Copyright
°c2002 IUBMB 1521-6543/02 $12.00 + .00 DOI: 10.1080/15216540290114586
Research Communication
Effects of Epidermal Growth Factor and Linoleic Acid on Lipid Contents in Human Intestinal C2BBe1 Cells
Chiu-Fang Liang,
1Jane C.-J. Chao,
1Shiaw-Min Hwang,
2and Ya-Hui Tsai
31
Graduate Institute of Nutrition and Health Sciences, Taipei Medical University, 250 Wu Hsing Street, Taipei, Taiwan 110, R.O.C.
2
Food Industry Research and Development Institute, 331 Shih-Pin Road, Hsinchu, Taiwan 300, R.O.C.
3
School of Nutrition and Health Sciences, Taipei Medical University, 250 Wu Hsing Street, Taipei, Taiwan 110, R.O.C.
Summary
Epidermal growth factor (EGF) was reported to regulate triacyl glycerol synthesis in various cells. Linoleic acid and its metabolites were thought to modulate the signal transduction of growth factors.
This study determined whether linoleic acid regulated the effect of EGF on lipid contents in human intestinal C2BBe1 cells. Conflu- ent cells were incubated with serum-free medium (control), EGF (45 ng/mL), linoleic acid (42 µg/mL), or combined EGF (45 ng/mL) and linoleic acid (42 µg/mL) for 48 h. The results showed EGF and linoleic acid significantly increased intracellular cholesterol and triglyceride levels compared with the control and combined groups.
EGF was a more potent stimulator for triacyl glycerol synthesis in C2BBe1 cells than linoleic acid. However, intracellular cholesterol and triglyceride levels did not differ between the control and com- bined groups. The secretion of cholesterol and triglyceride into the medium by C2BBe1 cells did not differ among four groups. Both EGF and linoleic acid strongly stimulated the expression of EGF receptor mRNA in C2BBe1 cells at 48 h compared with the control and combined groups. Therefore, EGF and linoleic acid increased triacyl glycerol synthesis in C2BBe1 cells through stimulating the expression of EGF receptor mRNA. The effect of EGF and linoleic acid on this lipogenesis was reversed in the presence of both EGF and linoleic acid by downregulating the expression of EGF receptor mRNA.
IUBMB
Life , 54: 275–279, 2002
Keywords EGF receptor; epidermal growth factor; human intestinal cells; linoleic acid.
Received 12 July 2002; accepted 25 September 2002.
Address correspondence to Jane C.-J. Chao, Taipei Medical Uni- versity, Graduate Institute of Nutrition and Health Sciences, 250 Wu Hsing Street, Taipei, Taiwan 110, R.O.C. Fax: +886-2-2737-3112;
E-mail: [email protected]
INTRODUCTION
There is evidence that linoleic acid and its metabolites mod- ulate the signal transduction of growth factors, such as epi- dermal growth factor (EGF), insulin, and insulin-like growth factor-1, in vitro (1–8). Previous studies showed that linoleic acid and its eicosanoid metabolites enhanced and sustained the mitogenic activity of EGF in fibroblasts and mammary epithe- lial cells (1–6). Additionally, EGF-stimulated biosynthesis of 13-hydroxyoctadecadienoic acid, the major lipoxygenase prod- uct, in Syrian hamster embryo fibroblasts was blocked by the inhibitors of EGF receptor tyrosine kinase activity, suggesting that EGF induction of the linoleic acid pathway was linked to the activation of the EGF receptor (3).
Epidermal growth factor, distributed widely in a variety of body fluid in the mammals (9), is secreted by the salivary glands, gastric mucosa, and duodenum (10). It was reported that EGF regulated the secretion and formation of lipids in various cells (11–13). The secretion of triglycerides in human colon adeno- carcinoma Caco-2 cells was stimulated by EGF (11). However, EGF decreased the intracellular accumulation of triglycerides in hamster auricular sebocytes (12). The stimulation of triacyl glycerol synthesis by insulin in primary cultures of adult rat hep- atocytes was also suppressed by EGF through the inhibition of malic enzyme activity (13).
