生理生化反應的數據收集,都有針 對生物體用藥後的運動數據進行分析,而本研究也不例外。根據文 獻內容,使用格線交叉的方式進行計數,並透過實驗室現有的設備 開發方法進行分析,了解渦蟲經過奈米銀的處理後所產生的變化。
三、渦蟲圓形軌道運動速度(Planarian Circular Orbit LocoMotion Velocity,pCOLMV)
由於文獻中的實驗是以格線交叉計數的研究為主,我們為了限 制渦蟲的運動空間與取得標準化的設備,在多方面的考量下,我們 將開發多種實驗方式,並選擇了市售的飲料瓶,利用商品的標準規 格來確保實驗器材的穩定性,收集更多關於渦蟲的運動數據。
10
第二章 材料與方法
壹、藥品與試劑一、藥品清單
藥品名 來源
硝酸銀(Silver Nitrate) Sigma-Aldrich 聚乙烯吡咯烷酮(PVP) Sigma-Aldrich 檸檬酸鈉(Sodium Citrate) Sigma-Aldrich 硼氫化鈉(Sodium Borohydride) Sigma-Aldrich 二、奈米銀溶液配製
AgNPs:將硝酸銀、檸檬酸鈉、硼氫化鈉、PVP 以二次水進行 配製,分別為 1.1*10-4M 硝酸銀溶液、3*10-2M 檸檬酸鈉溶液、2% PVP 溶液、0.1M 硼氫化鈉溶液。
三、渦蟲來源
購自邰港科技股份有限公司,學名:Dugesia japonica,鑑定 單位:國立嘉義大學淡水生物資源中心(附錄八)。
11
四、渦蟲準備工作
(一) 器材清單(圖 1)
1.飼養箱:長 47 公分、寬 34.5 公分、高 26 公分,容積為 42159 立方公分,保持水量 20 公升。
2.預備缸:長 26 公分、寬 17 公分、高 19 公分,容積為 8398 立方公分,保持水量 4 公升,實驗完畢後清洗,清洗 後裝水持續進行曝氣。
(二) 飼養方式
1.在塑膠箱中準備 20 公升二次水,打氣 48 小時以上,另 外準備容器裝水持續打氣,以便實驗所需或應變臨時狀 況。
2.將渦蟲放入,以 100 隻渦蟲/1 公升的比例飼養。
3.每天早上餵食,輕灑飼料 0.1 公克。
4.每周抽掉 1/3 水並補入 1/3 新鮮曝氣後的水。
(三) 實驗前準備:在實驗前 48 小時,將渦蟲從飼養箱中取出,
放置於預備缸中(圖 2),停止餵食。
12
圖 1 渦蟲實驗所需器材。A.飼養箱。B.預備缸。C.飼料:AZOO 9 合 1 人工輪蟲。D.AZOO 高溶氧生物過濾器 100L(水妖精)。
圖 2 在實驗室飼養渦蟲情形。A.飼養箱內情形-1。B.飼養箱內-2。
C.預備缸內情形-1。D.預備缸內情形-2。
13
貳、儀器
名稱 型號
超音波振盪器 DELTA DC300H 振盪器 VORTEX GENIE 2 搖盪器 FIRSTEK S-4100
電子天平 Electronic balance PL202-S 紫外光可見光光譜儀 Lambda EZ 201
參、實驗方法
一、奈米銀的合成與對渦蟲的毒性 (一) 奈米銀的合成
在奈米銀製備部分,我們以化學還原法製備 10nm 的黃色 奈米銀1。將檸檬酸鈉(Sodium Citrate)、聚乙烯吡咯烷酮(PVP)
及硝酸銀(AgNO3)混合,避光搖晃 10 分鐘,再將硼氫化鈉(NaBH4) 加入,超音波震盪一小時,生成球型黃色奈米銀溶液(大約 10 奈米)(圖 3)。
圖 3 本研究所使用的奈米銀。A.奈米銀避光保存。B.奈米銀溶液。
註 1.奈米銀尺寸為 10nm、黃色,由於論文發表中,無法展示相關資料。
14
(二)對渦蟲的進行藥物(奈米銀)處理
1.取 28 隻渦蟲,每 7 隻一組,對其中兩組渦蟲進行橫切手 術,並分別命名為沒切 0%、沒切 50%、切 0%、切 50% 1。 2.將渦蟲放置於培養皿中,把渦蟲身邊的水分吸乾。
