早發性停經患者的 DAZL 基因單一核甘酸多型性分析
計畫類別: 個別型計畫
計畫編號: NSC93-2314-B-006-078-
執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立成功大學醫學系婦產科
計畫主持人: 郭保麟
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 94 年 10 月 27 日
研究計劃年度報告:早發性停經患者的 DAZL 基因單一核苷酸多形性分析
93-2314-B-006-078 計劃主持人:郭保麟
Haplotypes of DAZL Are Associated with Susceptibility to Spermatogenic Failure in the
Taiwanese Population
Yen-Ni Teng1, Yung-Ming Lin3, Ying-Hung Lin5, Wan-Ching Tsai3, Hsiao-Fang Sunny Sun6, Pei-Yang Hsu4, Chia-Ling Chung 4, Ting-Yi Kuo2, Pao-Lin Kuo4
Financial Support: This study was supported by grants from the National Science Council of the Republic of China (NSC- 91-2314-B-006-149, NSC 91-3112-B-006-008, NSC 92-3112-B-006-002, NSC 93-3112-B-006-004, and NSC 93-2314-B-006-078).
Departmental Affiliations: 1Department of Early Childhood Education and Nursery,
2Biotechnology Department, Chia Nan University of Pharmacy and Science, Tainan, Taiwan;
3Department of Urology, 4Department of Obstetrics and Gynecology, 5Graduate Institute of
Basic Sciences, and 6Institute of Molecular Medicine, National Cheng Kung University College of Medicine, Tainan, Taiwan
RUNNING TITLE: Haplotype of DAZL
Reprint Requests: Pao-Lin Kuo, MD, Division of Genetics, Department of Obstetrics and
Gynecology, National Cheng Kung University Hospital, 138 Sheng-Li Road, Tainan, Taiwan 704 (Fax: 886-6-276-6185; E-mail: [email protected])
Key Words: single nucleotide polymorphism, DAZL gene, spermatogenic failure, haplotype,
linkage disequilibrium
ABSTRACT
To investigate the role of DAZL gene in human spermatogenesis, DAZL was genotyped in 231 men with spermatogenic defects and in 191 fertile men (control group). Five single nucleotide polymorphisms (SNPs) were analyzed: 260A>G (Thr12→Ala), 386A>G (Thr54→
Ala), 520+34c>a, 584+28c>t, and 796+36g>a, the latter three of which were novel. There was a significant difference in the allelic frequency for the 386A>G DAZL gene
polymorphism between infertile men and the control group (P < 0.0001). Three major haplotypes constituted 88.0% of the patient population. Three major haplotypes also constituted 87.0% of the control population. The linkage disequilibrium (LD) map patterns were different for infertile men and control subjects. Of all haplotypes studied, five
demonstrated significant differences in frequency between infertile men and control subjects (P < 0.05). Some haplotypes were over-transmitted in patients with severe spermatogenic failure, while some haplotypes were under-transmitted in these patients. It appears that the over-transmitted haplotyes conferred susceptibility to spermatogenic failure, while
under-transmitted haplotypes were protective against spermatogenic failure. All deleterious haplotypes belonged to minor haplotypes in the fertile men (in combination making up less than 20% of the population), possibly due to negative selection. These results suggested an important role for the DAZL gene in human spermatogenic failure. The functional
significance of each haplotype and the distribution of these haplotypes in different ethnic
groups deserves further investigation.
Introduction
Between 2% and 12% of couples worldwide are affected by reduced fertility. In roughly half of these cases the defect can be traced to the man (World Health Organization, 1992). A large proportion of infertile men have insufficient sperm production or impaired
spermatogenesis (spermatogenic failure). Spermatogenesis is a complex developmental process in which male germ cells progress through mitotic proliferation, two rounds of
meiotic divisions, and finally undergo dramatic morphological changes to form mature sperm.
The DNA content of the final product is half that of the progenitor cells. This process involves a large portion of the genome of an organism to ensure the quality of the final products. It is estimated that mutations in any of up to 11% of all genes in Drosophila might lead to male sterility (Hackstein et al. 2000). This is also likely to be true for humans, considering the extremely high incidence of infertility in men (De Krester et al. 1997).
Genetic factors have been associated with impaired sperm production in humans. Screening of the long arm of the human Y chromosome has detected Yq11.2 microdeletion in 5-15% of males with spermatogenic failure. It is believed that many genes located in Yq11.2 are azoospermia factors (AZFs) (Reijo et al. 1996a; Vogt et al. 1996; Lahn and Page 1997; Pryor et al. 1997; Simoni et al. 1998). Other genetic factors involved in spermatogenic failure include HLA-haplotypes, mutations at the mitochondrial DNA polymerase locus, a polymorphism of the cytochrome P4501A1 gene, mutations of the follicle-stimulating hormone (FSH) receptor gene, the FATE gene, and the synaptonemal complex protein 3 gene
(Trannainen et al. 1997; Fritsche et al. 1998; Tsujimura et al. 1999; Rovio et al. 2001; Olesen et al. 2003; Miyamoto et al. 2003). The association between short CAG repeat expansion in
X-linked androgen receptor genes and the risk of impaired spermatogenesis remains
uncertain (Patrizio and Leonard 2001). In a previous study, we identified an autosomal DAZL gene SNP, -386 A>G, which confers susceptibility to spermatogenic failure in the
Taiwanese population. The goal of the present study was to identify novel DAZL SNPs and to investigate whether specific haplotypes of DAZL were associated with male infertility in the Taiwanese population. We identified eight haplotypes that were over- or under-transmitted in men with severe spermatogenic failure. These findings suggested an important role for DAZL in human spermatogenesis.