The C2BBe1 cell line, cloned from human colon adenocar- cinoma Caco-2 cells, has high homology in morphology com- pared with Caco-2 cells that was often used as an experimental model for the synthesis, absorption, or transport of macronutri- ents, minerals, vitamins, or drugs in the intestine (14, 15). It is unclear whether linoleic acid also serves a regulatory function in the action of EGF on lipogenesis in the intestinal cells. There- fore, the purpose of this study investigated the effects of EGF
275
and linoleic acid on lipid levels and EGF receptor expression in human intestinal C2BBe1 cells.
EXPERIMENTAL PROCEDURES Cell Line and Culture
Human intestinal C2BBe1 cells (CCRC 60182, Culture Col- lection and Research Center, Food Industry Research and Devel- opment Institute, Hsinchu, Taiwan, R.O.C.) were grown in 90%
Dulbecco’s modified Eagle’s medium (DMEM), containing 10%
fetal bovine serum, 10 µg/mL transferrin, and 1 mM sodium pyruvate at 37
◦C in a humidified CO
2incubator (95% air and 5% CO
2). Upon approximately 80–90% conflunency, the cells were subcultured using 0.25% trypsin and 0.02% EDTA. Prior to the addition of the treatment, the cells were grown to 100%
confluency, overgrown for another 7 days, and synchronized by incubating in the basal medium (100% DMEM) for 24 h.
The cells were then incubated with recombinant human EGF (45 ng/mL; 7.5 nM) (16), linoleic acid (42 µg/mL; 150 µM) (17) in physiological concentrations, or combined EGF (45 ng/mL) and linoleic acid (42 µg/mL) for 48 h. Conjugated 2 moles of α-linoleic acid with 1 mole of bovine serum albumin (BSA) (Sigma L9530, Sigma-Aldrich, Inc., St. Louis, MO) was added to the medium to increase its water solubility. Each treatment was also added an equal amount (100 mg/mL) of BSA.
The cells and conditioned media were collected by triplet at 48 h. The cells were washed three times with phosphate-buffered saline (PBS), resuspended in PBS containing 100 µL/10
7cells protease inhibitors [4-(2-aminoethyl)-benzenesulfonyl fluoride, pepstatin A, transepoxysuccinyl-
L-leucylamido (4-guanidino) butane, bestatin, leupeptin, and aprotinin, Sigma P8340, Sigma- Aldrich, Inc.], and ultrasonicated for 5–10 s to break the mem- brane for further analyses.
Endogenous EGF Secretion
Prior to the addition of the treatment, endogenous EGF secre- tion into the conditioned medium was measured at 450 nm (18) by a commercial EGF immunoassay kit (Quantikine, DEG00, Research and Diagnostics Systems, Inc., Minneapolis, MN). The serum-free conditioned medium (200 µL) was incubated with EGF antibody coated in a 96-well plate for 2 h at room temper- ature, washed 3 times with wash buffer, and then incubated with 200 µL polyclonal EGF antibody conjugated to horseradish per- oxidase for 1 h. After several washes, samples were incubated with 200 µL substrate (tetramethylbenzidine:H
2O
2= 1:1) for 20 min. The reaction was terminated by 50 µL of 1 M sulfu- ric acid. EGF levels were determined at 450 nm and corrected at 540 nm using an ELISA reader (Multiskan RC, Labsystems, Helsinki, Finland).
Lipid Analyses
Total cholesterol and triglyceride levels in C2BBe1 cells and the conditioned medium were measured spectrophotometrically
at 490 nm (U2000 double-beam spectrophotometer, Hitachi, Tokyo, Japan) by cholesterol oxidase/p-aminophenazone (CHOD-PAP) and glycerol-3-phosphate oxidase/p-amino- phenazone (GPO-PAP) methods, respectively, using commer- cial kits (Randox Laboratories Ltd., Antrim, UK). Total lipid levels were measured spectrophotometrically at 540 nm using a commercial kit (Randox Laboratories Ltd.). Cholesterol and triglyceride levels were expressed as the percentage of total lipids.