3.取出稀釋後的 50%奈米銀 5 毫升加入培養皿中,並計時 1 小時。
4. 1 小時後,將渦蟲吸出置於 15 毫升離心管中。
5.使用塑膠吸管吸取 5 毫升二次水清洗渦蟲,並靜置 10 分 鐘,重複 3 次。
6.進行各項運動實驗並分析資料。
(三)奈米銀對渦蟲的毒性
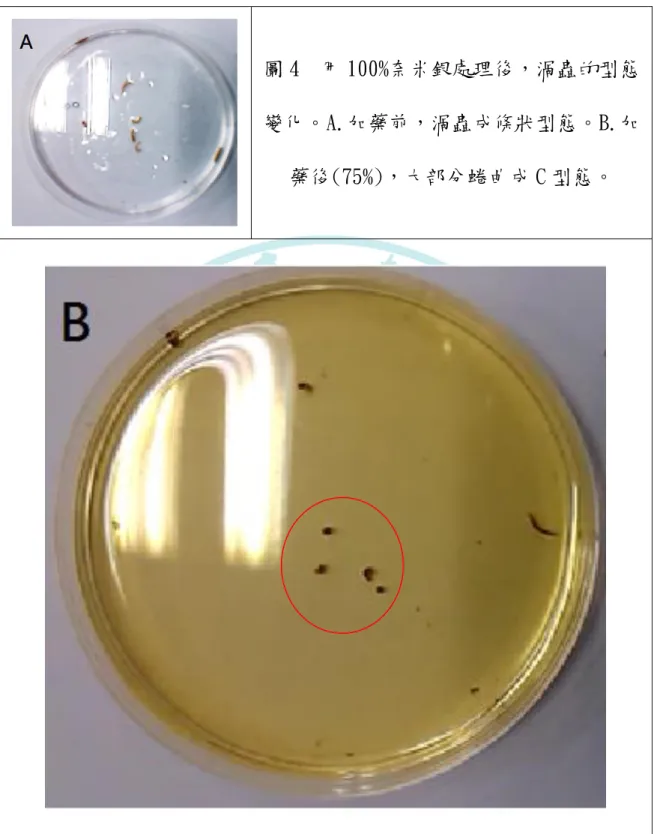
根據圖 4B 顯示,可以發現以奈米銀處理的渦蟲,會對環境 中的刺激(75%、100%奈米銀)產生 C 型態(C shape),其他濃度 請見附錄一、附錄二。在 100%奈米銀溶液內的渦蟲無法待超過 90 分鐘。
註 1.沒切 0%係指沒有進行橫切手術且以濃度 0%奈米銀處理。沒切 50%係指沒有進 行橫切手術且以濃度 50%奈米銀處理。切 0%係指有進行橫切手術且以濃度 0%奈米 銀處理。切 50%係指有進行橫切手術且以濃度 50%奈米銀處理。
15
圖 4 用 100%奈米銀處理後,渦蟲的型態 變化。A.加藥前,渦蟲成條狀型態。B.加
藥後(75%),大部分蜷曲成 C 型態。
16
二、渦蟲的預備實驗(色光、pLMV、圓形軌道) (一)色光實驗
由於生物體對光會有反應(例如:趨光性),渦蟲也是如此,
所以我們設計這個色光實驗,以自然光、紅光、藍光、綠光照 射渦蟲(圖 5),並觀察渦蟲在白色塑膠長盤中的活動情況。
實驗所使用的白色塑膠長盤有 10 個軌道,每個軌道寬度 為 0.6 公分,雖然軌道形狀可以讓渦蟲在裡面進行直線運動,
但空間太大,影片拍攝無法獲得較佳的效果,不利後續分析。
白色塑膠長盤 燈條及變壓器
色光實驗-對照組 色光實驗-紅光組
色光實驗-藍光組 色光實驗-綠光組
圖 5 色光實驗組。以光的三原色-紅光、綠光、藍光為主的實驗,
在塑膠長盤中,觀察渦蟲的運動狀態。
17
(二)pLMV 實驗
根據文獻,我們以格線版進行運動能力測試,雖然技術方 法簡單,但渦蟲靠近容器牆壁時,容易造成視覺誤差,而導致 數據錯誤(圖 6)。
格線交叉版 渦重運動速度(pLMV)數據收集中 圖 6 pLMV 實驗。將渦蟲放在裝水的容器中,背景用格線交叉版,
觀察並計數渦蟲所通過的方格。
(三)圓形軌道實驗
根據上述色光實驗與格線交叉實驗,我們希望透過容器的 限制,讓渦蟲在軌道中固定方向運動,而不產生過多的翻轉、
掉頭、東張西望等情況,本研究所使用的圓形軌道寬度為 0.2 公分(圖 7A),渦蟲的體寬為 0.05~0.08 公分(圖 7B),目的為 限制渦蟲的行動,且此圓形軌道為標準化商品,市面上極易取 得相同規格的產品,不必再耗費資源進行研發及改良。
圓形軌道 渦蟲寬度
圖 7 圓形軌道實驗。A.市售飲料瓶之圓形軌道。B.渦蟲寬度約為 0.05 公分~0.08 公分
18
三、運動行為資料蒐集:為了獲得良好的運動數據,我們制訂了幾 項條件:
(一)起始時間是蟲體入水到開始移動的過程,不記錄角度,屬 於探索時間。從其他研究中可以知道每種生物在進入一個陌生 的環境或是剛轉移環境時,都會先行探索,確認周遭狀況後才 會開始移動。
(二)身體呈現 U 形時,即可紀錄頭部位置。在本研究中,我們 以渦蟲頭部為記錄目標(起始位置與終點位置),當渦蟲想要改 變行進方向時,會將身體曲成 U 形,一旦頭部改變方向,便記 為下一段路程的起始位置。
(三)在圓形軌道中移動角度在 22.2 度以下的話,代表移動距 離沒有超過 1cm,記為探索時間。本研究所使用之渦蟲平均體 長為 1.0 公分,以圓形軌道的圓周角度換算約為 22.2 度,若 該蟲體移動角度不足 22.2 度(1.0 公分),則判斷該段時間是在 進行環境探索,故無法紀錄移動角度。
(四)順時針移動記為正;逆時針移動記為負。地球上有許許多 多的生物,其中有些生物可以藉由體內的磁性奈米粒子或是特 殊物質來感應地球磁場。由於本次渦蟲運動的場域是圓形軌道,
