Abstract In this study, we examined interaction between cadmium (Cd) and temperature in rice seedlings. Effects of CdCl2 (0.5 mM) simulta-neously applied at high (35/30°C day/night), medium (25/20°C) and low (15/13°C) tempera-tures to rice seedlings were detected by measur-ing changes in biomass production and NH4+ content. Results indicated that Cd-induced bio-mass reduction and NH4+accumulation increased in parallel with temperature increases. On treat-ment with CdCl2, the abscisic acid (ABA) content markedly increased in the leaves of seedlings grown at high temperature but not at medium and low temperatures. Exogenous application of ABA at medium temperature increased ABA and NH4+contents in the leaves of rice seedlings. Fluridone (Flu) treatment, an inhibitor of carot-enoid biosynthesis, reduced ABA content, as well as Cd-induced NH4+accumulation in the leaves of rice seedlings grown at high temperature. These Flu effects can be reversed by application of ABA. Furthermore, Flu treatment did not reduce Cd content in leaves of seedlings grown at high temperature. All these results suggest that Cd-induced NH4+accumulation at high temperature is mediated through ABA.
Keywords Abscisic acid Æ Ammonium ion Æ Cadmium Æ Oryza sativa Æ Temperature Abbreviations
ABA Abscisic acid DW Dry weight
ELISA Enzyme-linked immunosorbent assay Flu Fluridone
GS Glutamine synthetase
NCED 9-cis-epoxycarotenoid dioxygenase PAL Phenylalanine ammonia-lyase
Introduction
Cadmium (Cd) is a divalent heavy metal cation and is one of the most toxic heavy metals. It is supplied to soil, air, and water mainly by effluents from industries, mining, burning, and leakage waste, and by application of phosphate and sew-age sludge. Cd is readily taken up by plants, leading to inhibition of physiological processes such as respiration, photosynthesis, cell elonga-tion, plant-water relationships, nitrogen metabo-lism, and mineral nutrition (Sanita´ di Toppi and Gabbrielli1999).
Global warming, accompanied by an increased frequency of periods with exceptionally high Y. T. Hsu Æ M. C. Kuo Æ C. H. Kao (&)
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China e-mail: [email protected]
DOI 10.1007/s11104-006-9076-5 O R I G I N A L P A P E R
Cadmium-induced ammonium ion accumulation of rice
seedlings at high temperature is mediated through abscisic
acid
Yi Ting Hsu Æ Mei Chun Kuo Æ Ching Huei Kao
Received: 21 February 2006 / Accepted: 28 June 2006 / Published online: 24 August 2006 ÓSpringer Science+Business Media B.V. 2006
temperatures, is one of the most important char-acteristics of the accelerated climatic changes. Climatic models predict that global temperature will continue to increase in the future. Peng et al. (2004) analyzed weather data at the International Rice Research Institute Farm from 1979 to 2003 to examine temperature trends and reported that annual mean maximum and minimum tempera-tures have increased by 0.35°C and 1.13°C, respectively. Hence, plants will be more often exposed to high temperature conditions. It has been shown that Cd toxicity (the reduction of plant height, the decrease in chlorophyll content, and the accumulation of phenolic compounds and proline) of wheat seedlings increases in parallel to temperature increases (O¨ ncel et al. 2000). Tem-perature is a key factor in altering ion accumu-lation (Chawla et al. 1991; Hooda and Alloway 1993; Macek et al. 1994; Mautsoe and Beckett 1996). The high toxicity of Cd induced by high temperature may be the results of much more Cd uptake from the medium.
NH4+ is a central intermediate of nitrogen metabolism (Miflin and Lea 1976). It has been shown that on treatment with CdCl2,NH4+content increased in rice leaves (Hsu and Kao 2003). Glutamine synthetase (GS) is the key enzyme in NH4+assimilation and catalyzes the ATP-depen-dent condensation of NH4+ with glutamate to produce glutamine (Miflin and Lea 1976). Phen-ylalanine ammonia–lyase (PAL) catalyzes the elimination of NH4+from phenylalanine and pro-duces trans-cinnamate (Hahlbrock and Grisebach 1979). NH4+, released from PAL reaction, is known to be trapped in the glutamine molecule by the action of GS (Razel et al. 1996; van Heerden et al. 1996). Sakurai et al. (2001) pro-vided evidence to show that GS is partly coupled to the reaction of PAL in developing rice leaves. Cd-induced NH4+accumulation in rice leaves has been proved to be associated with the decrease in GS activity and the increase in PAL specific activity (Hsu and Kao 2004).