Materials And Methods
Subjects
From January 2001 to June 2004, we studied a total of 281 consecutive, unselected infertile men presenting with severe oligozoospermia or non-obstructive azoospermia. One hundred ninety-one fertile men were enrolled as controls. All study and control subjects belonged to Han Taiwanese, the major ethnic group in Taiwan (making up more than 95%
of the country’s population). The control subjects were recruited from husbands of women who received regular prenatal care at the University Hospital. All of the control subjects had fathered at least 2 children within 5 years without assisted reproductive technologies. The experimental design was in accord with the Helsinki Declaration of 1975 on human experimentation, and was approved by the “insert institution name here” ethics review board. Signed informed consent was obtained from all enrollees. The recruitment of human subjects and evaluation of patients was performed according to our previous studies (Lin et al. 2002a; Teng et al. 2002). In brief, all patients underwent comprehensive characterization, including a detailed history, physical examination, at least 2 semen analyses, endocrinology profile testing (luteinizing hormone (LH), FSH, prolactin (PRL), and testosterone (T)), karyotyping, and a molecular test for Y-chromosome microdeletions. Severe
oligozoospermia was defined as a sperm count less than 5 × 106/ml. Patients with highly suspected non-obstructive azoospermia were advised to undergo bilateral testicular biopsies.
Non-obstructive azoospermia was defined as either spermatogenic defects in the testicular biopsy (such as hypospermatogenesis, maturation arrest, and Sertoli cell-only syndrome), or as elevated serum FSH, total testicular volume less than 30 ml, and no other applicable diagnoses. Semen analysis was performed according to standard methods (WHO 1992).
Serum levels of FSH, LH, PRL, and T were measured using commercial radioimmunoassay kits: Coat-A-Count FSH IRMA, Coat-A-Count LH IRMA, Coat-A-Count PRL IRMA, and IMMULITE Total Testosterone (Diagnostic Products Corp.). Chromosome analysis was performed using the GTG method (G-banding by Trypsin-Giemsa technique). Molecular analysis of Y-chromosome microdeletions included a combination of 16 gene-based primers, as described previously (Lin et al. 2002a). All control subjects also underwent a Y
chromosome deletion test. Control subjects with abnormal karyotypes, deletions of the Y chromosome, or other recognizable causes of male infertility were excluded from
genotyping for DAZL.
Single Strand Conformation Polymorphism (SSCP) And Sequence Analysis
A total of 112 patients were subjected to SSCP analysis for DAZL gene polymorphism screening. Genomic DNA was extracted from peripheral blood samples using a Puregene DNA isolation kit (Gentra Systems, Inc.). To amplify DAZL exons, PCR reactions were performed in 20 μl volumes containing 200 ng of genomic DNA, 10 mM Tris-HCl (pH 8.3),
50 mM KCl, 0.1% Triton X-100, 200 μM dNTPs, 100 pmol of each primer, and 1 U Taq
DNA polymerase (Promega). The primers and PCR conditions for different DAZL exons have been described previously (Teng et al. 2002). PCR analyses were performed in an automated thermal cycler (OmniGene Thermal Cycler, Hybaid Ltd.). The PCR products were mixed with an equal volume of formamide buffer (95% formamide, 10 mM EDTA, 0.1% bromophenol blue, 0.1% xylene cyanol). The mixtures were denatured for 5 min at 95°C and cooled rapidly on ice for 1 min. For each sample, 5 µl mixtures were subjected to SSCP analysis using GeneGel Excel gels as recommended by the manufacturer (Pharmacia Biotech). Prior to analysis by SSCP, all PCR products were sequenced to assure there was no cross-amplification with DAZ. After SSCP analysis, PCR products with aberrant
band-shifts were sequenced to identify mutations or polymorphisms. Sequence analysis was performed with an automatic sequencer (ABI 377, Applied Biosystems/PE).
Genotyping
Primers (sequences available on request) were designed using the Primer 3 program (Rozen and Skaletsky 2000). Each set of primers was selected to cover the entire coding sequence, the exon-intron junctions, and the 5'- and 3'- untranslated regions. The amplicons were amplified in a multiplex fashion and each 20 µl reaction consisted of 50 ng of genomic DNA, 10 pmol of primers, 4.0 mM MgCl2, 0.2 mM deoxynucleotide triphosphates, and 0.5 U
of AmpliTaq Gold DNA polymerase (Applied Biosystems). The cycling conditions were:
95°C for 5 min, followed by 35 cycles at 95°C for 1 min, 55°C for 1 min, and 72°C for 1 min with a final extension for 10 min at 72°C. Each allele was measured by primer extension and SNaPshot chemistry (Applied Biosystems). Multiplex PCR products (3 µl) were treated with 2.5 U of shrimp alkaline phosphatase (SAP, Amersham Pharmacia Biotech) and 2 U of exonuclease I (New England Biolabs) in a 10 μl reaction volume for 1 h at 37°C. SNaPshot multiplex PCR was performed in a 10 μl reaction volume containing 1.25 μl of SNaPshot Multiplex Ready Reaction Mix (Applied Biosystems), 4 μl of SAP/ exonuclease I–treated
PCR products, and 1 to 3 pmol of each SNaPshot primer. Thermal cycling for SNaPshot reactions consisted of 25 cycles at 96°C for 10 sec, 50°C for 5 sec, and 60°C for 30 sec. The SNaPshot multiplex reaction was then treated by SAP in an 11 μl reaction volume containing 10 μl of SNaPshot multiplex PCR product and 1 U of SAP. The samples were analyzed using
an ABI 3100 genetic analyzer (Applied Biosystems) and the allele determination was carried out with the Genotyper 3.7 program (Applied Biosystems).
Statistical Analysis
Tests for association with single markers and haplotypes in control samples were performed using a Chi-square test or Fisher’s exact test. A P-value <0.05 was considered statistically significant. The relative risk of spermatogenic failure was estimated from
logistic odds ratios (OR) and 95% CI in multivariate analysis. The Hardy–Weinberg equilibrium was calculated, in accordance with standard procedures, by use of χ2 analysis.
Tests for haplotype association with spermatogenic failure were performed using EHPLUS software (Xie and Ott 1993), and statistical significance was estimated using the
permutation test PMPLUS (Zhao et al. 2000). Associations between haplotypes and male infertility were analyzed using linear regression models. The LD coefficient (D') between each pair of SNPs was calculated by use of the ldmax program within the GOLD software package (Abecasis and Cookson 2000).