我們將渦蟲的運動行為以順時針、逆時針加以註記,可探討渦 蟲在圓形軌道的運動表現是否與地球磁場有些關聯。
19
(五)停頓的時間(2 秒內)不影響數據,可正常紀錄。渦蟲在移 動過程中,會有一些時後將頭部四周擺動,動作時間大約在 2 秒以內,對速率的計算而言,並不會影響數據的呈現,若 3 秒 以上,會明顯影響到數據的表現。
20
第三章、結果與討論
本研究針對圓形軌道進行分析,發現幾項特別且具有未來性的結 果,以下將針對移動前的探索時間(附錄三~附錄六)、單位時間內移動 方向(附錄七)等進行討論。
壹、渦蟲進行移動前的探索時間
一、橫切手術的行為會使探索時間增加
根據圖 8A 的資料可以發現,在 0%奈米銀溶液處理後,橫切手 術後的渦蟲平均探索時間為 5.9 秒,沒進行手術的渦蟲則是 4.5 秒;
在 50%奈米銀溶液處理後,橫切手術後的渦蟲平均探索時間為 6.6 秒,沒進行手術的渦蟲則是 6.3 秒。這樣的結果顯示,橫切手術會 導致探索的時間增加。
二、奈米銀溶液處理會使探索時間增加
根據圖 8A 的資料顯示,在沒進行手術的渦蟲中,以 0%奈米銀 溶液處理的渦蟲平均探索時間為 4.5 秒,以 50%奈米銀溶液處理的 是 6.3 秒;在有進行橫切手術的渦蟲中,以 0%奈米銀溶液處理的 渦蟲平均探索時間為 5.9 秒,以 50%奈米銀溶液處理的是 6.6 秒。
這樣的結果表示,用奈米銀溶液處理後的渦蟲,探索時間是增加 的。
21
三、奈米銀溶液處理的影響比進行橫切手術明顯
四種情況依探索時間進行排序:切 50%>沒切 50%>切 0%>沒切 0%。透過排序,我們可以發現,不論有沒有橫切手術,奈米銀溶液 處理後的渦蟲在平均探索時間排在前兩名,分別是 6.6 秒和 6.3 秒,
這樣的結果顯示,奈米銀溶液對渦蟲的運動行為影響比橫切手術更 加明顯。
四、奈米銀溶液處理與橫切手術有加成的效果
根據以上的討論,橫切手術會使探索時間增加,而奈米銀溶液 的處理也會使探索時間增加,若同時使用兩種條件處理的渦蟲,平 均探索時間(6.6 秒)是最多的,因此,我們可以得知,奈米銀溶液 處理與橫切手術會導致渦蟲的探索時間增加更多。
圖 8 探索時間。A.平均探索時間(秒)。B.平均表觀探索秒數(秒)。
22
貳、渦蟲在方向上的表現
表 1 渦蟲在順時針方向移動與逆時針方向移動之比例。
沒切 0%(對照組) 沒切 50% 切 0% 切 50%
順時針方向 移動比例
16.7% 16.7% 46.2% 35.7%
逆時針方向 移動比例
83.3% 83.3% 53.8% 64.3%
一、渦蟲移動時以逆時針方向為主
根據下表對照組(沒切-0%)所顯示的結果,大部分渦蟲在圓形 軌道中的移動表現是以逆時針方向為主,由表格(附錄七)可以發現,
逆時針方向的移動路徑總和普遍大於順時針方向。
沒切 0%(對照組) 沒切 50%
順時針方向移動比例 16.7% 16.7%
逆時針方向移動比例 83.3% 83.3%
23
二、橫切手術會造成渦蟲失去方向感
橫切手術導致渦蟲的順時針方向移動次數變多,從沒切 0%的 16.7%上升到切 0%的 46.2%;從沒切 50%的 16.7%上升到切 50%的 35.7%。該結果顯示,雖然渦蟲在切後繼續成長,但感官尚未完全 恢復,導致無法順利辨識方向。
沒切 0%(對照組) 切 0%
順時針方向移動比例 16.7% 46.2%
逆時針方向移動比例 83.3% 53.8%
沒切 50% 切 50%
順時針方向移動比例 16.7% 35.7%
逆時針方向移動比例 83.3% 64.3%
三、奈米銀粒子可以促進渦蟲恢復方向感
