國立臺灣大學醫學院解剖學暨細胞生物學研究所 碩士論文
Graduate Institute of Anatomy and Cell Biology College of Medicine
National Taiwan University Master Thesis
虎皮蛙視網膜及松果體中的感光細胞
The Photoreceptor Cells in Retina and Pineal Gland of Tiger-skinned Frog
林子純 Tzu-Chuen Lin
指導教授: 錢宗良 博士 Advisor: Chung-Liang Chien, Ph.D.
中華民國 108 年 7 月
July 2019
致謝
時光飛逝,兩年的碩士生活隨著論文的完成也將畫下休止符。猶記得當初剛 確定錄取研究所的興奮之情、一年級和同學們一起撐過的繁重課業、課餘時所辦 的活動及二年級的實驗室生活。兩年後,這些點點滴滴都還歷歷在目,我卻已經 要離開這個地方了。
在這兩年的研究生生活中,幸運的遇到了許多幫助我的人,不論是課業上或 是實驗上,甚至是生活上,任何的幫忙都非常的感激。首先要感謝的是我的指導 教授 錢宗良老師,在課業以及研究上讓我獲益良多,更增進了我對於邏輯思考 及科學訓練的能力,讓我一生受用。在實驗過程中,錢老師給予我極大的空間思 考如何安排實驗及想要達成的研究方向,雖然過程跌跌撞撞,但老師總會在我需 要幫助時適時地提供意見;而在最後的論文撰寫、編修以及口試投影片的修改,
老師都能找出問題並給予修正的建議,在此非常的感謝錢老師這兩年的指導,能 夠加入老師的實驗室,真的非常的幸運。同時也感謝所上所有老師的教導,以及 我的口試委員 陳玉玲教授及 楊懿如副教授對於論文的提供寶貴的意見,讓這篇 論文更趨完整。
在實驗室生活中,要特別感謝偉豪學長及孟琳學姊,在忙碌的助教工作之 餘,還能在我實驗過程遇到困難的時候,又或找不到接下來方向時給予我很大的 幫助;接著要感謝為家學姊、品君學長、貞明學長,在我剛進實驗室,對於實驗 並不非常熟悉的情況下,教我許多實驗方法及技巧,並讓我實驗脈絡更加清楚,
平常也都會帶給實驗室許多的歡樂,讓 650 每天都充滿了笑聲,讓我能在這個環 境下心情愉快的進行實驗;也感謝思婗學妹幫忙分擔一些實驗室的雜事。另外還 要感謝我的同學們,和大家一起度過的解剖室生活是一個非常難忘的回憶,一起 吃飯、一起辦活動的回憶也都不會忘記的,謝謝欣惠、小涵和宛儒陪我度過每天 的午餐時刻,也時常給我鼓勵,恭喜大家順利畢業。
最後感謝我的家人,在我這兩年的碩士生生活中,給予我最大的支持與鼓
勵,讓我能無後顧之憂地繼續我的學業。我由衷地感謝曾經幫助過我的所有人以 及被犧牲的所有蝌蚪青蛙,因為有你們,今天我才能完成我的實驗、我的論文。
在此我想說聲,謝謝大家,我畢業了。
摘要
本研究我們選擇虎皮蛙 (學名: Hoplobatrachus rugulosus) 作為實驗動物;依 據實驗室先前的研究,我們發現雞的 α-internexin 及斑馬魚的 inaa 不僅會出現在 視網膜的感光細胞中,也可以在松果體中的類感光細胞中有表現分布。然而,α- internexin 在兩生類松果體中的表現尚未被探討,作為在演化史上可以彌補斑馬魚 及脊椎動物中空缺的角色,虎皮蛙可為一個重要的兩生類神經發育模式動物。
關於一種常用於兩生類發育研究的實驗動物:爪蟾 Xenopus laevis 的研究指出,
Xenopus α-internexin mRNA 序列已被命名為“xefiltin”。雖然虎皮蛙和 Xenopus 並
不是同一物種,但是屬於同一目。因此,本論文研究目的是在探討 α-internexin 的 同源基因 xefiltin: ina.S 及 nif.S 在虎皮蛙發育中,於視網膜及松果體內之表現與 分布狀況。因此,我們特別製作多株抗體可以標定虎皮蛙的 α-internexin 的同源基 因 ina.S 及 nif.S 蛋白,並使用西方墨點法與免疫組織染色確認 ina.S 及 nif.S
在發育中視網膜及松果體之表現與分布情況。為了確認 ina.S 及 nif.S 可以在松
果體中的類感光細胞中之可能表現分布,另外選擇了兩隻可用於標定感光細胞的 抗體: recoverin 及 XAP1 於實驗中來比較其分布情形。
根據免疫染色結果, nif.S 可以於感光細胞的外段 (outer segment) 被觀察到,
但是另一隻抗體: ina.S 只有在視網膜神經元 (retinal neurons) 中可檢測到。相對 的,並未在感光細胞中檢測到。在松果體中,recoverin 及 XAP1 皆可以標示出感 光細胞;另一方面,ina.S 及 nif.S 只能在早期虎皮蛙的松果體少許細胞中被偵測 到。綜合本研究結果,我們針對感光細胞在發育中虎皮蛙視網膜及松果體的表達和
分布做詳細的觀察及比較。我們認為XAP1,而非 ina.S 或 nif.S,可以應用成為研
究青蛙松果體中的感光細胞的特定標示物,有助於提供在青蛙的松果體中感光細 胞演化發育生物學研究更多線索。
關鍵字: 虎皮蛙(Hoplobatrachus rugulosus)、感光細胞、視網膜、松果體、α- internexin
Abstract
In this study, Hoplobatrachus rugulosus, commonly referred to as tiger-skinned frog,
is to be chosen the animal model. In previous studies, it has been found that both chicken α-internexin and zebrafish inaa (an ortholog of mammalian α-internexin) were
developmentally distributed not only in photoreceptors of retina but also in the photoreceptor-like cells of the pineal gland. However, from an evolutionary point of view, there is still lack of evidence to illustrate the temporal and spatial distribution of frog α- internexin protein in the developing pineal gland.
According to the previous studies about Xenopus laevis, the animal model that is usually used for amphibian developmental research, the mRNA sequence which encodes Xenopus α-internexin had been identified as “xefiltin”. Although H. rugulosus is not the
same species with X. laevis, both of them are classified in the same order. The aims of this study is to characterize the expression pattern of α-internexin-like proteins, internexin neuronal intermediate filament protein alpha S homeolog (ina.S) and low molecular weight neuronal intermediate filament S homeolog (nif.S) which might be expressed in the developing retina and the pineal gland of the tiger-skinned frog. Thus, for this study these 2 antibodies were generated to detect the frog α-internexin-like proteins and to confirm the distribution of these proteins in the developing frog retina as well as the pineal gland by western blot and immunohistochemistry. Furthermore, the markers applied to
detect the photoreceptor cell in the retina and the pineal gland are recoverin and XAP1
(Clone 3D2), both of them are reported to be specific to photoreceptors.