RNA Extraction and Quantitation
The cells (10
7) were lysed by 1 mL TRIZOL reagent (Life Technologies GIBCO BRL, Taipei, Taiwan, R.O.C.), and mixed with 200 µL chloroform. After centrifugation at 12,000 × g for 15 min at 4
◦C, RNA was separated in the upper aqueous phase.
The aqueous phase containing RNA was mixed with 500 µL of cold 100% isopropanol for 10 min. The RNA pellet was then washed with 1 mL of cold 75% ethanol, air dried, and rehy- drated in 10 to 20 µL of 0.1% diethylpyrocarbonate-water. The concentration and purity of total cellular RNA were determined spectrophotometrically by the absorbance at 260 nm and by the ratio of the absorbance at 260 nm to that at 280 nm, respectively.
Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
The level of EGF receptor mRNA was determined by RT- PCR. Total cellular RNA (5 µg) as a template mixed with 0.5 µg oligo(dT)
12−18was heated at 70
◦C for 10 min, and chilled on ice at least for 1 min. The RNA and primer mixture was incubated with 2 µL of 10 × PCR buffer, 2 µL of 25 mM MgCl
2, 1 µL of 10 mM dNTP mix, and 2 µL of 0.1 M dithiothreitol at 42
◦C for 5 min, and then incubated with 200 units of S
UPERS
CRIPTII reverse transcriptase (Life Technologies GIBCO BRL) at 42
◦C for 50 min. The reaction was terminated at 70
◦C for 15 min.
The mixture was incubated with RNase H at 37
◦C for 20 min prior to the amplification of the target cDNA. The cDNA (2 µL) was incubated with 1 × PCR buffer, 1.5 mM MgCl
2, 0.2 mM dNTP mix, 1.25 units Taq DNA polymerase (Life Technologies GIBCO BRL), and 0.2 µM primer of EGF receptor or β-actin (Life Technologies GIBCO BRL) at 94
◦C for 30 s, 55
◦C for 30 s, and 72
◦C for 60 s for 40 cycles. The 20-mer sequence of EGF receptor primer was 5
0- ACCAGAGTGATGTCTGGAGC- 3
0for sense strand and 5
0-GATGAGGTACTCGTCGGCAT-3
0for antisense strand. The 20-mer sequence of β-actin primer was 5
0-CGGATGTCCACGTCACACTT-3
0for sense strand and 5
0-GTTGCTATCCAGGCTGTGCT-3
0for antisense strand. The PCR product for EGF receptor and β-actin was 367 and 469 bp, respectively. The amplified products were separated by 2%
agarose gel electrophoresis, and visualized by staining with
ethidium bromide under ultraviolet light. The bands were quanti-
tated by an image analysis system (Gel analysis system,
EverGene Biotechnology, Taipei, Taiwan, R.O.C.) and Phoretix
1D Lite software (Phoretix International Ltd., Newcastle upon
Tyne, UK).
Statistical Analysis
Data are expressed as the mean ± SD. All data were analyzed by Statistical Analysis System (SAS) software (v6.12, SAS In- stitute Inc., Cary, NC). Two-way ANOVA and Fisher’s least significant difference test were used to determine the effect of EGF and linoleic acid on lipid levels at different times. Statistical significance was assigned at the 0.05 level.