Results
Patient Characteristics
Of 281 patients, 24 showed abnormal karyotypes and 26 were found to have deletions in Y chromosome genes. All patients with gross karyotypic abnormalities and deletions in Y chromosome genes were excluded. Of the 231 patients included in this study, 43 showed severe oligozoospermia, and 188 had non-obstructive azoospermia. None of the control subjects was found to have deletions in Y-chromosome genes.
Identification of SNPs by SSCP and DNA Sequencing Analysis
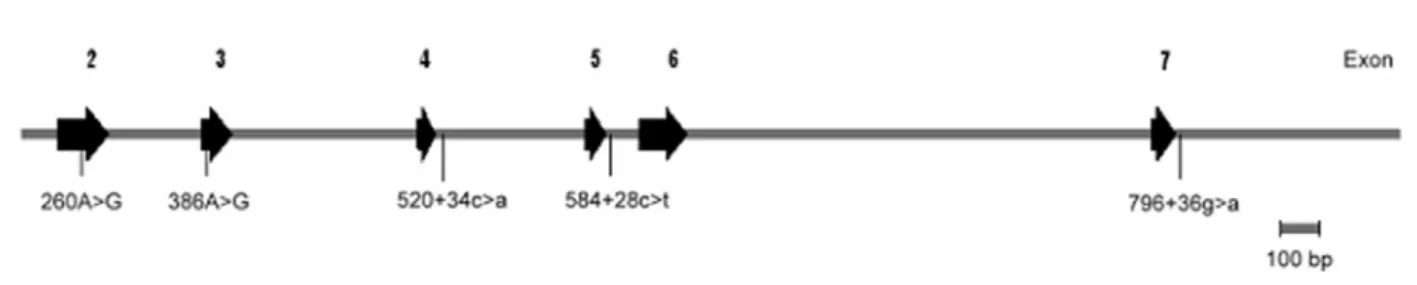
Alterations in conformation were detected as electrophoretic mobility differences in polyacrylamide gels. Of 11 exons examined, abnormal SSCP patterns were observed with exon 2-, 3-, 4-, 5-, and 7-containing DNA fragments. The DNA fragments with aberrant bands were sequenced to identify molecular lesions. Five SNPs were identified in 112 patients: 260A>G (Thr12→Ala) in exon 2, 386A>G (Thr54→Ala) in exon 3, 520+34c>a in
intron 4, 584+28c>t in intron 5, and 796+36g>a in intron 7. Of the five SNPs, two have been previously described: 260A>G in exon 2 and 386A>G in exon 3 (Teng et al. 2002).
Locations of the five SNPs are shown in Figure 1.
Association of SNPs With Susceptibility to Spermatogenic Failure
We used the five SNPs for genotyping of all enrollees. The allelic frequencies of the five SNPs are presented in Table 1. Although the nucleotides in 584+28 and 796+36 are
cytosine and guanine, respectively, in the GenBank database (accession number U77471 and U77472, respectively), thymidine and adenine occurred more frequently in these two positions for the Han Taiwanese (Table 1). The results of the Hardy–Weinberg equilibrium test are shown in Table 2. All markers were in Hardy-Weinberg equilibrium (p < 0.05). Of the five SNPs, only the 386 A>G polymorphism was significantly associated with severe spermatogenic failure (P<0.0001), with an odds ratio of 11.667 (Table 2).
Linkage Disequilibrium Between Different SNPs
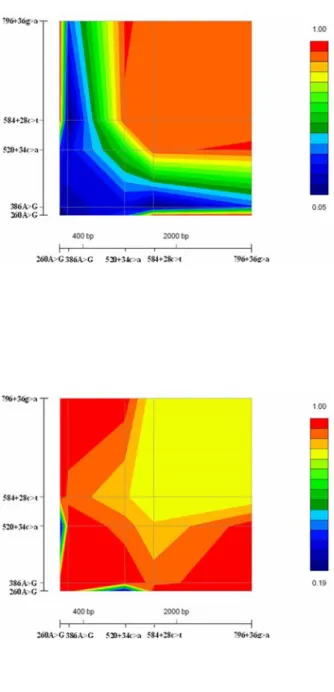
The pairwise disequilibrium measures (D′) between alleles at polymorphic loci were calculated. For patients with spermatogenic failure, 260A>G was found to be in complete LD with 584+28c>t and 796+36g>a (D′=1). SNP 520+34c>a had strong LD with 584+28c>t and 796+36g>a (D′ = 0.931 and 0.945 respectively). SNP 386A>G did not show LD with
260A>G, 520+34c>a, 584+28c>t, or 796+36g>a (Table 3). For control subjects, 260A>G was found to be in complete LD with 386A>G, 584+28c>t and 796+36g>a (D′=1). Strong linkage disequilibrium was observed for 386A>G with 260A>G, 520+34c>a, 584+28c>t, and 796+36g>a (D′>0.926) (Table 3). The LD map of infertile men was different from control subjects (fig. 2).
Haplotype Analysis
The major haplotype for spermatogenic failure was 11111 (accounting for 45.8% of the population), followed by 11122 (17.8%), 11211 (15.6%), 11222 (8.7%), and 22222 (4.7%). The frequencies of other haplotypes were less than 2% (data not shown). As shown in Table 4, the major haplotypes for the control subjects were 11122 (41.7 %), 11211 (30.3
%), and 11111 (14.9 %). The frequencies for other haplotypes were less than 2% (data not shown). Of all haplotypes, five showed significant differences in frequency between infertile men and control subjects (Table 4). Haplotypes 11111, 11222, and 22222 were over-transmitted in patients with severe spermatogenic failure, while haplotypes 11122 and 11211 were under-transmitted in these patients.
Discussion
In the present study, we confirmed our previous finding that the 386A>G allele of DAZL confers susceptibility to severe spermatogenic failure in the Han Taiwanese population. We also identified three novel SNPs that were not associated with spermatogenic failure.