The plant hormone ABA is a sesquiterpenoid derived from xanthophyll (Nambara and Marion-Poll 2005) and appears to influence several physiological and developmental events (Seo and Koshiba2002; Zeevaart and Creelman1988). The level of ABA in plant tissues is known to increase
in reponse to exposure to heavy metals such as Cd, Ni, Zn, and Al (Fediuc et al.2005; Foy1998; Hollenbach et al.1997; Hsu and Kao 2003; Pos-chenrieder et al. 1989; Rauser and Dumbroff 1981). Fediuc et al. (2005) demonstrated that Cd-induced ABA accumulation was observed in roots, but not shoots, of Typha and Phragmites plants. Hsu and Kao (2003) found that, on treat-ment with CdCl2, the ABA content rapidly in-creased in the leaves and roots in Cd-tolerant rice cultivar (cv. Tainung 67) grown at normal grow-ing temperature (30/25°C, day/night) but not in Cd-sensitive cultivar (cv. Taichung Native 1).
In this study, effects of Cd applied at low (15/ 13°C day/night), medium (25/20°C day/night), and high (35/30°C day/night) temperatures on the content of NH4+ in leaves of rice seedlings were compared. Recently, we showed that ABA treat-ment resulted in an accumulation in NH4+ in de-tached rice leaves (Hung and Kao2005). We thus examined the possibility that ABA plays a role in mediating the Cd-induced NH4+ accumulation in leaves of rice seedlings grown at high temperature.
Materials and methods Plant material and treatments
Rice (Oryza sativa L., cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypo-chlorite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri-dishes with wetted filter papers at 37°C in the dark. After 48 h incubation, uniformly ger-minated seeds were selected and cultivated in a 250 ml beaker containing half-strength Kimura B solution containing the following macro- and micro-elements: 182.3 lM (NH4)2SO4, 91.6 lM KNO3, 273.9 lM MgSO4Æ7H2O, 91.1 lM KH2PO4, 182.5 lM Ca(NO3)2, 30.6 lM Fe-citrate, 0.25 lM H3BO3, 0.2 lM MnSO4ÆH2O, 0.2 lM ZnSO4 Æ7-H2O, 0.05 lM CuSO4Æ5H2O, and 0.07 lM H2MoO4. The nutrient solutions (pH 4.7) were replaced every 3 days. The hydroponically culti-vated seedlings were grown in a Phytotron (Agri-cultural Experimental Station, National Taiwan University, Taipei, Taiwan) with natural sunlight at 30/25°C day/night and 90% relative humidity.
Twelve-day-old seedlings with three leaves were grown in basic nutrient solution with or without CdCl2 at high (35/30°C day/night), medium (25/ 20°C day/night), or low temperature (15/13°C day/ night). Based on our experience from the experi-ments of Cd effect on rice seedlings, consistent Cd toxicity was observed in 2 days when 0.5 mM CdCl2 was used for experiments. However, no consistent Cd toxicity was observed when lower CdCl2 con-centrations, in the range 5–50 mM, were applied over a longer period (6 days). For this reason in this study, 0.5 mM CdCl2was used and added directly to nutrient solution. For the experiments in which the effect of exogenous ABA was examined, various concentrations of ABA (5–40 mM, mixed isomers) were added directly to the culture solution for 2 days at medium temperature. In experiments to understand the role of endogenous ABA, 0.2 mM fluridone (Flu), which is known to block the con-version of phytoene to phytofluene in carotenoid biosynthesis pathway (Kowalczyk-Schro¨der and Sandmann1992), was added directly to the nutrient solution for 2 days.
Growth analysis
At the end of treatment, the seedlings were di-vided into separate parts (shoot, and roots). The dry weight (DW) of the shoot and roots (adven-titious roots plus primary roots) were then mea-sured. For DW estimation, the shoot and roots were dried at 65°C for 48 h.
Cd determination
For determination of Cd, leaves were dried at 65°C for 48 h. Dried material was ashed at 550°C for 20 h. The ash residue was incubated with 31% HNO3 and 17.5% H2O2 at 72°C for 2 h, and dissolved in distilled water. Cd was then quanti-fied using an atomic absorption spectrophotomer (Model AA-6800, Shimadzu, Kyoto, Japan). Amount of Cd is expressed on the basis of DW. Determinations of protein, NH4+, and abscisic acid
For protein determination, leaves were homoge-nized in a 50 mM sodium phosphate buffer (pH
6.8). The extracts were centrifuged at 17,600 g for 20 min, and the supernatants were used for determination by the method of Bradford (1976). NH4+ was measured in the crude extract by the Berthelot reaction, modified according to Weatherburn (1967). The detailed procedure was described previously (Lin and Kao 1996). NH4+ contents are expressed on the basis of DW.