Based on the distribution patterns in frog retina, immunopositive staining of anti- nif.S could be observed in the outer segment of photoreceptors, but anti-ina.S could only be found in retinal neurons. In the pineal gland, both photoreceptor markers, recoverin and XAP1, could be detected. The α-internexin-like protein, ina.S and nif.S, could be detected only in a few photoreceptor-like cells at the early stage of the frog pineal gland.
In summary, this thesis has studied the distribution patterns of photoreceptor cells in retina and in pineal gland of developing tiger-skinned frogs. It can be concluded that XAP1 could be a good marker to identify the photoreceptor cells in the developing pineal gland of frogs.
Keywords: Tiger-skinned frog (Hoplobatrachus rugulosus), Photoreceptor, Retina, Pineal gland, α-internexin
Contents
誌謝 ..………...………...………. i
摘要 ..………...………...………... iii
Abstract ………….……….……….……… iv
List of Figures ……….………….……... vii
List of Tables ……….……….……..…..…... viii
Chapter 1: Introduction ………... 1
Chapter 2: Materials and Methods …………...………. 12
Chapter 3: Results ………..………...……… 18
Chapter 4: Discussion ….………...…… 24
Figures and Figure Legends ……….…. 29
Tables ……….……… 60
References ……….…. 64
List of Figures
Figure 1. Transverse section through the retina of salamander
(Hydromantes shastae) ……….. 29 Figure 2. Schematic drawing of photoreceptors in the retina of frog (Rana pipiens) ……….. 31 Figure 3. Stage of the tiger-skinned frog ………... 33 Figure 4. Phylogenetic analysis of Xenopus laevis nIFs with those from other species ……….. 35 Figure 5. View of male adult tiger-skinned frog brain ……….. 37 Figure 6. The protein levels of ina.S and nif.S in developing frog
tissues ……….………... 39 Figure 7. Distribution patterns of recoverin and XAP1 during the
development of frog retina ……… 41 Figure 8. Distribution patterns of ina.S and XAP1 during the development of frog retina ……….. 43 Figure 9. Distribution patterns of nif.S and XAP1 during the development of frog retina ……….. 45 Figure 10. Distribution patterns of recoverin and XAP1 during frog
developmental pineal gland ………... 47 Figure 11. Distribution patterns of ina.S and XAP1 during frog
developmental pineal gland ………... 51 Figure 12. Distribution patterns of nif.S and XAP1 during frog
developmental pineal gland ………... 55
List of Tables
Table 1. List of antibodies used for immunohistochemistry and western blot in the current study ………....……… 60 Table 2. Summary of protein expression in the developing tiger-skinned frog retina ……….…. 61 Table 3. Summary of protein expression in the developing tiger-skinned frog pineal gland ……….………... 62
Chapter 1: Introduction
The animal model: Xenopus laevis and Hoplobatrachus rugulosus
The African clawed frog Xenopus laevis is an amphibian species and frequently used in laboratory research for developmental biology and cell biology (Feehan et al., 2017). There are several proper properties for Xenopus to be used. The first reason is that standard protocols for Xenopus laboratory breeding and husbandry are available, which is comparably simpler and with lower cost than those required by rodents.
Secondly, it is easy to gain eggs and embryos because females lay large clutches of thousands of eggs and egg laying could be inducible by hormone priming (Browne et al., 2006).
On the other side, unlike X. laevisis who are native to sub-Saharan Africa,
Hoplobatrachus rugulosus’ natural habitats are in East Asian freshwater marshes, arable
land, pasture land, ponds, irrigated land, seasonally flooded agricultural land, and so forth. They are more familiar to us rather than Xenopus. This species is commonly referred to as being “field chicken” or “tiger-skinned frog” in Taiwan. Moreover, this thesis will refer to H. rugulosus as “tiger-skinned frog”. H. rugulosus is a kind of large, robust frog. In snout-vent length, males are about 6 to 8 cm, and females are 6 to 12 cm.
They can grow up to 15 cm. Observed from the appearance, the head length is slightly
larger than the head width, and the snout is sharp and long. Females are usually larger than males. This species of frogs’ breeding season is from spring to early summer. They are primarily carnivore. Their color is diversity, from yellow-green to dark gray. Also, they have dark speckles (Diesmos et al., 2004; Lin and Ji, 2005; Lue, 2012). When they are tadpoles, they are larger than other frogs. Their total length is about 5 cm and the tail length can be twice as long as the body length. Their back is green-brown with some small black spots, and on the eyes and mouth, some golden yellow spots can be observed. The tadpoles are demersal and likes to live in still waters, so they could usually be seen in ponds (Chou and Lin, 1997). In addition, their developmental processes are similar to X. laevisis.
Retina
Adult frogs are essentially dependent on vision. They feed on worms, flies, and other insects which they catch them by striking with the tongue or directly with the mouth. For this functional reflex, they use visual clues. Furthermore, frogs prey only on moving insects, and their attention is never attracted by stationary creatures or objects (Maturana et al., 1960). Moreover, when tiger-skinned frog are tadpoles, they could exercise all day, not limited by day-night cycle. Besides the eye vision, tadpoles
could also use the electrotactile stimulation of the skin to transduce visual data
(Blackiston and Levin, 2013). The function of the eyes is important for the daily life of adult frogs.
Looking into the eye, the amphibian retina shows the typical five-layered structure of vertebrates (Fig. 1). The outer nuclear layer (ONL), the inner nuclear layer (INL), and the ganglion cell layer (GCL) are separated by two fiber layers: the outer plexiform layer (OPL), which is thin, and the much thicker inner plexiform layer (INL). The two plexiform layers are the main site of synaptic contacts between five major types of retinal cells: photoreceptors, bipolar cells, ganglion cells, horizontal cells, and amacrine cells. (Heatwole et al., 1998; Purves et al., 2001). In the previous study, the ONL was formed by two layers of cell bodies in most frogs and salamanders. The rod cells nuclei are aligned at the distal side of the ONL, and the cone cells nuclei are more proximal (Gordon and Hood, 1976). In most other vertebrates, it is the opposite situation. Another special point is that the GCL consists of more than one row of ganglion cells in the amphibian (Heatwole et al., 1998).
Unlike primates or birds, there is no fovea in the retina of the amphibian. Only a few species even have a concentrated area sophisticated enough to be considered an area centralis, an analogue of the macula (Schwab, 2004). In some frogs, Hyla raniceps
(Bousfield and Pessoa, 1980), Heleioporus eyrie (Dunlop and Beazley, 1981), Bufo marinus (Nguyen and Straznicky, 1989), a streak of high cell density exists in the GCL.
The same situation expresses in the ONL (Zhang and Straznicky, 1991) and the INL (Zhu et al., 1990).