Haplotypes including 386A>G do not constitute major haplotypes of DAZL because the allelic frequency of 386A>G was rare in both infertile men (5.97%) and control subjects (0.54%, see Table 1). The major haplotypes for cases with spermatogenic failure were 11111 (45.8%), 11122 (17.8%), 11211 (15.6%), and 11222 (8.7%). These haplotypes accounted for 88.0% of the patient population. The major haplotypes for control subjects were 11122 (41.7%), 11211 (30.3%), and 11111 (14.9%), which, in combination, accounted for 87.0% of the control population. It appeared that haplotypes 11111, 11222, and 22222 conferred susceptibility to spermatogenic failure, while haplotypes 11122 and 11211 were protective against spermatogenic failure.
In humans, the DAZ gene family consists of three members: BOULE, DAZL and DAZ.
BOULE is the ancestor gene and is conserved from flies to humans (Xu et al. 2001). In
Drosophila, boule is required for posttranscriptional regulation of gene expression in male germ cells and is considered to be a meiotic regulator. Loss-of-function mutations in boule resulted in meiotic arrest of the male fly (Eberhart et al. 1996). In human testis, BOULE
protein expression is restricted to the period from the leptotene stage to the late spermatocyte stage (Xu et al. 2001). The absence of BOULE expression in the spermatocytes of patients with meiotic arrest suggested a role for human BOULE in spermagoenesis (Luetjens et al. 2004). However, neither a mutation nor a clinically
significant BOULE sequence variant could be identified in infertile men vs. controls (Xu et al. 2003; Luetjens et al. 2004; Lepretre et al., 2004).
It was believed that DAZL was generated by a transposition of its ancestor gene,
BOULE, from chromosome 2 to chromosome 3 (Xu et al. 2001). The DAZL protein is
expressed in the germ cells of diverse species. In Caenorhabditis elegans, disruption of dazl causes meiotic arrest in oogenesis (Karashima et al. 2000). In Xenopus, Xdazl is required for early primordial germ cell differentiation and is indirectly necessary for the migration of primordial germ cells through the endoderm (Houston and King 2000; Houston et al.1998).
In mice, loss of function of the DAZL homologue leads to loss of germ cells in both males and females (Ruggiu et al. 1997). Immunohistochemical studies in human testis reveal that
DAZL is expressed in many compartments from spermatogonia and meiotic spermatocytes
to mature spermatozoa (Lin et al. 2001; Lin et al. 2002b). We have previously reported significantly lower transcriptional levels of DAZL in men with spermatogenic failure (Lin et al. 2001; Kuo et al. 2004). The results of the present study clearly demonstrate the important
role of DAZL in human spermatogenesis.
The third member of the DAZ gene family, DAZ, is located on the Y chromosome. It is highly homologous to DAZL, with 83% similarity in the coding region of the cDNA (Saxena et al. 1996; Shan et al. 1996; Yen et al. 1996; Chai et al. 1997). Aside from humans, only the Y chromosomes of great apes and Old World monkeys contain DAZ orthologs. Other
mammals have only the autosomal BOULE and DAZL genes (Cooke et al. 1996; Reijo et al.
1996b; Eberhart et al. 1996; Houston et al. 1998; Agulnik et al. 1998; Xu et al. 2001). It is believed that the DAZ gene arose 40 million years ago, during primate evolution, from the transposition, repeat amplification, and pruning of DAZL (Saxena et al. 1996). The DAZ protein was expressed in spermatogonia, spermatocytes, and in round spermatids to mature sperm (Habermann et al. 1998; Reijo et al. 2000). Normal men usually have four copies of
DAZ (Kuroda- Kawaguchi et al. 2001). Complete deletion of the DAZ gene cluster accounts
for ~10% of cases of men with spermatogenic failure (Silber et al. 1998; Ferlin et al. 1999;
Krausz et al. 1999; Lin et al. 2002a). However, other AZFs in the Y chromosome AZFc region are usually deleted along with DAZ (Kuroda- Kawaguchi et al. 2001). It is, therefore, very difficult to define the role of the DAZ gene in human spermatogenesis based on cases with complete deletions of the DAZ gene cluster. There are two types of partial deletions of
DAZ gene cluster, and both types of deletions had a modest effect on reproductive fitness.
The first type-a 1.6-Mb deletion-also occurs in about 2% of the American population (Repping et al. 2003). The second type-a 1.8 Mb deletion-was described in North Eurasia.
Intriguingly, the negative effect of the 1.8 Mb deletion could be counterbalanced by another genetic factor-possibly a Y-linked factor (Repping et al. 2004). The presence of partial
deletions in both fertile men and men with impaired sperm production suggests that DAZ functions are affected by as yet unidentified genetic factors.
Several functions of the DAZ gene family are known. All DAZ family members encode a highly conserved RNA binding motif (Xu et al. 2001). In Drosophila, Boule is found to regulate the transition from G2 to M phase during meiosis via control of Twine (Cdc25) translation (Maines and Wasserman 1999). Several messenger RNAs have been shown to be substrates for the DAZL protein, including Cdc25C, Cdc25A, Pam, GRSF1, and TRF2 transcripts (Venables et al. 2001; Jiao et al. 2002). Homology between BOULE and
DAZL/DAZ is limited to the RNA binding motif and the DAZ repeat region, which share 78
and 50% nucleotide sequence similarity. Human BOULE and Xenopus Xdazl can partially rescue the phenotype of a Drosophila boule mutant. Likewise, when human DAZ and DAZL
transgenes were introduced into Dazl-null mice, partial rescue of the mutant phenotype was observed (Houston et al. 1998; Slee et al. 1999; Vogel et al. 2002, Xu et al. 2003). These findings suggest a high degree of functional conservation between DAZ family members.
Because of this partial conservation of function, DAZ family members are likely to share similar sets of interacting proteins and RNA substrates and they are likely to serve similar functions.