For extraction of ABA, leaves were homog-enized with a pestle and mortar in extraction solution (80% methanol containing 2% glacial acetic acid). To remove plant pigments and other non-polar compounds which could inter-fere in the immunoassay, extracts were first passed through polyvinylpyrrolidone column and C18 (Sep-Pak Vac) cartridges (Waters, Milford, MA). The eluates were concentrated to dryness by vacuum-evaporation and resuspended in Tris-buffered saline before enzyme-linked immuno-sorbent assay (ELISA). ABA was quantified by ELISA (Walker-Simmons, 1987). ABA immu-noassay detection kit (PGR-1) was purchased from Sigma Chemical Co. (St. Louis, MO) is specific for (+)-ABA. By evaluating 3H-ABA recovery, 3H-ABA loss was (3% by the method described here. ABA content is expressed on the basis of DW.
Enzyme assays
For extraction of GS, leaf samples were homog-enized with 10 mM Tris–HCl buffer (pH 7.6, containing 1 mM MgCl2, 1 mM EDTA, and 1 mM 2-mercaptoethanol) using a chilled pestle and mortar. The homogenate was centrifuged at 15,000 g for 30 min and the resulting supernatant was used for determination of GS activity. The whole extraction procedure was carried out at 4°C. GS was assayed by the method of Oaks et al. (1980). The reaction mixture contained in a final volume of 1 ml was 80 lmol Tris–HCl buffer, 40 lmol L-glutamic acid, 8 lmol ATP, 24 lmol MgSO4, and 16 lmol NH2OH; the final pH was 8.0. The reaction was started by addition of the enzyme extract and, after incubation for 30 min at 30°C, was stopped by adding 2 ml 2.5% (w/v) FeCl3and 5% (w/v) trichloroacetic acid in 1.5 M HCl. After centrifugation the absorbance of the supernatant was read at 540 nm. One unit of
GS activity is defined as 1 lmol L-glutamate
c-monohydroxamate formed per min.
Phenylalanine ammonia-lyase (PAL) was extracted and determined according to Hyodo and Fujinami (1989). The calculation was based on the extinction coefficient (9500 M–1cm–1) for trans-cinnamic acid. One unit of activity for PAL was defined as the amounts of enzyme which caused the formation of 1 lmol trans-cinnamic acid per hour. Transpiration rate
The transpiration rate was measured according to Greger and Johansson (1992). The weight of rice seedlings grown in hydroponic solution was determined at the beginning and the end of interval, respectively. The transpiration rate was calculated for the water loss during each interval and converted to a per day per seedling basis. Statistical analysis
Statistical differences between measurements (n = 4) on different treatments or on different times were analyzed following LSD test.
Results
Biomass production
In the absence of CdCl2, no significant difference in shoot DW of rice seedlings treated with dif-ferent temperatures was observed (Fig.1A). CdCl2 treatment resulted in a 15% reduction of shoot DW at high temperature but not at low and medium temperatures (Fig. 1A). Low tempera-ture treatment had lower root DW of rice seed-lings in the absence of CdCl2 than medium and high temperatures (Fig.1B). CdCl2 treatment reduced root DW to the level of 70% of the un-treated seedlings at high and medium tempera-tures (Fig.1B). However, Cd had no effect on root DW at low temperature (Fig. 1B).
Effect of temperatures on NH4+content
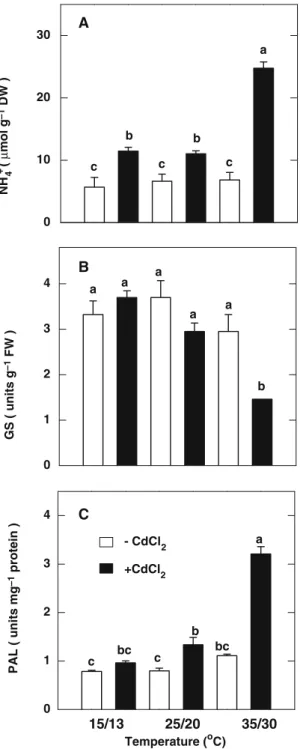
NH4+ is a central intermediate of nitrogen metabolism in plants (Miflin and Lea1976). Here,
we show that on treatment with CdCl2, NH4+ content increases about 3-fold in the second leaves of rice seedlings grown at high tempera-ture, but 80% at medium and low temperatures (Fig.2A). At high temperature, CdCl2treatment resulted in a 50% decrease in GS activity in the second leaves (Fig.2B). However, there was no decrease in GS activity in the second leaves of rice seedlings treated with CdCl2at medium and low temperatures (Fig.2B). CdCl2 treatment re-sulted in about 3-fold increase in PAL specific activity in leaves of rice seedlings grown at high
15/13 25/20 35/30 Temperature ( oC) Shoot DW (mg seedling –1 ) 0 2 4 6 8 10 12 14 0 2 4 6 8 a a a a
b
Root DW (mg seedling –1 ) a a a A B -CdCl 2 +CdCl 2 b b b b bFig. 1 Effect of temperatures on the dry weight of shoot (A) and roots (B) of rice seedlings treated with or without CdCl2(0.5 mM). All measurements were made two days after treatment. Bars indicate standard error (n = 4). Values with the same letter are not significantly different at P < 0.05
temperature (Fig.2C). However, slight (30%) and no increase in PAL specific activity in the
second leaves of rice seedlings treated with CdCl2 at medium and low temperatures, respectively (Fig.2C).