根據下表,可以發現橫切手術後的渦蟲經過奈米銀處理後,順 時針方向的移動比例下降(46.2%到 35.7%)、逆時針方向移動比例 上升(53.8%到 64.3%)。結果顯示,奈米銀可以讓渦蟲的方向感恢 復。
切 0% 切 50%
順時針方向移動比例 46.2% 35.7%
逆時針方向移動比例 53.8% 64.3%
24
參、渦蟲實驗的一些新發現和設計
從文獻中發現有些研究是透過藥物使渦蟲產生變異,而本研究 在實驗期間,也發現了不少新奇的東西。
一、雙頭渦蟲。本實驗室僅出現一隻,推測是自然變異,並非 以藥物進行改變。
圖 9 雙頭渦蟲在顯微鏡底下的影像。
二、紅色渦蟲。這個結果是令人興奮的,在文獻中,我們沒有 發現任何將渦蟲改變體色的相關研究,本次的體色改變方式並 非透過染劑或螢光染料,而是自然發生的體色改變,故可以將 顏色透過代謝排出體外,這樣的方法對渦蟲而言是無害的。
正常體色之渦蟲 紅色體色的渦蟲
圖 10 顯微鏡下,渦蟲體色改變的樣子。
25
三、3D 列印圓形軌道。本研究所使用的飲料瓶圓形軌道寬為 0.2 公分,對渦蟲而言還是太寬,且渦蟲可以採用「飛簷走壁」
的方式行動,所以我們嘗試尋找更小且可標準化的軌道,這樣 的難題讓我們想到時下最夯的科技-3D 列印,透過參數設定,
即可產生更棒的標準化軌道,這對未來渦蟲或微小動物的運動 資料收集有更好的發展空間。
圖 11 3D 列印圓形軌道實驗。A. 3D 列印機所印出來的圓形軌道寬 為 0.1 公分。B.渦蟲順利進入軌道中進行運動。
26
第四章 結論與未來展望
壹、渦蟲在接受物理性刺激(橫切手術)和化學性刺激(奈米銀)後,都 會有增加探索時間的狀況發生,顯示出這些刺激都會造成生物體的影 響,且藥物的刺激會大於物理性的刺激,所以在使用任何化學藥劑進 行研究時,都要詳細規劃與注意。
貳、學界對生物體在圓周運動時採逆時針策略的研究結果眾說紛紜,
而我們透過圓形軌道進行運動資料收集和分析,提出了渦蟲具有方向 感的概念,相信對未來在此方面的學說可以有一些貢獻。
參、紅色渦蟲的發現是令人驚豔的,透過食物改變體色,不但可以將 蟲體以顏色進行標示,且過程無須任何藥品或儀器協助,渦蟲還能自 行代謝排出體外,如此環保的生物標記方式,有助於渦蟲的相關研究。
肆、若能順利結合 3D 列印題材,對渦蟲的限制可以更加靈活,能獲得 的運動資料會更完善,同時參考本研究之開發模式,針對不同種類的 微小生物、生物體型,發展出一套檢測系統或檢測平台,讓運動行為 模式與毒性測試的研究更加廣泛。
27
第五章 參考文獻
[1] C. E. Stringer, "The means of locomotion in planarians," Proc. Natl. Acad. Sci. U. S. A., vol. 3, (12), pp. 691-692, Dec, 1917.
[2] L. Bertalanffy, O. Hoffmann-Ostenhof and O. Schreier, "A quantitative study of the toxic action of quinones on planaria gonocephala," Nature, vol. 158, (4026), pp. 948, Dec 28, 1946.
[3] W. C. Curtis, "Migration of formative cells in regeneration of planarian pharynx," Anat.
Rec., vol. 94, pp. 358, Mar, 1946.
[4] E. D. Goldsmith, "Thiourea and head regeneration in planarians," Anat. Rec., vol. 96, (4), pp. 581, Dec, 1946.