DAZL SNP 386A>G is located within the RNA-binding motif. It is likely that the
386A>G substitution significantly reduces the binding of substrates critical for cell cycle regulation (Teng et al. 2002). Interestingly, SNP 386G has not been identified in Caucasian populations (Bartoloni et al. 2004; Tschanter et al., 2004; Becherini et al., 2004). The 260A>G polymorphism is not located in a critical protein domain and is most likely a true variant. In the Han Taiwanese, SNP 260G follows Mendelian inheritance laws while in the Caucasian population, SNP 260G does not (Tschanter et al. 2004). It seems that significant differences in genetic background exist between the Han Taiwanese and Caucasians. Three novel SNPs, -520+34c>a, 584+28c>t, and 796+36g>a, -are located within introns.
Though SNPs 520+34c (a) and 796+36g (a) are located in proximity to the splice donor site, substitution of cytosine by adenine or guanine by adenine, respectively, did not seem affect splicing. SNP 584+28c (t) is located in the pyrimidine tract of intron 5. Substitution of cytosine by thymidine did not affect splicing. It is not known whether these mutations acted as intronic splicing enhancers or silencers, or whether they affected mRNA stability
(Nissim-Rafinia et al. 2002). It is intriguing that of the haplotypes without the 386G allele,
11111 and 11222 had a moderate negative impact on reproductive fitness while 11122 and 11211 were protective against spermatogenic failure. These findings indicate that substitution of a single, seemingly unimportant variant in a haplotype may significantly alter gene
function. Although different haplotype functions may be determined by the combinatorial effect of variants, it seems more likely that haplotypes 11111, 11122, 11211, and 11222 are linked, in cis or in trans, to factors that modify DAZL functions. Given the evidence of partial functional conservation between DAZ family members, it is tempting to hypothesize that BOULE, DAZL and DAZ operate in a partially complementary or synergistic manner during
human spermatogenesis. Therefore, both BOULE and DAZ are potential trans modifiers of
DAZL. Because only 16.8 kb of genomic sequence encompassing DAZL was analyzed in this study, the potential cis –acting locus surrounding DAZL remains to be identified.
It will be important to investigate whether the 386A>G allele is restricted to certain ethnic groups, considering the lack of 386G allele in the Caucasian population (Bartoloni et al. 2004; Tschanter et al., 2004; Becherini et al., 2004). The haplotype 22222 included the 386G allele and was more prevalent in cases with spermatogenic failure. In a previous study, we demonstrated that the 386G allele was not expressed in testicular tissue (Teng et al.
2002). It is quite likely that haplotypes 11111, 11222, 22222 are associated with suboptimal function of DAZL while haplotypes 11122 and 11211 are associated with optimal function
of DAZL. In cases with spermatogenic failure, 260A>G was found to be in complete LD with 584+28c>t and 796+36g>a (D′=1). The 520+34c>a allele showed strong LD with 584+28c>t and 796+36g>a (D′> 0.931). However, 386A>G was not linked with other alleles. Paradoxically, 386A>G had strong LD with other sequence variants in control subjects. Different LD maps for fertile and infertile men suggest distinct genetic
backgrounds for these two groups of subjects. It is likely that 386G is a deleterious mutation arising recently in certain ethic groups, and the development of certain testicular phenotypes depends on the subject’s genetic background. Of the deleterious haplotypes, those
containing the 386A allele (11111 and 11222) are more interesting than those containing the 386G allele, because of their higher frequency in the Han Taiwanese population. It is noteworthy that all deleterious haplotypes, with or without the 386A>G allele, belonged to minor haplotypes in the fertile men (in combination making up less than 20% of the population), possibly due to negative selection (Bielawski and Yang 2001).
In the present study, we have described DAZL haplotypes occurring in the Han Taiwanese. Although a large number of association studies have led to the discovery of genetic factors for many common diseases, there have been few association studies focused on sterile genes in humans (The International HapMap Corsortium 2003). Y chromosomal haplotypes have been reported to be associated with reproductive fitness in both Drosophila
and humans, however, these haplotypes are not related to regions linked to specific genes (Chippindale and Rice 2001; Krausz et al. 2001). For the first time, we have demonstrated the association of autosomal gene haplotypes with human spermatogenic failure. The functional significance and distribution of these haplotypes in different ethnic groups deserves further investigation.
Acknowledgements
We thank Dr. Cathy S-J Fann, Dr. H-M Lin, and Miss C-C Chang for Statistical analysis, Professor R.-B Lu for revision of the manuscript. This study was supported by grants from the National Science Council of the Republic of China (NSC- 91-2314-B-006-149, NSC 91-3112-B-006-008, NSC 92-3112-B-006-002, NSC 93-3112-B-006-004, and NSC 93-2314-B-006-078).
Electronic-Database Information
URLs for data presented herein are as follows:
GenBank, http://www.ncbi.nlm.nih.gov/Genbank/
GOLD, http://www.sph.umich.edu/csg/abecasis/GOLD/ (for graphical overview of linkage disequilibrium)
Primer3, http://www-genome.wi.mit.edu/cgi-bin/primer/ primer3_www.cgi (for primer design)
Online Hardy–Weinberg equilibrium, http://ihg.gsf.de/cgi-bin/hw/hwa1.pl
References
Abecasis GR, Cookson WO (2000) Gold – graphical overview of linkage disequilibrium.
Bioinformatics 16:182-183
Agulnik AI, Zharkikh A, Boettger-Tong H, Bourgeron T, McElreavey K, Bishop CE (1998) Evolution of the DAZ gene family suggests that Y-linked DAZ plays little, or a limited, role in spermatogenesis but underlines a recent African origin for human populations.