RESULTS
Endogenous EGF Secretion and Lipid Levels
Endogenous EGF secretion into the conditioned medium of C2BBe1 cells was 0.12 ± 0.01 pg/mL. Because the secretion of cholesterol into the medium by C2BBe1 cells was unde- tectable in all groups at 24 h, lipid levels were measured after incubation for 48 h. Intracellular levels of cholesterol (Fig. 1A)
Figure 1. Intracellular (A) and secreted (B) cholesterol levels in C2BBe1 cells after the addition of 45 ng/mL recombinant hu- man EGF, 42 µg/mL linoleic acid, or combined EGF (45 ng/mL) and linoleic acid (42 µg/mL) for 48 h. Without EGF or linoleic acid was the control group. Data are means ± SD (n = 3). Means not sharing a letter differ significantly ( P < 0.05) by Fisher’s least significant difference test.
Figure 2. Intracellular (A) and secreted (B) triglyceride levels in C2BBe1 cells after the addition of 45 ng/mL recombinant hu- man EGF, 42 µg/mL linoleic acid, or combined EGF (45 ng/mL) and linoleic acid (42 µg/mL) for 48 h. Without EGF or linoleic acid was the control group. Data are means ± SD (n = 3). Means not sharing a letter differ significantly ( P < 0.05) by Fisher’s least significant difference test.
and triglyceride (Fig. 2A) in the EGF and linoleic acid groups significantly increased (P < 0.05) compared with the control and combined groups at 48 h. The EGF group had significantly higher intracellular levels of cholesterol (Fig. 1A) and triglyc- eride (Fig. 2A) than the linoleic acid group ( P < 0.05). However, intracellular cholesterol and triglyceride levels did not differ be- tween the control and combined groups. Additionally, the secre- tion of cholesterol (Fig. 1B) and triglyceride (Fig. 2B) into the medium by C2BBe1 cells did not differ in all groups.
EGF Receptor mRNA Expression
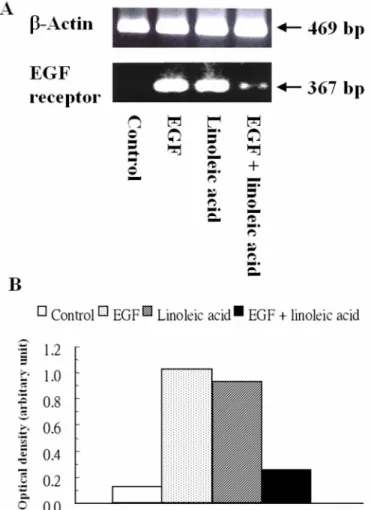
The level of EGF receptor mRNA determined by RT-PCR
was undetectable in the control group at 48 h (Fig. 3A). The
expression of EGF receptor mRNA was strongly stimulated to
7- to 8-fold of the control in the EGF and linoleic acid groups
(Fig. 3B). However, the level of EGF receptor mRNA was only
twofold of the control in the combined group.
Figure 3. The expression of EGF receptor mRNA determined by RT-PCR (A) and quantitated by the optical density ratio of EGF receptor to β-actin (B) in C2BBe1 cells after the addition of 45 ng/mL recombinant human EGF, 42 µg/mL linoleic acid, or combined EGF (45 ng/mL) and linoleic acid (42 µg/mL) for 48 h in C2BBe1 cells. Without EGF or linoleic acid was the control group. Samples were obtained from three independent experiments (n = 3), and pooled together for analysis. The pu- rified total cellular RNA (5 µg) isolated from freshly harvested C2BBe1 cells was used as a template for reverse transcription by S
UPERS
CRIPTII reverse transcriptase. The synthesized cDNA was then amplified by PCR. The amplified products were sepa- rated by 2% agarose gel electrophoresis and visualized by ethid- ium bromide staining. The 469- and 367-bp positions represent β-actin and EGF receptor cDNA, respectively.