Photoreceptors
The eye is a specialized organ whose job is to convert or transduce light energy into neural impulses. A photoreceptor cell is a specialized type of neuroepithelial cell found in the retina that is capable of visual phototransduction. Photoreceptor cells of the vertebrate retina are highly specialized cells which respond to the stimulus of light, and transmit this response to adjoining neurons for ultimate relay to the visual centers of the brain (Young, 1967). Photoreceptor cells of amphibians, like those of other vertebrates, exhibit two main cells, rod cells and cone cells.
(i) Rod cells
In vertebrates, rod cells can functionalize in less intense light than the other types of visual photoreceptor. So, rods are responsible for dusk or even night vision.
Also, the rod pigments have a maximum absorption at 496 nm of visual spectrum and the image provided is one composed of gray tones, meaning that for seeing in black
and white (Pawlina et al., 2018). As for location, in the human eye, rods can be found everywhere in the retina, except in and near the fovea, and are used in peripheral vision. The number of rod cells is much higher than that of cone cells. There are approximately 90 million rod cells in the human retina (Curcio et al., 1990).
In the amphibians, rod cells are the largest photoreceptors and the most frequent receptors (Heatwole et al., 1998; Yovanovich et al., 2017). Rods are named after the cylindrical shape of their outer segments. The outer segment is the photosensitive region of the photoreceptor, and it is densely filled with free-floating disks formed by a double-layered membrane. As the reaction to light and darkness, photoreceptors can change their position. The myoid, the area between ellipsoid and nucleus, can be shortened or lengthened (Heatwole et al., 1998). The rod cells are narrower than the cones and distributed differently across the retina (Fig. 2).
Different from typical mammals which have a brightness-sensitive system and a three-color cone system, rods and cones of frog retina are able to both distinguish color (Denton and Wyllie, 1955; Heatwole et al., 1998; Yovanovich et al., 2017).
There are two types of rod cells, red rods and green rods. The most frequent
photoreceptors are red rods. The pigment of red rods in frogs is rhodopsin, which has its maximum absorbance in the green spectrum at about 502 nm. The green rods
contain the pigment with an absorbance at 432 nm, which is similar to the blue- sensitive cones in mammals, and have much shorter outer segments (Denton and Wyllie, 1955; Kelber et al., 2003; Yovanovich et al., 2017).
(ii) Cone cells
As opposed to rod cells, cones perform best in relatively bright light and are more sensitive to red, green, and blue regions of the visual spectrum in mammals.
Therefore, cones exist in three classes: L (588 nm), M (531 nm), and S (420 nm) (long-, middle-, and short-wavelength sensitive, respectively), but cannot be distinguished morphologically (Pawlina et al., 2006). In earlier studies, there were found to be about 4 to 7 million cones in a human eye and most are concentrated towards the macula (Osterberg et al., 1935; Curcio et al., 1990). Cone cells are densely packed in the fovea centralis, a nearly 0.3 mm diameter rod-free area with thin, densely packed cones which reduce in number towards the periphery of the retina. From shape, the outer segment of cone cells is much shorter and cone-shaped (Fig. 2).
Cone cells could be differentiated into two types, single and double in frog retina (Bowmaker, 1990; Heatwole et al., 1998; Pignatelli et al., 2010). The cone pigment is sensitive to yellow light at about 575 to 580 nm. The component of a double cone cell
carries a typical single cone and an accessory cone which pigment is the green- sensitive rhodopsin of the red rod (Heatwole et al., 1998). While the outer segments of the double cone cells are separated, the inner segments are fused.
The organization and function of the pineal gland
The pineal gland, also called the median third eye, develops as an evagination of the neuroepithelium of the dorsal diencephalon and serves as a photoreceptive
neuroendocrine organ (Sapede and Cau, 2013). The shapes and organizations of pineal glands depend on different species. In cold-blooded animals, pineal gland is located beneath the skull and is connected to the diencephalon by a stalk (Collin, 1971;
McNulty, 1984). The pineal of fish and frogs are a vesicle connected to the roof of the diencephalon by a slender stalk, and the lumen is opened to the third ventricle and is thus filled with cerebrospinal fluid (CSF) (Falcón et al., 1992; Omura and Oguri, 1969).
In lizards and avian, the pineal gland becomes follicular. In mammals, the pineal gland becomes glandular and more compact (Collin, 1971; Falcon, 1999; Vollrath, 1981).
In some of the lower vertebrates, pineal gland has the sensory function of light.
Since, this kind of pineal gland has the existence of photoreceptors, and is superficially situated in the brain, it is so-called the parietal eye (Kappers, 1979). Although the pineal
glands of postnatal mammalians no longer have photoreceptors, there are other nerve conduction pathways that indirectly affect the pineal gland for photosensory responses (Klein, 1985; Relkin, 1966). The mammals’ pineal gland is only endocrine. The function of the pineal gland is to act as the endocrine gland that secretes melatonin (N- acetyl-5-methoxytryptamine). Melatonin can regulate the circadian rhythm of the organism due to the difference in light duration, which includes changes in the daily cycle and seasonal changes (Falcón, 1999).
Intermediate filaments
Intermediate filaments (IFs) (~10 nm diameter) are expressed in cell-, tissue- and differentiation-specific function and play important roles in maintaining the mechanical stability through interconnection with other filamentous systems and provide
specialized functions via decorated by a variety of proteins (Oshima, 2007; Toivola et al., 2005). The IF proteins exhibit a common tripartite domain structure, with non- helical amino (head) and carboxyl-terminal (tail) domains flanking a central coil-coil α- helical core region (310-352 amino acids) (Kim and Coulombe, 2007; Sihag et al., 2007). IFs have been classified into six types based on the gene structure and amino acid sequence. IFs of neurons, or neuronal intermediate filaments (nIFs), are made of
five subunit proteins: the low, middle and high molecular mass neurofilament (NF) triplet proteins (designated as NF-L, NF-M and NF-H), α-internexin and peripherin (Thyagarajan et al., 2007). These proteins provide support and scaffold in outgrowth and stabilization of axon and dendrite (Lariviere and Julien, 2004). The developing nervous system undergoes progressive changes in the molecular composition of nIFs. α- Internexin, a 66 kDa nIF protein, is wildly expressed from early development through adulthood. Its expression in developing central nervous system (CNS) of mammals precedes the NF triplet proteins, arrives at its highest level before birth and declines postnatally (Fliegner et al., 1994; Kaplan et al., 1990). In adult mammals, chickens, and zebrafish, α-internexin has been found in retina, olfactory bulb, telencephalon, optic tectum and cerebellum (Fliegner et al., 1994; Chien and Liem, 1995; Chien et al., 1996;
Chien et al., 1998; Liu and Chien, 2013; Liu et al., 2013; Liao et al., 2016; Liao et al.,2018; Hao, 2018). These studies suggest that α-internexin might play a key role in neuronal cytoskeleton network during development.