[5] G. Marsh and H. W. Beams, "Electrical control of growth polarity in regenerating Dugesia tigrina," Fed. Proc., vol. 6, (1 Pt 2), pp. 163, 1947.
[6] T. Inoue, H. Hoshino, T. Yamashita, S. Shimoyama and K. Agata, "Planarian shows decision-making behavior in response to multiple stimuli by integrative brain function,"
Zoological Lett., vol. 1, pp. 7-014-0010-z. eCollection 2015, Feb 1, 2015.
[7] R. Langer, M. Fefferman, P. Gryska and K. Bergman, "A simple method for studying chemotaxis using sustained release of attractants from inert polymers," Can. J. Microbiol., vol. 26, (2), pp. 274-278, Feb, 1980.
[8] P. R. Mason, "Chemo-klino-kinesis in planarian food location," Anim. Behav., vol. 23, (2), pp. 460-469, May, 1975.
[9] S. D. Cross, A. A. Johnson, B. J. Gilles, L. A. Bachman, T. Inoue, K. Agata, L. Y.
Marmorstein and A. D. Marmorstein, "Control of Maintenance and Regeneration of Planarian Eyes by ovo," Invest. Ophthalmol. Vis. Sci., vol. 56, (12), pp. 7604-7610, Nov, 2015.
[10] J. P. Dexter, M. B. Tamme, C. H. Lind and E. M. Collins, "On-chip immobilization of planarians for in vivo imaging," Sci. Rep., vol. 4, pp. 6388, Sep 17, 2014.
28
[11] T. R. Paskin, J. Jellies, J. Bacher and W. S. Beane, "Planarian Phototactic Assay Reveals Differential Behavioral Responses Based on Wavelength," PLoS One, vol. 9, (12), pp.
e114708, Dec 10, 2014.
[12] G. Pescetto and D. Dettore, "Negative phototaxis and conditioning in the planarian Dugesia dorotocephala," Riv. Neurobiol., vol. 27, (2), pp. 287-295, Apr-Jun, 1982.
[13] L. Balestrini, A. Di Donfrancesco, L. Rossi, S. Marracci, M. E. Isolani, A. M. Bianucci and R. Batistoni, "The natural compound sanguinarine perturbs the regenerative capabilities of planarians," Int. J. Dev. Biol., vol. 61, (1-2), pp. 43-52, 2017.
[14] Y. Ben Khadra, M. Sugni, C. Ferrario, F. Bonasoro, A. Varela Coelho, P. Martinez and M. D. Candia Carnevali, "An integrated view of asteroid regeneration: tissues, cells and molecules," Cell Tissue Res., Mar 22, 2017.
[15] N. A. Bonar and C. P. Petersen, "Integrin suppresses neurogenesis and regulates brain tissue assembly in planarian regeneration," Development, vol. 144, (5), pp. 784-794, Mar 1, 2017.
[16] D. D. R. Brown and B. J. Pearson, "A Brain Unfixed: Unlimited Neurogenesis and Regeneration of the Adult Planarian Nervous System," Front. Neurosci., vol. 11, pp. 289, May 23, 2017.
[17] J. D. Chan, D. Zhang, X. Liu, M. Zarowiecki, M. Berriman and J. S. Marchant,
"Utilizing the planarian voltage-gated ion channel transcriptome to resolve a role for a Ca2+
channel in neuromuscular function and regeneration," Biochim. Biophys. Acta, vol. 1864, (6), pp. 1036-1045, Jun, 2017.
[18] X. Chen and C. Xu, "Proteomic Analysis Reveals the Contribution of TGFbeta/Smad4 Signaling Pathway to Cell Differentiation During Planarian Tail Regeneration," Appl.
Biochem. Biotechnol., vol. 182, (2), pp. 529-545, Jun, 2017.
[19] A. Y. T. Lin and B. J. Pearson, "Yorkie is required to restrict the injury responses in planarians," PLoS Genet., vol. 13, (7), pp. e1006874, Jul 7, 2017.
29
[20] J. Morokuma, F. Durant, K. B. Williams, J. M. Finkelstein, D. J. Blackiston, T.
Clements, D. W. Reed, M. Roberts, M. Jain, K. Kimel, S. A. Trauger, B. E. Wolfe and M.
Levin, "Planarian regeneration in space: Persistent anatomical, behavioral, and
bacteriological changes induced by space travel," Regeneration (Oxf), vol. 4, (2), pp. 85-102, Jun 13, 2017.
[21] C. P. Petersen, "Regeneration: Organizing the Blastema in Planarians," Curr. Biol., vol.
27, (5), pp. R181-R183, Mar 6, 2017.
[22] C. Bezerra da Silva, A. Pott, S. Elifio-Esposito, L. Dalarmi, K. Fialho do Nascimento, L.