Effect of temperatures on Cd content
CdCl2treatment resulted in 30-, 11-, and 13-fold increase in Cd content in the third leaves (Fig.3A), 32-, 3-, and 2.4-fold increase in the second leaves (Fig.3B), and 69-, 48-, and 19-fold increase in roots (Fig.3C) at high, medium, and low temperatures, respectively. The results in Figs.3A–C are consistent with the idea that the amount of Cd reaching the shoot is usually lower than the amount in the roots (Javis et al.1976). Figure4 demonstrated that the increase in Cd content in the second leaves was evident 8, 24, and 36 h after CdCl2treatment at high, medium, and low temperatures, respectively.
Effect of temperatures on transpiration rate and ABA content
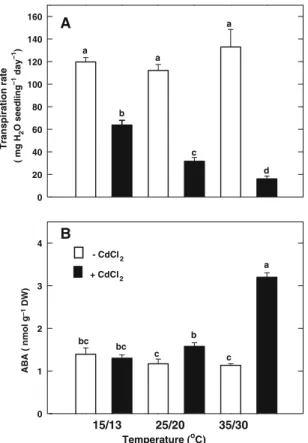
Cd has been shown to decrease transpiration rate in several plants (Hagemeyer et al. 1986; Kirkham 1978; Lamoreaux and Chaney 1978; Schlegel et al. 1987). We also observed that Cd decreased transpiration rate in rice seedlings (Fig.5A). CdCl2 treatment reduced transpira-tion rate of rice seedlings grown at low, med-ium, and high temperatures, respectively, to 53%, 29%, and 13% of the control values (Fig.5A). At high temperature, CdCl2 treat-ment resulted in about 3-fold increase in endogenous ABA in the second leaves (Fig.5B). However, 25% and no increase in ABA content in the second leaves of rice seedlings treated with CdCl2at medium and low temperatures, respectively (Fig.5B).
Exogenous application of ABA
To study the effect of exogenous ABA, various concentrations of ABA (5–40 lM) were added directly to the culture solution at medium tem-perature. The results indicated that exogenous ABA treatment increased endogenous ABA and NH4+ contents in the second leaves of rice seed-lings grown at medium temperature (Figs.6A, B).
NH ( µ mol g –1 DW ) 4 + 0 10 20 30 c c c b b a Temperature (oC) GS ( units g –1 FW ) 0 1 2 3 4 0 1 2 3 4 a a a a a b PAL ( units mg –1 protein ) a b bc bc c c A B C - CdCl +CdCl2 2 15/13 25/20 35/30
Fig. 2 Effect of temperatures on the contents of NH4+(A), and the activities of GS (B), and the specific activities of PAL (C) in the second leaves of rice seedlings treated with or without CdCl2(0.5 mM). All measurements were made two days after treatment. Bars indicate standard error (n = 4). Values with the same letter are not significantly different at P < 0.05
Cd ( µ g g – 1 DW ) 0 50 100 150 200 a c c b b c 0 50 100 150 200 0 200 400 600 800 c c c b b c b a A B C - CdCl2 + CdCl2 d d d Cd ( µ g g – 1 DW ) Cd ( µ g g -1 DW ) a Temperature (oC) 15/13 25/20 35/30
Fig. 3 Effect of temperatures on Cd concentrations in the third leaves (A), the second leaves (B), and roots (C) of rice seedlings treated with or without CdCl2(0.5 mM). Cd concentrations were determined 2 days after treatment. Bars indicate standard error (n = 4). Values with the same letter are not significantly different at P < 0.05
0 20 40 60 80 100 120 140 160 180 A * * ** ** ** Time (h) 0 4 8 12 24 36 48 0 20 40 60 80 100 120 140 160 180 C * * 0 20 40 60 80 100 120 140 160 180 B * * * - CdCl2 + CdCl2 Cd ( µ g g –1 DW ) Cd ( µ g g –1 DW ) Cd ( µ g g –1 DW )