The neuronal intermediate filaments (nIFs) in frog
According to the GenBank of NCBI (http://www.ncbi.nlm.nih.gov/genbank/), there are three α-internexin-like (ina.S, nif.L and nif.S), two NF-M, one NF-L and two
peripherin (plasticin) genes in the X. laevis. These nIF proteins could be classified into the same nIF categories, except nif.L (Charnas et al., 1992; Klein et al., 2002;
Strausberg et al., 2002) and nif.S (Klein et al., 2002; Strausberg et al., 2002). Some of the general features of expression of these nIF proteins have been documented in X.
laevis, including ina.S (internexin neuronal intermediate filament protein alpha S
homolog, known as xefiltin, an ortholog of mammalian α-internexin), nif.S (low molecular weight neuronal intermediate filament S homolog, also known as xefiltin), NF-M and NF-L (Sharpe et al., 1989; Gervasi and Szaro, 1997; Zhao and Szaro, 1997a;
b; Gervasi et al., 2000; Klein et al., 2002; Strausberg et al., 2002). However, to our best knowledge, there are very few studies to investigate what the role the nIFs play in the development of photoreceptors of frogs, so based on the phylogenetic analysis of Xenopus nIFs both ina.S and nif.S are candidates for studying the nIFs in the development of retina and pineal gland in tiger-skinned frogs.
Specific Aims
In previous studies of our laboratory, the distribution of α-internexin, inaa and inab, both the ortholog of mammalian α-internexin, could be discovered in the developing zebrafish pineal glands. In addition, inaa was found in cone photoreceptors of zebrafish
retina and also distinctively expressed in the photoreceptor-like cells of pineal gland, where inab was sparsely detected (Liao et al., 2016; Liao et al., 2018). Also, the expression of chicken α-internexin (chkINA) could be detected in the early stage of developing brains and found as the major IF protein in the parallel processes of
cerebellar granule neurons (Liu and Chien, 2013). Moreover, chkINA was expressed in all neuronal lineages of the developing chicken retina and pineal gland (Liu et al., 2013;
Hao, 2018). In the mouse model, α-internexin was reported to be detected postnatally in some pinealocytes and nerve processes (Ko et al., 2005). Moreover, some of
pinealocytes are shown to have the similar function to the photoreceptor of retina in many non-mammal vertebrates (Klein, 2004). However, from an evolutionary point of view, there is still lack of evidence to illustrate the temporal and spatial distribution of frog α-internexin protein in the developing pineal gland. The aims of this study are to characterize the expression patterns of α-internexin-like proteins, ina.S and nif.S, and to examine which are expressed in the pineal photoreceptor-like cells in the developing pineal gland of frog H. rugulosa. The photoreceptor markers, recoverin and XAP1, are also applied for the study of the developing retina and pineal gland in tiger-skinned frogs.
Chapter 2: Materials and Methods 1. Maintenance of tiger-skinned frog
Tadpoles and adults of H. rugulosa were purchased from the Tiger-Skinned Frog Plant, Frogs World, in Pingtung, Taiwan. Tadpoles were maintained at 25°C in a tank with shallow water which could cover all tadpoles and lived under a 12-h/12-h
light/dark cycle. The water in the tank was changed every three days (Ebinuma, 2013;
Mizutani, 2013). The tadpoles of tiger-skinned frogs are also carnivore, so the fresh fish fillets were prepared and been fed twice a day. Depend on postembryonic process driven by thyroid hormone signaling, there are four stages: premetamorphosis (growing fast), prometamorphosis (starting differentiation and hyperplasia), metamorphic climax (tail begins to retract) and completion of metamorphosis (Dodd and Dodd, 1976; Tata, 2006). The precise developing stages were done in reference to a table of Xenopus laevis, NF staging (Nieuwkoop and Faber, 1994). Tadpoles were collected at stage 46
(premetamorphosis, about 1 week after post-fertilization), stage 54 (prometamorphosis, with obvious hindlimbs, about 10 days from stage 46), stage 58 (prometamorphosis, with obvious forelimbs, approximately the other 10 days from stage 54), young adult (metamorphic climax, without a tail, approximately 1 month from stage 58) and adult (completion of metamorphosis, 4 months after metamorphosis) (Fig. 3). The growing time of tadpole is provided by the frog plant. The male adult frogs were collected for
this study. All animal experiments will be in accordance with the protocols approved by the Animal Care and Use Committee at the College of Medicine, National Taiwan University.
2. Generation of an epitope-specific polyclonal anti-ina.S and anti-nif.S antibodies
The sequence alignments and phylogenetic analysis were performed with Clustal Omega program (https://www.ebi.ac.uk/Tools/msa/clustalo/) (Fig. 4) and shaded by BoxShade 3.21 server (http://www.ch.embnet.org/software/BOX_form.html).
The specific polyclonal antibodies were produced by LTK BioLaboratories (Taoyuan, Taiwan). In brief, rabbit antiserum was prepared with synthetic oligopeptides, PSNPH PPPSYTYQSRVLS and KSKSGETIQEKTPQKSAA, corresponding to the C-terminal amino acid residues from frog ina.S and nif.S, respectively. The peptides were conjugated to complete adjuvant for antibody production in rabbits. The antiserum was further purified by peptide/protein specific affinity column. Fractions containing purified antibodies were collected and
concentrated into 1.5 mg/ml. Purified antibodies were collected and kept for further studies.
3. Protein extraction
For protein extraction, tissues were homogenized in protein RIPA buffer (iNtRON Biotechnology, Inc) 【50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1 mM DTT, 0.5%
NP-40, 1% Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate, 1 mM PMSF, 1 mM EDTA, 1 μM Pepstatin A, 1 μM Leupeptin, 0.1 μM Aprotinin】by
using tissue grinders. Homogenized tissues were incubated on ice for 15 mins, followed by sonication and centrifugation at 14000 rpm at 4°C for 15 mins. Finally, supernatants
were collected and the protein concentration was measured based on Bradford protein assay (Bio-Rad, Hercules, CA). The protein extract which was collected stored at -80°C until further studies.
4. SDS-PAGE and Western blot analysis
Protein lysates were denatured with 1Ⅹ sample buffer, boiled for 10 mins, and chilled on ice. Proteins (30 μg per lane) were then loaded onto 12% SDS-PAGE for
electrophoresis. Samples were transferred to 0.45 µm PVDF membranes (GE
Healthcare, Germany) via wet electrophoretic transfer and blocked with 5% nonfat dry
milk dissolved in 1Ⅹ TBST (0.1% Tween-20 in 1Ⅹ TBS) for 1 hr at room temperature.
The blot was incubated with appropriate antibodies (Tab. 1) diluted in 1Ⅹ TBST in 5%
nonfat dry milk at 4℃ overnight. Subsequently, the membranes were washed with 1Ⅹ
TBST and incubated with horseradish-peroxidase-conjugated secondary antibodies (Tab. 1) diluted in 1Ⅹ TBST in 5% nonfat dry milk at room temperature for 1 hr. The membranes were washed again and detected using enhanced chemiluminescence (Adansta, California, USA) and autoradiography.