Hum Mol Genet 7:1371-1377
Bartoloni L, Cazzadore C, Ferlin AG, Foresta C (2004) Lack of T54A polymorphism of the
DAZL gene in infertile Italian patients. Mol Hum Reprod 10:613-615
Becherini L, Guarducci E, Degl’innocenti S, Rotondi M, Forty G, Krausz C (2004) DAZL polymorphisms and susceptibility to spermatogenic failure: an example of remarkable ethnic differences. Int J Androl 27:375-381
Bielawski JP, Yang Z (2001) Positive and negative selection in the DAZ gene family. Mol Biol Evol 18:523-529
Chai NN, Phillips A, Fernandez A, Yen PH (1997) A putative human male infertility gene
DAZLA: genomic structure and methylation status. Mol Hum Reprod 3:705-708
Chippindale AK, Rice WR (2001) Y chromosome polymorphism is a strong determinant of male fitness in Drosophila melanogaster. Proc Natl Acad Sci 98:5677-5682
Cooke HJ, Lee M, Kerr S, Ruggiu M (1996) A murine homologue of the human DAZ gene
is autosomal and expressed only in the male and female gonads. Hum Mol Genet 5:513-516
De Krester DM (1997) Male infertility. Lancet 349:787-790
Eberhart CG, Maines JZ, Wasserman SA (1996) Meiotic cell cycle requirement for a fly homologue of human Deleted in Azoospermia. Nature 381:783-785
Ferlin A, Moro E, Garolla A, Foresta C (1999) Human male infertility and Y chromosome deletions: role of the AZF-candidate genes DAZ, RBM and DFFRY. Hum Reprod 14:1710-1716.
Fritsche E, Schuppe HC, Dohr O, Ruzicka T, Gleichmann E, Abel J (1998) Increased frequencies of cytochrome P4501A1 polymorphisms in infertile men. Andrologia 30:125-128
Habermann B, Mi HF, Edelmann A, Bohring C, Backert IT, Kiesewetter F, Aumuller G, Vogt PH (1998) DAZ (Deleted in Azoospermia) genes encode protein located in human late spermatid, and in sperm tails. Hum Reprod 13:363-369.
Hackstein JHP, Hochstenbach R, Pearson PL (2000) Towards an understating of the genetics of human male infertility. TIG 16:565-572
Houston DW, Zhang J, Maines JZ, Wasserman SA, King ML (1998) A Xenopus DAZ-like gene encodes an RNA component of germ plasm and is a functional homologue of
Drosophlila boule. Development 125:171-180
Houston DW, King ML (2000) A critical role for Xdazl, a germ plasm-localized RNA, in the differentiation of primordial germ cells in Xenopus. Development 127:447-456
Jiao X, Trifillis P, Kiledjian M (2002) Identification of target messenger RNA substrates for the murine Deleted in Azoospermia-Like RNA binding protein. Biol Reprod 66:475-485 Karashima T, Sugimoto A, Yamamoto M (2000) Caenorhabditis elegans homologue of the
human azoospermia factor DAZ is required for oogenesis but not for spermatogenesis.
Development 127:1069-1079
Krausz C, Bussani-Mastellone C, Granchi S, McElreavey K, Scarselli G, Forti G (1999) Screening for microdeletions of Y chromosome genes in patients undergoing
intracytoplasmic sperm injection. Hum Reprod 14:1717-1721
Krausz C, Quintana-Murci L, Rajpert-De Meyts E, Jorgensen N, Jobling MA, Rosser ZH, Skakkebaek NE, McElreavey K (2001) Identification of a Y chromosome haplogroup associated with reduced sperm counts. Hum Mol Genet 10:1873-1877
Kuo PL, Wang ST, Lin YM, Lin YH, Teng YN, Hsu CC (2004) Expression profiles of the
DAZ gene family in human testis with and without spermatogenic failure. Fertil Steril
81:1034-1040
Kuroda-Kawaguchi T, Skaletsky H, Brown LG, Minx PJ, Cordum HS, Waterston RH, Wilson RK, Silber S, Oates R, Rozen S, Page DC (2001) The AZFc region of the Y chromosome features massive palindromes and uniform recurrent deletions in infertile
men. Nat Genet 29:279-286
Lahn BT, Page DC (1997) Functional coherence of the human Y chromosome. Science 278:675-680
Lepretre AC, Patrat C, Jouannet P, Bienvenu T (2004) Mutation analysis of the BOULE gene in men with non-obstructive azoospermia: identification of a novel polymorphic variant in the black population. Int J Androl 27:301-303
Lin YM, Chen CW, Sun HS, Tsai SJ, Hsu CC, Teng YN, Lin J SN, Kuo P (2001) Expression patterns and transcript concentrations of the autosomal DAZL gene in testes of
azoopsermic men. Mol Hum Reprod 11:1015-1022
Lin YM, Lin YH, Teng YN, Hsu CC, Lin JSN, Kuo PL (2002a) Gene-based screening for Y-chromosome deletions in Taiwanese men presenting with spermatogenic failure.
Fertil Steril 77:897-903
Lin YM, Chen CW, Sun HS, Tsai SJ, Lin J SN, Kuo PL (2002b) Presence of DAZL transcript and protein in mature spermatozoa. Fertil Steril 77:626-629
Luetjens CM, Xu EY, Rejo Pera RA, Kamischke A, Nieschlag E, Gromoll J (2004)
Assocation of meiotic arrest with lack of BOULE protein expression in infertile men. J Clin Endocrinol Metab 89:1926-1933
Maines JZ, Wasserman SA (1999) Post-transcriptional regulation of the meiotic Cdc25 protein Twine by the Dazl orthologue Boule. Nat Cell Biol 1:171-174
Miyamoto T, Hasulke S, Yogev L, Maduro MR, Ishikawa M, Westphal H, Lamb DJ (2003) Azoospermia in patients heterozygous for a mutation in SYCP3. Lancet 362:1714-1719 Nissim-Rafinia M, Kerem B (2002) Splicing regulation as a potential genetic modifier.