Fig. 4 Changes in Cd concentrations in the second leaves of rice seedlings treated with or without CdCl2(0.5 mM) at 35/30°C (A), 25/20°C (B), and 15/13°C (C). Bars indicate standard error (n = 4). Asterisks represent values that are significant between –CdCl2 and + CdCl2 treatments at P < 0.05
Fluridone treatment
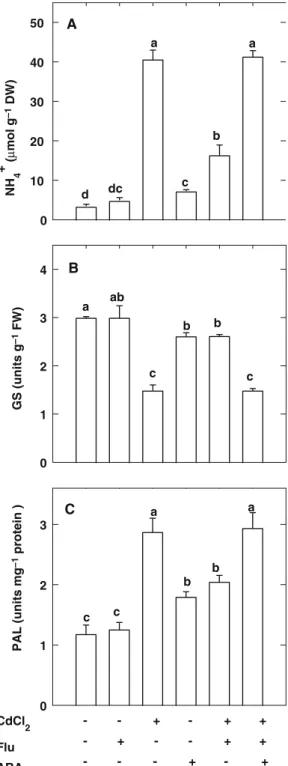
The role of ABA in Cd-induced increase in NH4+ content at high temperature was tested further by using Flu, which blocks the conversion of phyto-ene to phytofluphyto-ene in cartenoid biosynthesis pathway (Kowalczyk-Schro¨der and Sandmann 1992). Flu treatment was observed to inhibit the increase in ABA content caused by CdCl2 (Fig. 7A) and reduce Cd-induced accumulation of NH4+(Fig.8A), decrease in GS activity (Fig.8B), and increase in PAL specific activity (Fig.8C) in the second leaves of rice seedlings grown at high temperature. The effects of Flu on reducing Cd-induced increase in NH4+ content and PAL spe-cific activity, and the decrease in GS activity can be reversed by the application of ABA (Figs. 8A–C). ABA alone was also found to be
able to increase NH4+ content and PAL specific activity and to decrease GS activity in the second leaves of rice seedlings grown at high temperature when compared with untreated control (Fig-s.8A–C). Figs.7B, C also show that Cd content in the second leaves and transpiration rate of rice seedlings treated with Flu plus CdCl2 at high temperature are similar to those with CdCl2 alone.
Discussion
The interactive effects of temperature and heavy metal stress (Cd and Pb) on the growth and some biochemical compounds have been studied in wheat seedlings (O¨ ncel et al.2000). In general, Cd toxicity (reduction of plant height, decrease in chlorophyll content, and accumulation of pheno-lic compounds) of wheat seedlings increases in parallel to temperature increases (O¨ ncel et al. 2000). In rice seedlings, Cd-induced biomass reduction and NH4+ accumulation are also in parallel to temperature increases (Figs.1A, B and 2A). The high biomass reduction and NH4+ accu-mulation caused by CdCl2 at high temperature seem to be a result of more and rapid Cd uptake from the medium (Figs.3A–C and4A–C). It has been shown that enhanced temperature increased the uptake of Cd in Solanum nigrum (Macek et al. 1994), Lemna minor (Chawla et al. 1991), Lolium perenne (Hooda and Alloway1993), and mosses (Mautsoe and Beckett1996). The massive difference (7-fold) in Cd uptake in the leaves of rice seedlings grown at high and medium tem-peratures suggests that the uptake of Cd in rice seedlings is also temperature-dependent. In the present study, rice seedlings were exposed to different air temperatures. In our system, high air temperature treatment resulted in high root temperature in about 1 h after treatment (unpublished data). A great body of evidence has demonstrated that Cd is taken by the roots via essential metal transporters (Cohen et al. 1998; Korshunova et al.1999; Lasat et al. 2000; Pence et al. 2000; Salt et al. 1995). It is not known whether high root temperature increases the activities of metal transporters or Cd uptake in rice roots in the presence of Cd. Further research in this direction is likely to be highly rewarding.