5. Immunohistochemistry
First, tadpoles and frogs were anesthetized with 0.04% MS-222 (Sigma-Aldrich, St. Louis, USA) prior to sacrifice. Eyes of tadpoles, heads of tadpoles with their eyes removed, eyes of adults and brains of adults were removed and immersed in 4%
paraformaldehyde (PFA) in 0.1M phosphate buffer (PB), pH 7.4, for 12-16 hrs at 4℃, depending on the size of the samples. Samples were cryoprotected with 30% sucrose for 3 days. Afterward, selected specimens were embedded in optimal cutting temperature compound (OCT, Thermo-Shandon Limited, Cheshire, UK), and frozen in isobutene with dry ice and stored in -80℃, as described by Redies and Takeichi (1993).
We sliced from the bottom of frog brains through the horizontal plane (Fig. 5).
Cryostat sections of the tissue were collected and placed on microscope slides (Superfrost, Thermo Scientific, Waltham, MA) at a thickness of 20 µm by a cryostat (Leica CM3050) for immunohistochemistry. Sections were fixed with 100% ice-cold methanol for 15 mins and then blocked with 3% fetal bovine serum (FBS) in PBS for 1
hr. Subsequently, sections were incubated with primary antibodies in blocking solution (3% fetal bovine serum in PBS) overnight at 4℃, then rinsed 3 times 5 mins in PBS,
and incubated with secondary antibodies and Hoechst 33342 (1:1000, stock concentration 10 μg/ml; Catalog # H1399 Invitrogen) diluted in PBS at room temperature for 1 hr. Finally, sections were washed in PBST (0.1% Triton X100 in PBS) for 3 times 5 mins each and in PBS for 5 mins, and mounted with Fluoro-Gel (Cat
# 17985-10, Electron Microscopy Sciences, Hatfield, PA). All images were acquired with a Leica TCS SP5 confocal microscope (Leica) and Carl Zeiss LSM880 confocal microscope (Zeiss). The following primary antibodies were used in this study: rabbit anti-ina.S, rabbit anti-nif.S, anti-XAP1 (clone 3D2, DSHB), and anti-recoverin (AB5585, Chemicon International). The secondary antibodies Alexa 488 donkey anti- rabbit and Alexa 594 donkey anti-mouse (1:200, Life Technologies) were applied in this study (Tab. 1).
6. Statistical analysis
The cells which were positive for both XAP1 and Hoechst by
immunohistochemistry were count in order to comprehend the distribution patterns of XAP1 in the developmental pineal gland. The statistical data was analysed by
software Image J 1.43 (NIH). For comparison of multiple conditions, one-way
AVONA was performed and plotted by GrapdPadPrism® 7.0. All data was presented as mean values ± SD. A p-value less than 0.05 was considered statistically
significant. N=5 for each stage.
Chapter 3: Results
Expression levels of ina.S and nif.S proteins in developing tiger- skinned frog tissues
To certify generated polyclonal anti-ina.S and anti-nif.S antibodies could be expressed in developing tiger-skinned frog tissues, western blotting were performed for protein identification. Brain, retina and liver were taken and extracted. The brain tissues were specified in diencephalon including pineal gland. St.46, st.54, st.58 and adult were selected to perform the stages from developing to mature (Fig. 6). Immunoblotting used anti-ina.S antibody predicted 55-60 kDa and anti-nif.S predicted 53-54 kDa. The result revealed that both ina.S and nif.S could be observed in frog brain and retina.
Immunohistochemical patterns of photoreceptor marker, recoverin and XAP1 in developing tiger-skinned frog retina
Previous study has shown that the antibody against recoverin labels only cone cells in mouse retina (Biswas et al., 2014). Also, XAP1 reactivity was primarily expressed on the surfaces of the outer segments of both rods and cones in Xenopus (Harris et al., 1992). In the study, these two antibodies, recoverin and XAP1, were used in immunohistochemistry staining to recognize tiger-skinned frog retina. The
immunofluorescence staining of recoverin and XAP1 were performed at retina of tadpoles at st.46, st.54, st.58, young adult and adult. The immunoreactivity of recoverin could be detected in the cell body of photoreceptors in outer nuclear layer in retina (Fig.
7, A-E) in every stages of frogs that were collected. Same as the patterns of recoverin in developing retina, the immunoreactivity of XAP1 could be observed in the outer
segment of photoreceptor (Fig. 7, A’-E’). The result demonstrated that the photoreceptor cells in tiger-skinned frog retina could be identified by both anti-recoverin and anti- XAP1 antibodies.
Immunohistochemical patterns of ina.S in developing tiger-skinned frog retina
In previous studies, both zebrafish inaa (Liao et al., 2016) and chicken α-internexin (chkINA) (Liu et al., 2013; Hao, 2018) could be identified in the photoreceptor-like cells in both retina and pineal gland. To test whether the antibody of ina.S is able to identify photoreceptor-like cells, we used epitope-specific polyclonal anti-ina.S antibody to exam the patterns of tiger-skinned frog retina via immunohistochemistry.
The retina of tadpoles at st.46, st.54, st.58, young adult and adult were performed by the immunofluorescence staining. The immunoreactivity of ina.S could be found only in the
retinal neurons of ganglion cell layer and inner nuclear layer, but not in photoreceptors (Fig. 8, A-E). The distribution pattern of ina.S is similar to that of zebrafish inab (Liao et al., 2016).
Immunohistochemical patterns of nif.S in developing tiger-skinned frog retina
Since anti-ina.S failed to be identified in photoreceptors of frog retina, another α- internexin-like protein, frog nif.S, were tried to examine the immunoreaction in frog retina by way of immunohistochemistry. The stages of immunofluorescence staining were performed at tadpoles at st.46, st.54, st.58, young adult and adult. The
immunopositive staining of nif.S in the retina could be detected not only in ganglion cell layer and inner nuclear layer, but also in the outer segment of photoreceptors (Fig.
9, A-E). From a single optic section of the confocal image (Fig. 9, F”), nif.S could be colocalized with XAP1 in the outer segment of photoreceptors of adult retina. Based on this observation, this study suggests that the distribution pattern of nif.S is similar to that of zebrafish inaa (Liao et al., 2016) and chkINA (Liu et al., 2013; Hao, 2018).
Immunohistochemical patterns of photoreceptor marker, recoverin and XAP1 in developing tiger-skinned frog pineal gland
Pineal tissues from five stages of frog brains were collected and sectioned:
tadpoles at st.46, st.54, st.58, young adult and adult. The immunostaining of anti- recoverin could be detected within pineal tissues at st.46 (Fig.10a, A), and decreased at the following developmental stages (Fig. 10a, B-E). The expression of XAP1 could be detected in the photoreceptor-like cells of pineal gland at all stages (Fig. 10a, B’-E’).