Trends Genet 18:123-127
Olesen C, Silber J, Eiberg H, Ernst E, Petersen K, Lindenberg S, Tommerup N (2003) Mutation analysis of the human FATE gene in 144 men. Hum Genet 113:195-201 Patrizio P, Leonard DG (2001) Expansion of the CAG trinucleotide repeats in the androgen
receptor gene and male infertility: a controversial association. J Androl 22:748-749 Pryor JL, Kent-First M, Muallem A, Van Bergen AH, Nolten WE, Meisner L, Roberts KP
(1997) Microdeletions in the Y chromosome of infertile men. N Engl J Med 336:534-539
Reijo R, Alagappan RK, Patrizio P, Page DC (1996a) Severe oligospermia resulting from deletions of spermatogenic failure factor gene on Y chromosome. Lancet
347:1290-1293
Reijo R, Seligman J, Dinulos MB, Jaffe T, Brown LG, Disteche CM, Page DC (1996b) Mouse autosomal homolog of DAZ, a candidate male sterility gene in humans, is expressed in male germ cells before and after puberty. Genomics 35:346-352
Reijo RA, Dorfman DM, Slee R, Renshaw AA, Loughlin KR, Cooke H (2000) DAZ family proteins exist throughout male germ cell development and transit from nucleus to
cytoplasm at meiosis in humans and mice. Biol Reprod 63:1490-1496
Repping S, Skaletsky H, Broan L, van Daalen SKM, Korver CM, Pyntikova T,
Kuroda-Kawaguchi T, de Vries JW, Oates RD, Silber S, van der Veen F, Page DC, Rozen S (2003) Polymorphism for a 1.6-Mb deletion of the human Y chromosome persists through balance between recurrent mutation and haploid selction. Nat Genet 35:247-251 Repping S, van Daalen SKM, Korver CM, Brown LG, Marszalek JD, Gianotten J, Oates RD,
Silber S, van der Veen F, Page DC, Rozen S (2004) A family of Y chromosomes has dispersed through northern Eurasia despite a 1.8-Mb deletion in the azoospermia factor c region. Genomics 83:1046-1052
Rozen S, Skaletsky HJ (2000) Primer3 on the WWW for general users and for biologist programmers. In: Krawetz S, Misener S (eds) Bioinformatics Methods and Protocols:
Methods in Molecular Biology. Humana Press, Totowa, NJ, pp 365-386
Rovio AT, Marchington DR, Donat S, Schuppe HC, Abel J, Fritsche E, Elliott DJ, Laippala P, Ahola AL, McNay D, Harrison RF, Hughes B, Barrett T, Bailey DM, Mehmet D, Jequier AM, Hargreave TB, Kao SH, Cummins JM, Barton DE, Cooke HJ, Wei YH, Wichmann L, Poulton J, Jacobs HT (2001) Mutations at the mitochondiral DNA polymerase (POLG) locus associated with male infertility. Nat Genet 29:261-262
Ruggiu M, Speed R, Taggart M, McKay SJ, Kilanowski F, Saunders P, Dorin J, Cooke HJ (1997) The mouse Dazla gene encodes a cytoplasmic protein essential for
gametogenesis. Nature 389:73-77
Saxena R, Brown LG, Hawkins T, Alagappan RK, Skaletsky H, Reeve MP, Reijo R, Rozen S, Dinulos MB, Disteche CM, Page DC (1996) The DAZ gene cluster on the human Y chromosome arose from an autosomal gene that was transported, repeatedly amplified and pruned. Nat Genet 14:292-299
Shan Z, Hirschmann P, Seebacher T, Edelmann A, Jauch A, Morell J, Urbitsch P, Vogt PH (1996) A SPGY copy homologous to the mouse gene Dazla and the Drosophila gene
boule is autosomal and expressed only in the human male gonad. Hum Mol Genet
5:2005-2011
Silber SJ, Alagappan R, Brown LG, Page DC (1998) Y chromosome deletions in azoospermic and severely oligozoospermic men undergoing intracytoplasmic sperm injection after testicular sperm extraction. Hum Reprod 13:3332-3337.
Simoni M, Kamischke A, Nieschlag E (1998) Current status of the molecular diagnosis of Y-chromosomal microdeletions in the work-up of male infertility. Initiative for
international quality control. Hum Reprod 13:1764-1768
Slee, R., Grimes, B., Speed, R.M. Taggart M, Maguire SM, Ross A, McGill NI, Saunders PTK, Cooke HJ (1999) A human DAZ transgene confers partial rescue of the mouse
Dazl phenotype. Proc Natl Acad Sci USA 96:8040-8045.
Teng YN, Lin YM, Lin YH, Tsao SY, Hsu CC, Lin SJ, Tsai WC, Kuo PL (2002) Association
of a single-nucleotide polymorphism of the Deleted-in-Azoospermia-Like gene with susceptibility to spermatogenic failure. J Clin Endocrinol Metab 87:5258-5264 The International HapMap Consortium (2003) The International HapMap Project. Nature
426:789-796
Trannainen JS, Aittomaki K, Min J, Vaskivuo T, Huhtaniemi IT (1997) Men homozygous for an inactivating mutation of the follicle-stimulating hormone (FSH) receptor gene present variable suppression of spermatogenesis and infertility. 15:205-206
Tschanter P, Kostova E, Luetjens CM, Cooper TG, Nieschlag E, Gromoll J (2004) No association of the A260G and A386G DAZL single nucleotide polymorphisms with male infertility in a Caucasian population. Hum Reprod 19(12):2771-2776
Tsujimura A, Takahara S, Kitamura M, Miura H, Koga M, Sada M, Tsuji T, Matsumiya K, Okuyama A (1999) HLA-DR antigen and HLA-DRB1 genotyping genotyping with non-obstructive azoospermia in Japan. J Androl 20:545-550
Venables JP, Ruggiu M, Cooke HJ (2001) The RNA-binding specificity of the mouse Dazl protein. Nucl Acid Res 29:2479-2483
Vogel T, Speed RM, Ross A, Cook HO (2002) Partial rescue of the Dazl knockout mouse by the human DAZL gene. Mol Hum Reprod 8:797-804
Vogt PH, Edelmann A, Kirsch S, Henegariu O, Hirschmann P, Kiesewetter F, Kohn FM, Schill WB, Farah S, Ramos C, Hartmann M, Hartschuh W, Meschede D, Behre HM,
Castel A, Nieschlag E, Weidner W, Grone HJ, Jung A, Engel W, Haidl G (1996) Human Y chromosome spermatogenic failure factors (AZF) mapped to different subregions in Yq11. Hum Mol Genet 5:933-943
World Health Organization (1992) WHO Laboratory Manual for the Examination of Human Semen and Semen-Cervical Mucus Interaction 3rd ed. Cambridge University Press, Cambridge, UK pp 3-21
Xie X, Ott J (1993) Testing linkage disequilibrium between a disease gene and marker loci.