ABA ( nmol g –1 DW) 0 1 2 3 4 - CdCl2 + CdCl2 c c b bc bc a Transpiration rate ( mg H 2 O seedling –1 day –1 ) 0 20 40 60 80 100 120 140 160 a a a b c d A B Temperature (oC) 15/13 25/20 35/30
Fig. 5 Effect of temperatures on transpiration rates (A) and (B) ABA contents in the second leaves of rice seedlings treated with or without CdCl2(0.5 mM). ABA contents were determined 2 days after treatment. Bars indicate standard error (n = 4). Values with the same letter are not significantly different at P < 0.05
It has been shown that the decrease in GS activity and the increase on PAL activity are responsible for NH4+ accumulation in Cd-treated rice leaves (Hsu and Kao 2004). Here, we also show that on treatment with CdCl2, the decrease in GS activity and the increase in PAL specific activity and NH4+content are more pronounced in the leaves of rice seedlings grown at high tem-perature than those at medium and low temper-atures (Figs.2B, C). 0 5 10 20 40 NH 4 ( µ mol g –1 DW ) 0 10 20 30 ABA ( nmol g –1 DW) 0 5 10 15 20 25 c c b a d b a d A B e e ABA (µM ) +
Fig. 6 Effect of ABA concentration on the content of ABA (A) and NH4+ (B) in the second leaves of rice seedlings grown at medium temperature. All measure-ments were made two days after treatment. Bars indicate standard error (n = 4). Values with the same letter are not significantly different at P < 0.05 Cd ( µ g g –1 DW ) 0 50 100 150 ABA ( nmol g –1 DW) 0 1 2 3 4 Flu CdCl 2 - + - + - - + + c c b a b b a a A B Transpiration rate ( mg H 2 O seedling –1 day –1 ) 0 50 100 150 200 250 C a a b b
Fig. 7 Effect of fluridone (Flu, 0.2 mM) on the contents of ABA (A) and Cd (B) in the second leaves of rice seedlings, and the transpiration rates (C) of the rice seedlings treated with or without CdCl2(0.5 mM) at 35/ 30°C. All measurements were made 2 days after treat-ment. Bars indicate standard error (n = 4). Values with the same letter are not significantly different at P < 0.05
At low temperature, CdCl2treatment resulted in an increase in NH4
+
content, but had no effect on GS activity and PAL specific activity in the second leaves (Figs.2A–C). Thus, Cd-induced NH4
+
accumulation in rice seedlings at low
temperature cannot be explained by GS and PAL. Ammonium ion is produced during nitrate assimilation, deamination of amino acids, and photorespiration (Miflin and Lea1976). Thus, the possibilities that nitrate reduction, deamination of amino acid, and photorespiration may be re-sponsile for Cd-induced NH4+accumulation at low temperature cannot be excluded.
ABA is a sesquiterpenoid derived from xan-thophylls (Nambara and Marion-Poll 2005). The level of ABA in plants increases upon their exposure to Cd stress (Fediuc et al. 2005; Hol-lenbach et al.1997; Hsu and Kao2003; Poschen-rieder et al. 1989). Here, we show that ABA accumulation in rice leaves is induced by Cd at high temperature (Fig.5B). However, high tem-perature itself does not cause ABA accumulation in rice seedlings leaves (Fig.5B). It is now well established that ABA in higher plants is derived from C40-carotenoids (Nambara and Morion-Poll, 2005). As Flu is an inhibitor of carotenoid bio-synthesis (Kowalczyk-Schro¨der and Sandmann 1992), the effect of this inhibitor on Cd-induced ABA in the leaves of rice seedlings at high tem-perature may imply that ABA biosynthetic pathway in response to Cd at high temperature appears to be the same as that established in other stress conditions (Nambara and Marion-Poll2005; Zeevaart and Creelman1988).
The present study indicated that ABA is in-volved in Cd-induced NH4+ accumulation in the leaves of rice seedlings grown at high temperature. This conclusion was based on observations that (a) the increase in endogenous ABA content in re-sponse to Cd in leaves of rice seedlings grown at high temperature was more pronounced than that at medium and low temperatures (Fig.5B); (b) exogenous application of ABA at medium tem-perature increased ABA and NH4+contents in the second leaves (Figs.6A, B); (c) Flu treatment
NH 4 + ( µ mol g –1 DW) 0 10 20 30 40 50 a a dc c b CdCl 2 Flu ABA - - + - + + - + - - + + - - - + - + GS (units g –1 FW) 0 1 2 3 4 PAL (units mg –1 protein ) 0 1 2 3 a b c c a a b c c d A B ab b b C
Fig. 8 Effect of fluridone (Flu, 0.2 mM) and ABA (5 lM) on the contents of NH4+(A), and the activities of GS (B) and the specific activities PAL (C) in the second leaves of rice seedlings treated with or without CdCl2(0.5 mM) at 35/30°C. All measurements were made 2 days after treatment. Bars indicate standard error (n = 4). Values with the same letter are not significantly different at P < 0.05
reduced ABA content, as well as Cd-induced accumulation of NH4
+
and changes of the activities of enzymes responsible for NH4+accumulation in the leaves of rice seedlings grown at high tem-perature (Figs.7A and8A–C); and (d) the effects of Flu on Cd-induced NH4+accumulation and en-zyme activities related to NH4+ accumulation of rice seedlings grown at high temperature can be reversed by the application of ABA (Figs. 8A–C). The fact that Flu treatment reduced ABA content, but did not reduce Cd content in leaves of rice seedlings in response to CdCl2at high tem-perature (Figs. 7A, B) suggests that Flu effect on the reduction of Cd-induced NH4+accumulation is attributable to reduction of ABA but not Cd content in leaves of rice seedlings grown at high temperature. These results strengthen further our conclusion that Cd-induced NH4+accumulation at high temperature is mediated through ABA. Re-cently, Fediuc et al. (2005) reported that ABA mediated the Cd-induced stimulation of O-acet-ylserine (thiol) lyase (OASTL), the enzyme responsible for cysteine biosynthesis. However, in case of growth of Arabidopis (root length and seedlings fresh weight), the magnitude of Cd-in-duced inhibition in ABA-insensitive mutants was generally comparable to that in the wild type (Sharma and Kumar 2002). Accumulation of proline in response to Cd has been reported in wheat and Silene vulgaris (O¨ ncel 2002; Schat et al. 1997). We also reported that NH4+accumulation in dark-treated and water-stressed rice seedlings is associated with proline accumulation (Yang and Kao 2000). Our unpublished data indicated that Cd-induced proline accumulation in leaves of rice seedlings grown at high temperature is also med-iated through ABA accumulation.