DISCUSSION
Endogenous EGF secretion was much lower than exogenous EGF concentration, indicating that the influences of EGF re- sulted from the addition of EGF. Our data showed that both EGF and linoleic acid stimulated triacyl glycerol synthesis in C2BBe1 cells. A previous study found that EGF could enhance cholesterol synthesis via the stimulation of HMG CoA reductase
activity (19). Additionally, the incubation with EGF at a concen- tration of 1 nM (6 ng/mL) at least for 4 days significantly in- creased intracellular triglyceride content in mouse differentiated adipocytes (3T3-L1 cells) by stimulating acyl-CoA synthetase activity, which is the rate-limiting enzyme for adipogenesis (20).
Because confluent Caco-2 monolayers primarily express intesti- nal fatty acid-binding protein for the uptake of long chain fatty acids (14), exogenous linoleic acid can serve as a substrate for lipid synthesis. Similarly, van Greevenbroek et al. demonstrated that the addition of
14C-linoleic acid was efficiently incorporated into triglycerides by Caco-2 cells (17). Our results showed nei- ther EGF nor linoleic acid significantly affect the secretion of cholesterol and triglyceride into the medium by C2BBe1 cells.
However, Levy et al. found that the addition of EGF (25, 50, and 100 ng/mL) significantly enhanced the release of triglyceride- rich lipoprotein by 25–40% in human fetal intestinal explants (21). More triglyceride was secreted into the medium during in- cubation with 1.0 mM linoleic acid than with 1.0 mM palmitic acid in Caco-2 cells (17).
Combined EGF and linoleic acid treatment did not affect in- tracellular levels and the secretion of cholesterol and triglyceride in C2BBe1 cells at 48 h. Contrast to the increases in intracellular cholesterol and triglyceride levels by EGF or linoleic acid treat- ment alone, it is suggested that the addition of linoleic acid may inhibit the action of EGF on lipid contents in C2BBe1 cells.
Our data demonstrated that both EGF and linoleic acid
strongly stimulated the expression of EGF receptor mRNA in
C2BBe1 cells at 48 h by RT-PCR analysis. Similar to our re-
sults, the addition of EGF increased the expression, the binding
capacity, and tyrosine phosphorylation of the EGF receptor in
confluent rat intestinal (IEC-6) cells (22). Previous studies found
that 13(S)-hydroperoxyoctadecadienoic acid, a specific lipoxy-
genase metabolite of linoleic acid, stimulated EGF-dependent
mitogenesis and upregulation of EGF-dependent tyrosine phos-
phoryaltion by inhibiting the dephosphorylation of the EGF re-
ceptor in Syrian hamster embryo fibroblasts (23, 24). It is specu-
lated that the stimulation of EGF receptor expression by linoleic
acid is regulated by the metabolite of linoleic acid. However,
combined EGF and linoleic acid obviously decreased the ex-
pression of EGF receptor mRNA compared with EGF or linoleic
acid alone, which was consistent with the findings of intracel-
lular lipid levels. It is suggested that both EGF and linoleic
acid may stimulate lipogenesis in C2BBe1 cells through mod-
ulating the expression of EGF receptor mRNA. The expres-
sion of EGF receptor mRNA was downregulated in the pres-
ence of both EGF and linoleic acid, resulting in an inhibition
of lipogenesis in C2BBe1 cells. However, EGF was found to
stimulate the conversion of exogenous linoleic acid to 13(S)-
hydroxyoctadecadienoic acid in Syrian hamster embryo fibro-
blasts through the activation of the EGF receptor tyrosine kinase
(25). Additionally, the proliferation of mammary epithelial cells
induced by EGF was enhanced and sustained by linoleic acid
and its eicosanoid metabolites (3). These results suggest that
combined EGF and linoleic acid may have a synergistic effect
on EGF-dependent signal transduction through the amplification of signals from the EGF receptor, which conflicted with the in- hibitory effect of linoleic acid on EGF-modulated lipid contents in C2BBe1 cells. Therefore, it is presumed that linoleic acid and/or its metabolites as signal transducers up- or downregulate the expression of the EGF receptor in different cell types.