Am J Hum Genet 53:1107
Xu EY, Moore FL, Reijo Pera RA (2001) A gene family required for human germ cell development evolved from an ancient meiotic gene conserved in metazoans. Proc Natl Acad Sci 98:7414-7419
Xu EY, Lee DF, Klebes A, Turek PJ, Kornberg TB, Reijo Pera RA (2003) Human BOULE gene rescues meiotic defects in infertile flies. Hum Mol Genet 12:169-175
Yen PH, Chai NN, Salido EC (1996) The human autosomal gene DAZLA: testis specificity
and a candidate for male infertility. Hum Mol Genet 5:2013-2017
Zhao J, Curtis D, Sham PC (2000) Model-free analysis and permutation tests for allelic associations. Hum Hered 50:133-139
Table 1
Allelic frequencies of five sequence variants of DAZL in infertile men and control subjects
Allele frequency (%) Genotype frequency (%) SNP Allele Control Spermatogenic
Failure P Genotype Control Spermatogenic
Failure P 260A>G A 96.74 95.97 0.5671 AA 93.48 91.94 .5595
G 3.26 4.03 AG 6.52 8.06
GG 0 0
386A>G A 99.46 94.03 <0.0001 AA 98.91 88.64 <0.0001
G 0.54 5.97 AG 1.09 10.80
GG 0 0.57
520+34c>a C 64.66 58.92 0.3000 CC 38.74 31.90 0.2169
A 35.34 41.08 CA 51.83 54.05
AA 9.42 14.05
584+28c>t T 51.92 52.17 0.9458 TT 27.47 25.54 0.7011
C 48.08 47.82 TC 48.90 53.26
CC 23.63 21.20
796+36g>a A 50.72 50.63 0.9832 AA 27.34 27.22 0.9997
G 49.28 49.37 GA 46.76 46.84
GG 25.90 25.94
NOTE —Allele and genotype frequencies were compared between infertile men and control subjects.
Table 2
Odds ratios for each sequence variant of DAZL Sequence
Variants
Tests For Associationa Allele
(Genotypes) Odds Ratio P C.I.
1/1 vs. 1/2+2/2 1.256 0.55954 0.583-2.704 260A>G
2/2 vs. 1/1+1/2 ** ** **
1/1 vs. 1/2+2/2 11.667 <0.0001* 2.749-103.986*
386A>G
2/2 vs. 1/1+1/2 ** ** **
1/1 vs. 1/2+2/2 1.351 0.16478 0.883-2.065 520+34c>a
2/2 vs. 1/1+1/2 0.636 0.16262 0.336-1.205 1/1 vs. 1/2+2/2 1.104 0.67588 0.694-1.757 584+28c>t
2/2 vs. 1/1+1/2 1.150 0.57709 0.703-1.881 1/1 vs. 1/2+2/2 1.006 0.98106 0.603-1.679 796+36g>a
2/2 vs. 1/1+1/2 0.997 0.99216 0.593-1.678
NOTE —Allele and genotype frequencies were compared between infertile men and control subjects. The common allele is referred to as allele 1 and the rare allele as allele 2. C.I.= 95%
confidence interval.
a Symbols: *, the exact test is considered; **: The frequency of genotype 2/2 is zero.
Pairwise linkage disequilibrium (D′) in infertile men and control subjects
260A>Ga 386A>Ga 520+34c>aa 584+28c>ta 796+36g>aa
260A>G - 0.108 0.288 1.000* 1.000*
386A>G 1.000 - 0.208 0.050 0.290
520+34c>a 0.186 0.999 - 0.931* 0.945*
584+28c>t 1.000* 0.926 0.841* - 0.918*
796+36g>a 1.000 1.000 0.977* 0.792* -
NOTE—Estimates in the upper right are for infertile men and in the lower left are for control subjects.
a * P < 0.05.
Table 4
Haplotypes showing significantly different frequencies between infertile men and control subjects
Haplotype Haplotype Frequency (%) Spermatogenic Failure vs. Controls 260A>G 386A>G 520+34c>a 584+28c>t 796+36g>a Spermatogenic
Failure Controls P Valuea Odds Ratio C.I.b 1 1 1 1 1 45.8 14.9 <0.0001 4.835 3.456 – 6.765 1 1 1 2 2 17.8 41.7 <0.0001 0.299 0.219 - 0.410 1 1 2 1 1 15.6 30.3 <0.0001 0.423 0.304 – 0.591 1 1 2 2 2 8.7 1.1 <0.0001 8.957 3.174 - 25.280 2 2 2 2 2 4.7 1.6 0.0100 3.133 1.257 - 7.810
NOTE — The common allele is referred to as allele 1 and the rare allele as allele 2.
a The P-value was analyzed for statistical significance by Chi-square test and <0.05 was considered statistically significant.
b C.I.= 95% confidence interval.
Figure legends
Figure 1 Genomic structure of the DAZL gene, indicating the locations of the five SNPs assayed.
Figure 2 Pairwise LD D' values of five SNPs across DAZL genome by use of true genomic distance for infertile men (A) and normal controls (B).
Figure 1 Genomic structure of the DAZL gene, indicating the locations of the five SNPs assayed.
A
B
Figure 2 Pairwise LD D' values of five SNPs across DAZL genome by use of true genomic distance for infertile men (A) and normal controls (B).