The immunopositive staining of recoverin could be found in some fields of adult pineal gland (Fig. 10b, G), but failed to be colocalized with XAP1 (Fig. 10b, J, white arrow).
Furthermore, according to statistical analysis, the distribution patterns of XAP1 in the developmental pineal gland did not show any significant changes (P>0.05) (Fig. 10b;
11b; 12b, F). From the results, it is suggested that some photoreceptor cells might exist in frog pineal gland during development.
Immunohistochemical patterns of ina.S in developing tiger-skinned frog pineal gland
From the previous studies, the distributions of inaa and inab, both the ortholog of mammalian α-internexin, could be discovered in developing zebrafish pineal gland
(Liao et al., 2016; Liao et al.,2018), and chkINA could be identified in the developing chicken retina as well as pineal gland (Liu et al., 2013; Hao, 2018). Therefore, we further examined the distribution patterns of internexin neuronal intermediate filament protein alpha S homeolog, ina.S in frog pineal gland via immunohistochemistry. The immunofluorescence staining of ina.S was performed at pineal gland of tadpoles at st.46, st.54, st.58, young adult and adult. The immunopositive staining of anti-ina.S could be rarely detected in the development of pineal gland (Fig. 11a, A’-E’). The immunopositive staining of ina.S could be found in some observation fields of adult pineal glands (Fig. 11b, G). However, ina.S could not be colocalized with XAP1 within pineal tissues from this study’s observations (Fig. 11 b, J, white arrows).
Immunohistochemical patterns of nif.S in developing tiger-skinned frog pineal gland
To examine the expression pattern of nif.S in the developing pineal gland of frogs, this study performed immunostaining of anti-nif.S antibody at pineal gland of tadpoles at st.46, st.54, st.58, young adult and adult. The expression of nif.S could be found in some cells of the early developing pineal gland (Fig. 12a, A-B), but decreased in the following stages (Fig. 12a, C-E). From the observation, the immunopositive staining of
nif.S could be only found in some areas in the pineal tissues from adults (Fig. 12b, G).
Yet, it failed to be colocalized with XAP1 (Fig. 12b, J, white arrow). Based on this study observations, it can be suggested that anti-nif.S might not be as good as a photoreceptor marker for identifying photoreceptor-like cells in pineal gland of tiger- skinned frogs.
Chapter 4: Discussion
Distribution patterns of ina.S and nif.S in developing tiger-skinned frogs
(i) Retina
α-internexin has been studied and found in retina of mouse, chicken, and zebrafish (Chien and Liem, 1995; Liu and Chien, 2013; Liu et al., 2013; Liao et al., 2016; Liao et al., 2018; Hao, 2018). Taking zebrafish for example, it expresses inaa and inab as homologs of the mammalian α-internexin. Inaa was reported to be strongly expressed in the cone photoreceptor of adult zebrafish retina, and also in interneurons, such as bipolar cells and amacrine cells (Liao et al., 2016); on the other hand, inab was predominantly expressed in ganglion cell layer, optic fiber layer and optic nerve (Liao et al., 2018). In this study of frog retina, 2 antibodies, ina.S and nif.S, were applied and presented similar to patterns. In retina, the immnopositive staining of anti-ina.S could be observed in ganglion cell layer and retinal neurons, but not in photoreceptors (Fig. 7, E; Tab. 2). On the other side, the immunoreactivity of nif.S could be found not only in ganglion cell layer and inner nuclear layer, but also in the outer segment of photoreceptors (Fig. 8, A-E; Tab. 2). Thus, this study suggests that the expression of frog nif.S is more like that of zebrafish inaa in retina.
(ii) Pineal gland
It has been reported that the neuronal intermediate filaments α-internexin and peripherin could be found in mouse pineal gland, and some cells may differentiate into neuron-like cells in postnatal mouse pineal gland. In addition, these cells possess dual properties of CNS and PNS neurons in nature (Ko et al., 2005). Researchers also found that postnatal neurodevelopment and differentiation may lead to a different expression of neuron-specifiic markers in different species (Badiu et al., 2001). In previous studies, the distribution of inaa and inab could be identified in developing zebrafish pineal gland. In addition, only inaa was distinctively expressed in the photoreceptor-like cells of the pineal gland, where inab was sparsely detected (Liao et al., 2016; Liao et al., 2018). Besides, the expression of chicken α-internexin
(chkINA) could be detected at the early stage of developing brains and found as the major IF protein in the parallel processes of cerebellar granule neurons (Liu and Chien, 2013). Moreover, chkINA was expressed in all neuronal lineages of the developing chicken pineal gland (Liu et al., 2013; Hao, 2018). In this study, 2 α- internexin-like proteins, ina.S and nif.S, were used to study the nIFs in the
development of frog pineal glands. The immunoreactivity of ina.S was hard to be detected in pineal gland (Fig. 11a, A-E). Yet, the expression of nif.S could be found in
a few cells of pineal gland (Fig. 12a, A-E). From the observation, α-internexin-like proteins (ina.S and nif.S) could be found only in the early developmental stages of pineal gland (Tab. 3). The existence of neuronal cells in the development of frog pineal gland is needed to be further clarified.
(iii) Patterns comparison
According to the GenBank of NCBI and sequence comparison by Clustal Omega, this study studied the nIFs, ina.S and nif.S , in the development of retina and pineal gland in tiger-skinned frogs. However, the results were not like those in other species. Only nif.S could be identified in photoreceptors of retina, and both ina.S and nif.S were hard to be detected in adult pineal gland. From this study’s data, it can be suggested that both α-internexin-like proteins might be only expressed transiently in the neuronal procursors during early development of frog pineal gland.
The photoreceptor cells in developing pineal gland of tiger-skinned frogs
Some studies showed that the antibody against recoverin labels only cone cells in mouse and macaque monkey retina (Biswas et al., 2014; Puthussery et al., 2014).
Another study reported that XAP1 reactivity was primarily expressed on the surfaces of
the outer segments of both rods and cones in Xenopus retina (Harris et al., 1992). From this study’s observation of tiger-skinned frogs, the expression of recoverin could be found to decrease following the developmental pineal gland, while the expression pattern of XAP1 is not significantly changed in frogs (Tab. 3). Based on this result, it can be hypothesized that the cell type of photoreceptors might be mostly rod cells in adult frog pineal gland.