In conclusion, EGF and linoleic acid increased intracellu- lar cholesterol and triglyceride levels in C2BBe1 cells through stimulating the expression of EGF receptor mRNA. However, combined EGF and linoleic acid reversed the action on lipogen- esis in C2BBe1 cells through downregulating the expression of EGF receptor mRNA.
ACKNOWLEDGEMENTS
The study was supported by the National Science Council of Taiwan, R.O.C. (grant NSC87-2413-B-038-007). We want to thank the fellows at the Culture Collection and Research Center in Food Industry Research and Development Institute for the technical assistance.
REFERENCES
1. Eling, T. E., and Glasgow, W. C. (1991) Transduction of the epidermal growth factor mitogenic signal in BALB/c 3T3 fibroblasts involves linoleic acid metabolism. Adv. Prostaglandin Thromboxane Leukot. Res. 21B, 843–
846.
2. Glasgow, W. C., Afshari, C. A., Barrett, J. C., and Eling, T. E. (1992) Mod- ulation of the epidermal growth factor mitogenic response by metabolites of linoleic and arachidonic acid in Syrian hamster embryo fibroblasts. Dif- ferential effects in tumor suppressor gene (+) and (−) phenotypes. J. Biol.
Chem. 267, 10771–10779.
3. Bandyopadhyay, G. K., Hwang, S., Imagawa, W., and Nandi, S. (1993) Role of polyunsaturated fatty acids as signal transducers: amplification of signals from growth factor receptors by fatty acids in mammary epithelial cells. Prostaglandins Leukot. Essent. Fatty Acids 48, 71–78.
4. Eling, T. E., Everhart, A. L., and Glasgow, W. C. (1995) Modulation of EGF cell signaling tyrosine phosphorylation by linoleic acid metabolites. Adv.
Prostaglandin Thromboxane Leukot. Res. 23, 453–455.
5. Eling, T. E., Everhart, A. L., Angerman-Stewart, J., Hui, R., and Glasgow, W. C. (1997) Modulation of epidermal growth factor signal transduction by linoleic acid metabolites. Adv. Exp. Med. Biol. 407, 319–322.
6. Park, Y., Allen, K. G., and Shultz, T. D. (2000) Modulation of MCF-7 breast cancer cell signal transduction by linoleic acid and conjugated linoleic acid in culture. Anticancer Res. 20, 669–676.
7. Meuillet, E. J., Leray, V., Hubert, P., Leray, C., and Cremel, G. (1999) Incor- poration of exogenous lipids modulates insulin signaling in the hepatoma cell line, HepG2. Biochim. Biophys. Acta 1454, 38–48.
8. Lanson, M., Besson, P., and Bougnoux, P. (1997) Supplementation of MCF-7 cells with essential fatty acids induces the activation of pro- tein kinase C in response to IGF-1. J. Lipid Mediat. Cell Signal. 16, 189–197.
9. Cohen, S. (1962) Isolation of a mouse submaxillary gland protein acceler- ating incisor eruption and eyelid opening in the newborn animals. J. Biol.
Chem. 237, 1555–1562.
10. Kelly, E. J., Newell, S. J., Brownlee, K. G., Farmery, S. M., Cullinane, C., Reid, W. A., Jackson, P., Gray, S. F., Primrose, J. N., and Lagopoulos, M.
(1997) Role of epidermal growth factor and transforming growth factor alpha in the developing stomach. Arch. Dis. Child. Fetal Neonatal Ed. 76, F158–F162.
11. Murthy, S., Mathur, S. N., Bishop, W. P., and Field, F. J. (1997) Inhibition of apolipoprotein B secretion by IL-6 is mediated by EGF or an EGF-like molecule in Caco-2 cells. J. Lipid Res. 38, 206–216.
12. Sato, T., Imai, N., Akimoto, N., Sakiguchi, T., Kitamura, K., and Ito, A. (2001) Epidermal growth factor and 1alpha, 25-dihydroxyvitamin D3 suppress lipogenesis in hamster sebaceous gland cells in vitro. J. Invest.