Cd is known to inhibit transpiration rate of several plants (Hagemeyer et al. 1986; Lamo-reaux and Chaney1978; Schlegel et al.1987). We also found that CdCl2 treatment reduced tran-spiration rate of rice seedlings (Fig.5A). Reduc-tion of transpiraReduc-tion rate and increase in Cd content caused by CdCl2 in seedlings grown at high temperature were more pronounced than those at medium and low temperatures (Figs.3B and 5A). Since Flu treatment had no effect on transpiration rate of rice seedlings in response to CdCl2 at high temperature (Fig. 7C). It appears
that the decrease of transpiration rate caused by CdCl2 is not attributable to increase in ABA content, but to increase in Cd content.
Acknowledgements This work was supported by a re-search grant from the National Science Council of the Republic of China.
References
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein uti-lizing the principle of protein-dye binding. Anal Bio-chem 72:248–254
Chawla G, Singh J, Wiswanathan PN (1991) Effect of pH and temperature on the uptake of cadmium by Lemna minor L. Bull Environ Contam Toxicol 47:84–90 Cohen CK, Fox TC, Garvin DF, Kochian LV (1998) The
role of iron-deficiency stress responses in stimulating heavy-metal transport in plants. Plant Physiol 116:1063–1072
Fediuc E, Lips SH, Erdei L (2005) O-Acetylserine (thiol) lyase activity in Phragmites and Typha plants under cadmium and NaCl stress conditions and the involvement of ABA in the stress response. J Plant Physiol 162:865–872
Foy CD (1998) Plant adaptation to acid, aluminum-toxic soils. Comm Soil Sci Plant Anal 19:959–987
Greger M, Johansson M (1992) Cadmium effect on leaf transpiration of sugar beet (Beta vulgaris). Physiol Plant 86:465–473
Hagemeyer J, Kahle M, Breckle SW (1986) Cadmium in Fagus sylvatica L. trees and seedlings: leaching, up-take and interconnection with transpiration. Air Soil Pollut 29:347–359
Hahlbrock R, Grisebach H (1979) Enzymic controls in the biosynthesis of lignin and flavonoids. Annu Rev Plant Physiol 30:105–130
Hollenbach B, Schreiber L, Hartung W, Dietz KJ (1997) Cadmium tends to stimulate expression of lipid transfer protein (ltp) in barley: implications for the involvement of LTP in wax assembly. Planta 203:9–19 Hooda PS, Alloway BJ (1993) Effects of time and tem-perature on the bioavailability of Cd and Pb from sludge-amended soils. J Soil Sci 44:97–110
Hsu YT, Kao CH (2003) Accumulation of ammonium ion in cadmium tolerant and sensitive cultivars of Oryza sativa. Plant Growth Regul 39:271–276
Hsu YT, Kao CH (2004) Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul 42:227–238 Hung KT, Kao CH (2005) Hydrogen peroxide is required
for abscisic acid-induced NH4+ accumulation in rice leaves. J Plant Physiol 162:1022–1029
Hyodo H, Fujinami H (1989) The effect of 2,5-norbornadiene on the induction of the activity of 1-aminocyclopropane-1-carboxylate synthase and of phenylalanine ammonia-lyase in wounded mesocarp tissue of Cucurbita maxima. Plant Cell Physiol 30:857–860
Javis SC, Jones LHP, Hopper MJ (1976) Cadmium uptake from solution by plants and its transport from roots to shoots. Plant Soil 44:179–191
Kirkham MB (1978) Water relations of cadmium-treated plants. J Environ Qual 7:334–336
Korshunova YO, Eide D, Clark WG, Guerinot ML, Pa-krasi HB (1999) The IRT1 protrein from Arabidopsis thaliana is a metal transporter with a broad substrate range. Plant Mol Biol 40:37–44
Kowalczyk-Schro¨der S, Sandmann G (1992) Interaction of fluridone with phytoene desaturation of Aphano-capsa. Pestic Biochem Physiol 42:7–12
Lamoreaux RL, Chaney WR (1978) The effect of cad-mium on net photosynthesis, transpiration and dark respiration of excised sliver maple leaves. Physiol Plant 43:231–236