Another study indicates in another species, trout, has the similar finding of photoreceptor cells in their pineal glands. The study indicates that the photoreceptors are mostly classified into two groups, which have the peak sensitivities of 495 and 521 nm by intracellular recordings (Marchiafava and Kusmic, 1993). Other research
indicates that fish and amphibians usually have their light absorption maxima clustered around 500 nm, containing most of the standard rod pigments (Liebman, 1972; Mariani, 1986; Lythgoe and Partridge, 1989; Perry and McNaughton, 1991; Hárosi, 1994; Xu et al., 1997). From the results, it can be suggested that some photoreceptor cells might exist in frog pineal glands during development. Also, the expression pattern of recoverin could be found decreased, while the expression of XAP1 is not appreciably changed (Tab. 3). From the previous studies in both antibody research of recoverin and XAP1, and the study of trout, these lead to a stronger speculation that the majority of cell
clusters in adult pineal gland of frogs are rod cells. These rod-like cells might play a role in the function of photo-sensation of the third eye in the amphibian tiger-skinned frog.
Figure and Figure Legends
Figure 1. Transverse section through the retina of salamander (Hydromantes shastae)
H. shastae is a species of salamander, a group of amphibians typically characterized by
a lizard-like appearance. The amphibian retina shows the typical five-layered structure of vertebrates. The outer nuclear layer, the inner nuclear layer, and the ganglion cell layer are separated by two fiber layers: the outer plexiform layer and the inner plexiform layer. PR, photoreceptor outer segments; ONL, outer nuclear layer; OPL, outer
plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer. Scale bar = 20μm. (Figure modified from Heatwole et al., 1998)
Figure 1.
Figure 2. Schematic drawing of photoreceptors in the retina of frog (Rana pipiens)
Photoreceptor cells of amphibians exhibit two main cells, rod cells and cone cells. There are two types of rod cells, red rods (absorbance in green spectrum at 502 nm) and green rods (sensitive to blue light at 432 nm). The most frequent photoreceptors are red rods.
Cone cells could also be differentiated into two types, single and double. The
component of a double cone cell carries a typical single cone (sensitive to yellow light at about 575 to 580 nm) and an accessory cone which pigment is green-sensitive rhodopsin. While the outer segments of the double cone cells are separated, the inner segments are fused. Compared to rod cells and cone cells: rods are much thinner and taller than cones. OS, outer segment. (Figure modified from Donner and Reuter, 1976)
Figure 2.
OS
Figure 3. Stage of the tiger-skinned frog
Tadpoles were collected at stage 46 (about 1 week after post-fertilization), stage 54 (with obvious hindlimbs, about 10 days from stage 46), stage 58 (with obvious
forelimbs, approximately the other 10 days from stage 54), young adult (without a tail, approximately 1 month from stage 58) and adult (4 months after metamorphosis). The male adult frogs were collected for this study. The developing stages are with reference to the NF staging table for Xenopus laevis (Nieuwkoop and Faber, 1994).
Figure 3.
Figure 4. Phylogenetic analysis of Xenopus laevis nIFs with those from other species.
The cladogram of nIFs from different species was constructed by Clustal Omega.
Comparison of sequence identities between nIFs from different species, including neurofilament triplets (NFL, NFM and NFH), α-internexin and peripherin, revealed the evolutionary relationship and classification of these proteins. Because the tiger-skinned frog developmental processes are similar to Xenopus and the sequence of tiger-skinned frogs are not yet confirmed, this study tested Xenopus nIFs on tiger-skinned frogs.
Homo sapiens is human. Mus musculus is mouse. Gallus gallus is chicken. Danio rerio is zebrafish.
Figure 4.
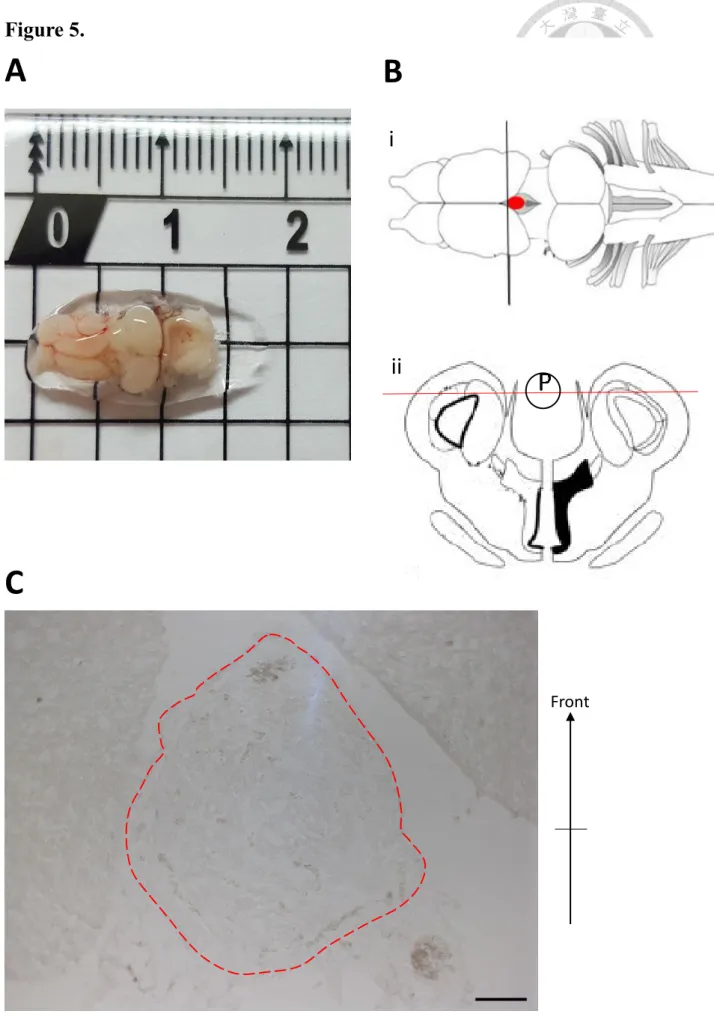
Figure 5. View of male adult tiger-skinned frog brain
A. The brain collected from male adult tiger-skinned frog. The unit in the picture is
centimeter (cm).
B. The illustration of top view of adult frog brain. Red dot is the location of the pineal
gland. Cut the brain along with black line, then we could get the coronal view of frog brain (i). Red line is the way we sliced frog brain (ii). (Figure modified from Cui et al., 2014)
C. The bright field of adult tiger-skinned frog brain. The area surrounded by red dashed
lines is pineal gland, and the area next to pineal gland are parts of forebrain.
Scale bars = 100 μm.
Figure 5.
A
C
B
i
ii
Front
Figure 6. The protein levels of ina.S and nif.S in developing frog tissues
Protein extracts were obtained from different stages (st.46, st.54, st.58 and adult) of frog brains containing pineal gland and other adult frog tissues, retina and liver. Proteins were subjected to 12 % SDS-PAGE followed by Western blotting. Specific antibodies against ina.S (55-60 kDa) and nif.S (53-54 kDa) were applied. Antibodies against β- actin was used as loading controls. The result revealed that both ina.S and nif.S could be observed in frog brain and retina. Also, both anti-ina.S and nif.S antibodies are
polyclonal antibodies, so there was a nonspecific band at the lane of nif.S of adult liver.
Figure 6.