Moura Burci, M. de Oliveira, J. de Fatima Gaspari Dias, S. M. Warumby Zanin, O. Gomes Miguel and M. Dallarmi Miguel, "Effect of Donepezil, Tacrine, Galantamine and
Rivastigmine on Acetylcholinesterase Inhibition in Dugesia tigrina," Molecules, vol. 21, (1), pp. 53, Jan 11, 2016.
[23] M. S. Farrell, K. Gilmore, R. B. Raffa and E. A. Walker, "Behavioral characterization of serotonergic activation in the flatworm Planaria," Behav. Pharmacol., vol. 19, (3), pp.
177-182, May, 2008.
[24] R. B. Raffa, F. Cavallo and A. Capasso, "Flumazenil-sensitive dose-related physical dependence in planarians produced by two benzodiazepine and one non-benzodiazepine benzodiazepine-receptor agonists," Eur. J. Pharmacol., vol. 564, (1-3), pp. 88-93, Jun 14, 2007.
[25] R. B. Raffa, L. J. Holland and R. J. Schulingkamp, "Quantitative assessment of dopamine D2 antagonist activity using invertebrate (Planaria) locomotion as a functional endpoint," J. Pharmacol. Toxicol. Methods, vol. 45, (3), pp. 223-226, May-Jun, 2001.
[26] R. B. Raffa and A. F. Martley, "Amphetamine-induced increase in planarian locomotor activity and block by UV light," Brain Res., vol. 1031, (1), pp. 138-140, Jan 7, 2005.
[27] L. Ramakrishnan, C. Amatya, C. J. DeSaer, Z. Dalhoff and M. R. Eggerichs,
"Galantamine reverses scopolamine-induced behavioral alterations in Dugesia tigrina," Invert.
Neurosci., vol. 14, (2), pp. 91-101, Sep, 2014.
[28] S. M. Rawls, T. Gomez and R. B. Raffa, "An NMDA antagonist (LY 235959) attenuates abstinence-induced withdrawal of planarians following acute exposure to a cannabinoid agonist (WIN 55212-2)," Pharmacol. Biochem. Behav., vol. 86, (3), pp. 499-504, Mar, 2007.
30
[29] S. M. Rawls, T. Rodriguez, D. A. Baron and R. B. Raffa, "A nitric oxide synthase inhibitor (L-NAME) attenuates abstinence-induced withdrawal from both cocaine and a cannabinoid agonist (WIN 55212-2) in Planaria," Brain Res., vol. 1099, (1), pp. 82-87, Jul 12, 2006.
[30] A. C. Rodrigues, J. F. Henriques, I. Domingues, O. Golovko, V. Zlabek, C. Barata, A.
M. Soares and J. L. Pestana, "Behavioural responses of freshwater planarians after short-term exposure to the insecticide chlorantraniliprole," Aquat. Toxicol., vol. 170, pp. 371-376, Jan, 2016.
[31] J. Zhang, Z. Yuan, M. Zheng, Y. Sun, Y. Wang and S. Yang, "Effects of
N,N-dimethylformamide on behaviour and regeneration of planarian Dugesia japonica,"
Toxicol. Ind. Health, vol. 29, (8), pp. 753-760, Sep, 2013.
[32] M. Fiore, R. Moroni and L. Aloe, "Removal of the submaxillary salivary glands and infection with the trematode Schistosoma mansoni alters exploratory behavior and pain thresholds in female mice," Physiol. Behav., vol. 62, (2), pp. 399-406, Aug, 1997.
[33] W. Haas and S. Haeberlein, "Penetration of cercariae into the living human skin:
Schistosoma mansoni vs. Trichobilharzia szidati," Parasitol. Res., vol. 105, (4), pp.
1061-1066, Oct, 2009.
[34] D. R. Hamilton, "Immunosuppressive effects of predator induced stress in mice with acquired immunity to Hymenolepis nana," J. Psychosom. Res., vol. 18, (3), pp. 143-153, Jun, 1974.
[35] J. Koprivnikar, C. H. Gibson and J. C. Redfern, "Infectious personalities: behavioural syndromes and disease risk in larval amphibians," Proc. Biol. Sci., vol. 279, (1733), pp.
1544-1550, Apr 22, 2012.
[36] O. R. Pagan, A. L. Rowlands and K. R. Urban, "Toxicity and behavioral effects of dimethylsulfoxide in planaria," Neurosci. Lett., vol. 407, (3), pp. 274-278, Oct 30, 2006.
[37] C. P. READ, "Soboliphyme jamesoni n. sp., a curious nematode parasite of California shrews," J. Parasitol., vol. 38, (3), pp. 203-206, Jun, 1952.
[38] T. Shomrat and M. Levin, "An automated training paradigm reveals long-term memory in planarians and its persistence through head regeneration," J. Exp. Biol., vol. 216, (Pt 20), pp. 3799-3810, Oct 15, 2013.