Lasat MM, Pence NS, Garvin DF, Ebbs SD, Kochian LV (2000) Molecular physiology of zinc transport in the Zn hypraccumulator Thlaspi caerulescens. J Exp Bot 342:71–79
Lin CC, Kao CH (1996) Disturbed ammonia assimilation is associated with growth inhibition of roots in rice seed-lings caused by NaCl. Plant Growth Regul 18:223–238 Macek T, Kotrba P, Suchova M, Skacel F, Demnerova K, Ruml T (1994) Accumulation of cadmium by hairy-root cultures of Solnum nigrum. Biotechnol Lett 16:621–624
Mautsoe PJ, Beckett (1996) A preliminary study of the factors affecting the kinetics of cadmium uptake by the liverwort Dumortiera hirsute. South Afr J Bot 62:332–336
Miflin BJ, Lea PJ (1976) The pathway of nitrogen assim-ilation in plants. Phytochemistry 15:873–885
Nambara E, Marion-Poll A (2005) Abscisic acid biosyn-thesis and catabolism. Annu Rev Plant Biol 56:165–185 Oaks A, stolen J, Jones K, Winspear MJ, Booesel IL (1980) Enzymes of nitrogen assimilation in maize roots. Planta 148:477–484
O¨ ncel I, Keles Y, u¨stu¨n AS (2000) Interactive effects of temperature and heavy metal stress on the growth and some biochemical compounds in wheat seedlings. Environ Pollut 107:315–320
Pence NS, Larsen PB, Ebbs SD, Letham DL, Lasat MM, Garvin DF, Eide D, Kochian LV (2000) The molec-ular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc Natl Acad Sci USA 97:4956–4960
Peng S, Huang J, Sheehy JE, Laza RC, Visperas RM, Zhong X, Centeno GS, Khush GS, Cassman KG
(2004) Rice yields decline with higher night temper-ature from global warming. Proc Natl Acad Sci USA 101:9971–9975
Poschenrieder C, Gunse B, Barcelo J (1989) Influence of cadmium on water relations, stomatal resistance and abscisic acid content in expanding bean leaves. Plant Phsyiol 90:1365–1371
Rauser WE, Dunmbroff EB (1981) Effect of excess cobalt, nickel and zinc on water relation of Phaseolus vulga-ris. Envion Exp Bot 21:249–255
Razel RA, Ellis S, Singh S, Lewis NG, Towers GHN (1996) Nitrogen recycling in phenylpropanoid metabolism. Phytochemistory 41:31–35
Sakurai N, Katayama Y, Yamaya T (2001) Overlapping expression of cytosolic glutamine synthetase and phenylalanine ammonia-lyase in immature leaf blades of rice. Physiol Plant 113:400–408
Salt DE, Prince RC, Pickering IJ, Raskin I (1995) Mech-anism of cadmium mobility and accumulation in In-dian mustard. Plant Physiol 109:1427–1433
Sanita´ di Toppi L, Gabbrielli R 1999 Response to cad-mium in higher plants. Environ Exp Bot 41:105–130 Schat H, Sharma SS, Vooijs R (1997) Heavy
metal-in-duced accumulation of free proline by a metal-toler-ant and a nontolermetal-toler-ant ecotype of Silene vulgaris. Physiol Plant 101:477–482
Schlegel H, Godbold DL, Hu¨ttermann (1987) Whole plant aspects of heavy metal induced changes in CO2 up-take and water relations of spruce (Picea abies) seedlings. Physiol Plant 69:265–270
Seo M, Koshiba T (2002) Complex regulation of ABA biosynthesis. Trends Plant Sci 7:41–48
Sharma SS, Kumar V (2002) Responses of wild type and abscisic acid mutants of Arabidopisis thaliana to cad-mium. J Plant Physiol 159:1323–1327
van Heerden PS, Towers GHN, Lewis NG (1996) Nitrogen metabolism in lignifying Pinus taeda cell cultures. J Biol Chem 271:12350–12355
Walker-Simmons M (1987) ABA levels and sensitivity in developing wheat embryos of sprouting resistant and susceptible cultivars. Plant Physiol 84:61–66
Weatherburn MW (1967) Phenol-hypochloride reaction for determination of ammonia. Anal Chem 39:971–974 Yang CW, Kao CH (2000) Ammonium in relation to
praline accumulation in detached rice leaves. Plant Growth Regul 30:139–144
Zeevaart JAD, Creelman RA (1988) Metabolism and physiology of abscisic acid. Annu Rev Plant Physiol Plant Mol Biol 39:439–473