Dermatol. 117, 965–970.
13. Yoshimoto, K., Nakamura, T., and Ichihara, A. (1983) Reciprocal effects of epidermal growth factor on key lipogenic enzymes in primary cultures of adult rat hepatocytes. Induction of glucose-6-phosphate dehydrogenase and suppression of malic enzyme and lipogenesis. J. Biol. Chem. 258, 12355–
12360.
14. Levy, E., Mehran, M., and Seidman, E. (1995) Caco-2 cells as a model for intestinal lipoprotein synthesis and secretion. FASEB J. 9, 626–635.
15. Mehran, M., Levy, E., Bendayan, M., and Seidman, E. (1997) Lipid, apolipoprotein, and lipoprotein synthesis and secretion during cellular dif- ferentiation in Caco-2 cells. In Vitro Cell Dev. Biol. Anim. 33, 118–128.
16. Chang, C.-J., and Chao, J. C.-J. (2002) Effect of human milk and epider- mal growth factor on growth of human intestinal Caco-2 cells. J. Pediatr.
Gastroenterol. Nutr. 34, 394–401.
17. van Greevenbroek, M. M., Voorhout, W. F., Erkelens, D. W., van Meer, G., and de Bruin, T. W. (1995) Palmitic acid and linoleic acid metabolism in Caco-2 cells: different triglyceride synthesis and lipoprotein secretion.
J. Lipid Res. 36, 13–24.
18. Abe, Y., Sagawa, T., Sakai, K., and Kimura, S. (1987) Enzyme-linked im- munosorbent assay (ELISA) for human epidermal growth factor (hEGF).
Clin. Chim. Acta 168, 87–95.
19. Singh-asa, P., Waters, M. J., and Wilce, P. A. (1985) A mechanism for the in vitro stimulation of adrenal cortisol biosynthesis by epidermal growth factor. Int. J. Biochem. 17, 857–862.
20. Adachi, H., Kurachi, H., Homma, H., Adachi, K., Imai, T., Morishige, K., Matsuzawa, Y., and Miyake, A. (1994) Epidermal growth factor promotes adipogenesis of 3T3-L1 cell in vitro. Endocrinology 135, 1824–1830.
21. Levy, E., Thibault, L., and M´enard, D. (1992) Intestinal lipids and lipopro- teins in the human fetus: modulation by epidermal growth factor. J. Lipid Res. 33, 1607–1617.
22. Polk, D. B., McCollum, G. W., and Carpenter, G. (1995) Cell density- dependent regulation of PLC gamma I tyrosine phosphorylation and cat- alytic activity in an intestinal cell line (IEC-6). J. Cell Physiol. 162, 427–433.
23. Glasgow, W. C., Hui, R., Everhart, A. L., Layawickreme, S. P., Angerman- Stewart, J., Han, B. B., and Eling, T. E. (1997) The linoleic acid metabolite, (13S)-hydroperoxyoctadecadienoic acid, augments the epidermal growth factor receptor signaling pathway by attenuation of receptor dephosphory- lation. Differential response in Syrian hamster embryo tumor suppressor phenotypes. J. Biol. Chem. 272, 19269–19276.
24. Hui, R., Kameda, H., Risinger, J. I., Angerman-Stewart, J., Han, B., Barrett, J. C., Eling, T. E., and Glasgow, W. C. (1999) The linoleic acid metabo- lite, 13-HpODE augments the phosphorylation of EGF receptor and SHP-2 leading to their increased association. Prostaglandins Leukot. Essent. Fatty Acids 61, 137–143.
25. Glasgow, W. C., Hill, E. M., McGown, S. R., Tomer, K. B., and Eling, T. E. (1996) Regulation of 13(S)-hydroxyoctadecadienoic acid biosynthe- sis in Syrian hamster embryo fibroblasts by the epidermal growth factor receptor tyrosine kinase. Mol. Pharmacol. 49, 1042–1048.