31
[39] A. M. Abdel-Mohsen, J. Jancar, R. M. Abdel-Rahman, L. Vojtek, P. Hyrsl, M. Duskova and H. Nejezchlebova, "A novel in situ silver/hyaluronan bio-nanocomposite fabrics for wound and chronic ulcer dressing: In vitro and in vivo evaluations," Int. J. Pharm., vol. 520, (1-2), pp. 241-253, Mar 30, 2017.
[40] A. Ali, I. U. Haq, J. Akhtar, M. Sher, N. Ahmed and M. Zia, "Synthesis of Ag-NPs impregnated cellulose composite material: its possible role in wound healing and photocatalysis," IET Nanobiotechnol, vol. 11, (4), pp. 477-484, Jun, 2017.
[41] Z. Di, Z. Shi, M. W. Ullah, S. Li and G. Yang, "A transparent wound dressing based on bacterial cellulose whisker and poly(2-hydroxyethyl methacrylate)," Int. J. Biol. Macromol., Jul 14, 2017.
[42] E. Gee Kee, K. Stockton, R. M. Kimble, L. Cuttle and S. M. McPhail,
"Cost-effectiveness of silver dressings for paediatric partial thickness burns: An economic evaluation from a randomized controlled trial," Burns, vol. 43, (4), pp. 724-732, Jun, 2017.
[43] F. M. Helaly, S. M. El-Sawy, A. I. Hashem, A. A. Khattab and R. M. Mourad,
"Synthesis and characterization of nanosilver-silicone hydrogel composites for inhibition of bacteria growth," Cont Lens Anterior Eye, vol. 40, (1), pp. 59-66, Feb, 2017.
[44] S. Huang, Z. Yu, Y. Zhang, C. Qi and S. Zhang, "In situ green synthesis of
antimicrobial carboxymethyl chitosan-nanosilver hybrids with controlled silver release," Int.
J. Nanomedicine, vol. 12, pp. 3181-3191, Apr 18, 2017.
[45] T. Jayaramudu, K. Varaprasad, G. M. Raghavendra, E. R. Sadiku, K. Mohana Raju and J. Amalraj, "Green synthesis of tea Ag nanocomposite hydrogels via mint leaf extraction for effective antibacterial activity," J. Biomater. Sci. Polym. Ed., pp. 1-15, Jun 22, 2017.
[46] S. Kaosaar, A. Kahru, P. Mantecca and K. Kasemets, "Profiling of the toxicity
mechanisms of coated and uncoated silver nanoparticles to yeast Saccharomyces cerevisiae BY4741 using a set of its 9 single-gene deletion mutants defective in oxidative stress response, cell wall or membrane integrity and endocytosis," Toxicol. in. Vitro., vol. 35, pp.
149-162, Sep, 2016.
[47] K. W. Kwok, W. Dong, S. M. Marinakos, J. Liu, A. Chilkoti, M. R. Wiesner, M.
Chernick and D. E. Hinton, "Silver nanoparticle toxicity is related to coating materials and disruption of sodium concentration regulation," Nanotoxicology, vol. 10, (9), pp. 1306-1317, Nov, 2016.
32
[48] J. D. Martin, T. L. Colson, V. S. Langlois and C. D. Metcalfe, "Biomarkers of exposure to nanosilver and silver accumulation in yellow perch (Perca flavescens)," Environ. Toxicol.
Chem., vol. 36, (5), pp. 1211-1220, May, 2017.
[49] D. Sundeep, T. Vijaya Kumar, P. S. S. Rao, R. V. S. S. N. Ravikumar and A. Gopala Krishna, "Green synthesis and characterization of Ag nanoparticles from Mangifera indica leaves for dental restoration and antibacterial applications," Prog. Biomater., vol. 6, (1-2), pp.
57-66, May, 2017.
[50] T. BETCHAKU, "A copper sulfate-silver nitrate method for nerve fibers of planarians,"
Stain Technol., vol. 35, pp. 215-218, Jul, 1960.
[51] I. D. Bowen, T. A. Ryder and C. Winters, "The distribution of oxidizable
mucosubstances and polysaccharides in the planarian Polycelis tenuis Iijima," Cell Tissue Res., vol. 161, (2), pp. 263-275, Aug 18, 1975.
[52] L. L. DiCiaula, G. L. Foley and D. J. Schaeffer, "Fixation and staining of planaria for histological study," Biotech. Histochem., vol. 70, (3), pp. 119-123, May, 1995.
[53] I. Hori, "Localization of newly synthesized precursors of basal lamina in the
regenerating planarian as revealed by autoradiography," Tissue Cell, vol. 12, (3), pp. 513-521, 1980.
[54] L. Kustov, K. Tiras, S. Al-Abed, N. Golovina and M. Ananyan, "Estimation of the toxicity of silver nanoparticles by using planarian flatworms," Altern. Lab. Anim., vol. 42, (1), pp. 51-58, Mar, 2014.
33
附錄一
C 型態觀察加藥前 加藥後
切 25%
切 50%
切 75%
切 100%
34
附錄二
C 型態觀察加藥前 加藥後
沒切 25%
X
沒切 50%
沒切 75%
沒切 100%
35
附錄三
探索時間數據表格(沒切 0%)
沒切0 % -1 沒切0 % -2 沒切0 % -3 沒切0 % -4 沒切0 % -5 沒切0 % -6 1 2.0 15 .0 8.0 10 .0 6.0 7.0 2 7.0 1.0 1.0 2.0 1.0 3.0 3 7.0 8.0 1.0 2.0 1.0 2.0 4 4.0 5.0 2.0 2.0 3.0 2.0 5 2.0 4.0 3.0 4.0 1.0 9.0 6 1.0 2.0 2.0 2.0 6.0 3.0 7 5.0 4.0 15 .0 4.0 4.0 1.0 8 1.0 9.0 5.0 3.0 2.0 2.0 9 7.0 6.0 2.0 1.0 1.0 28 .0 10 13 .0 1.0 3.0 2.0 11 4.0 5.0 8.0 2.0 12 4.0 3.0 2.0 13 6.0 14 2.0 總秒數 65 .0 60 .0 39 .0 44 .0 31 .0 57 .0 平均 4.6 5.5 4.3 3.7 2.6 6.3
36
附錄四
探索時間數據表格(沒切 50%)
沒切 50 % -1 沒切 50 % -2 沒切 50 % -3 沒切 50 % -4 沒切 50 % -5 沒切 50 % -6 1 2.0 2.0 3.0 1.0 20 .0 9.0 2 5.0 16 .0 6.0 7.0 8.0 1.0 3 26 .0 3.0 2.0 3.0 2.0 4 3.0 3.0 12 .0 5.0 5.0 5 2.0 3.0 18 .0 2.0 18 .0 6 9.0 3.0 7 2.0 2.0 8 1.0 9.0 9 3.0 4.0 10 7.0 總秒數 38 .0 42 .0 41 .0 8.0 38 .0 60 .0 平均 7.6 4.7 8.2 4.0 7.6 6.0
37
附錄五
探索時間數據表格(切 0%)
切0%-1切0%-2切0%-3切0%-4切0%-5切0%-6切0%-7切0%-8切0%-9切0%-10切0%-11切0%-12切0%-13113.04.03.03.019.031.012.033.08.01.01.047.012.028.015.09.03.02.012.08.02.02.02.03.02.02.033.03.04.01.03.011.04.01.01.02.025.04.016.041.018.01.01.05.03.02.01.02.01.02.09.055.07.08.03.06.01.07.03.02.03.02.04.068.04.05.01.03.07.011.02.01.03.09.01.073.03.08.02.02.02.02.02.06.081.01.03.04.02.01.02.095.07.04.01.09.02.03.0101.03.04.02.011.01116.03.07.0125.05.02.0132.0142.0總秒數47.051.016.033.068.079.058.056.019.039.038.077.044.0平均5.28.55.33.35.77.94.89.32.72.85.48.67.3
38
附錄六
探索時間數據表格(切 50%)
切50%-1切50%-2切50%-3切50%-4切50%-5切50%-6切50%-7切50%-8切50%-9切50%-10切50%-11切50%-12切50%-13切50%-1418.023.019.03.03.02.012.03.07.08.02.044.016.02.022.05.05.010.014.03.019.01.05.05.03.02.03.04.033.07.01.09.01.013.03.02.03.04.03.03.02.02.048.01.01.08.01.06.09.02.02.04.02.02.015.04.053.013.01.02.01.09.03.09.09.06.03.026.02.02.061.02.02.034.019.02.011.01.03.00.06.02.02.09.0712.05.03.023.010.06.026.03.03.05.07.084.035.04.02.04.05.011.03.03.013.093.09.02.02.04.02.01018.02.02.02.04.01112.03.0121.0131.0142.0153.0總秒數81.0100.036.066.039.035.086.032.040.068.031.094.045.043.0平均5.411.14.511.06.55.88.64.05.06.83.18.56.45.4
39
附錄七
順/逆時針方向移動數據
切0%196.851.5436.739.5262111-25.8-309.1-54.6-79.5-105.8-310.8-252.2切50%1139.7409.425.117.9-63.2-668.9-88.5-287.1-97.1-80.3-106.4-239.9-101.2沒切0%202.2-407.6-387.8-25.9-182-188.9沒切50%278.6-279.1-531.8-1428-335.8-231.5 順時針方向移動角度(度)逆時針方向移動角度(度)
40