Figure 7. Distribution patterns of recoverin and XAP1 during the development of frog retina
The tiger-skinned frog eyes were collected and sectioned from five developmental stages (tadpoles at st.46, st.54, st.58, young adult and adult). Immunohistochemical staining were applied by 2 photoreceptor markers, polyclonal anti-recoverin (green, A- E) and monoclonal anti-XAP1 (red, A’-E’), respectively. All sections were
counterstained with Hoechst 33342 (blue, A’’-E’’) to identify nuclei of retina. The immunoreactivity of recoverin could be detected in the cell body of photoreceptors in outer nuclear layer (ONL) for all stages we collected, while the immunoreactivity of XAP1 was observed in the outer segment of photoreceptor. Scale bars = 25 μm.
Figure 7.
Figure 8. Distribution patterns of ina.S and XAP1 during the development of frog retina
Eyes from frogs were collected and sectioned from five developmental stages (tadpoles at st.46, st.54, st.58, young adult and adult). Immunohistochemical staining were sectioned and labeled with polyclonal anti-ina.S antibody (green, A-E) and monoclonal anti-XAP1 (red, A’-E’). All sections were counterstained with Hoechst 33342 (blue, A’’-E’’) to identify nuclei. The immnopositive staining of anti-ina.S could be found in ganglion cell layer (GCL) and retinal neurons, but not in photoreceptors. It also could be observed that ina.S increased in inner nuclear layer (INL), while the
immunoreactivity of XAP1 was observed in outer segment of photoreceptor.
Scale bars = 25 μm.
Figure 8.
Figure 9. Distribution patterns of nif.S and XAP1 during the development of frog retina
Eyes from frogs were collected and sectioned from five developmental stages (tadpoles at st.46, st.54, st.58, young adults and adults). Immunohistochemical staining were sectioned and labeled with polyclonal anti-nif.S antibody (green, A-E) and monoclonal anti-XAP1 (red, A’-E’). All sections were counterstained with Hoechst 33342 (blue, A’’-E’’) to identify nuclei of retina. The immunopositive staining of nif.S in the retina could be detected not only in GCL and INL, but also in the outer segment of
photoreceptors, while the immunoreactivity of XAP1 was observed in the outer segment of photoreceptor. This study observed that nif. S and XAP1 were partially colocalized in the outer segments of photoreceptors in the developmental retina (A”-E”). From single optic section, it can be showed the colocalization between nif. S and XAP1 in the outer segment of photoreceptor at adult retina (F”). Scale bars = 25 μm.
Figure 9.
nif.SSingle Optic SectionXAP1 FF’F”
ONL INL GCL
Figure 10a. Distribution patterns of recoverin and XAP1 during frog developmental pineal gland
Pineal tissues from tiger-skinned frog brains were collected and sectioned at five stages (tadpoles at st.46, 54, 58, young adults and male adults). The immunostaining of anti- recoverin (green) could be detected at st.46 (A), and decreased at the following developmental stages (B-E). The expression of XAP1 (red) could be detected in the photoreceptor-like cells of pineal gland at all stages (A’-E’). All sections were
counterstained with Hoechst 33342 (blue, A’’-E’’) to identify the nuclei of pineal gland.
The Pattern diagram showed the area of pineal gland surrounding by red line, and the area surrounding by black dashed line might be forebrain or pia mater (A’’’-E’’’).
Scale bars = 25 μm.
Figure 10a.
Figure 10b. Distribution patterns of recoverin and XAP1 during frog developmental pineal gland
According to the statistical analysis, the distribution patterns of XAP1 did not show any significant changes (P>0.05) (F). The error bars represent the standard deviation. n=5 for each stage. In addition,adult frog pineal glands were collected, sectioned and stained with polyclonal anti-recoverin antibody (green, G), monoclonal anti-XAP1 (red, H), and also counterstained with Hoechst 33342 (blue, I). In the merge image J, it shows that the immunopositive staining of recoverin could be found around a single cell in the center (white arrow). This study observed this pattern in some fields of adult pineal gland.
Scale bars = 25 μm.
Figure 10b.
Figure 11a. Distribution patterns of ina.S and XAP1 during frog developmental pineal gland
Pineal tissues from tiger-skinned frogs were collected and sectioned at five stages (tadpoles at st.46, 54, 58, young adults and adults). Immunohistochemical staining were sectioned and labeled with polyclonal anti-ina.S antibody (green, A-E) and monoclonal
anti-XAP1 (red, A’-E’). All sections were counterstained with Hoechst 33342 (blue, A’’-E’’) to identify the nuclei. The Pattern diagram showed the area surrounding by red
line were pineal gland, and the area surrounding by black dashed line might be forebrain or pia mater (A’’’-E’’’). The immunopositive staining of anti-ina.S was rarely detected
in the development of pineal gland, while the immunostaining of anti-XAP1 could be detected in the photoreceptor-like cells of pineal gland at all stages. Scale bars = 25 μm.
Figure 11a.
Figure 11b. Distribution patterns of ina.S and XAP1 during frog developmental pineal gland
According to the statistical analysis, the distribution patterns of XAP1 did not show any significant changes (P>0.05) (F). The error bars represent the standard deviation. n=5 for each stage. Moreover, adult frog pineal glands were collected, sectioned and stained with polyclonal anti-ina.S antibody (green, G), monoclonal anti-XAP1 (red, H), and also counterstained with Hoechst 33342 (blue, I). In the merge image J, it shows that the immunopositive staining of ina.S could be found around some cells (white arrows). The study observed the pattern in some fields of adult pineal gland. Scale bars = 25 μm.
Figure 11b.
Figure 12a. Distribution patterns of nif.S and XAP1 during frog developmental pineal gland
Pineal tissues from tiger-skinned frogs were collected and sectioned at five stages (tadpoles at st.46, 54, 58, young adult and adult). Immunohistochemical staining were sectioned and labeled with polyclonal anti-nif.S antibody (green, A-E) and monoclonal anti-XAP1 (red, A’-E’). All sections were counterstained with Hoechst 33342 (blue, A’’-E’’) to identify the nuclei. The Pattern diagram showed the area of pineal gland
surrounding by red line, and the area surrounding by black dashed line might be
forebrain or pia mater (A’’’-E’’’). The expression of nif.S could be found in some cells
during early development of pineal gland, while the immunostaining of anti-XAP1 could be detected in the photoreceptor-like cells of pineal gland at all stages.
Scale bars = 25 μm.
Figure 12a.
Figure 12b. Distribution patterns of nif.S and XAP1 during frog developmental pineal gland
According to the statistical analysis, the distribution patterns of XAP1 did not show any significant changes (P>0.05) (F). The error bars represent the standard deviation. n=5 for each stage. Furthermore, adult frog pineal glands were collected, sectioned and stained with polyclonal anti-nif.S antibody (green, G), monoclonal anti-XAP1 (red, H), and also counterstained with Hoechst 33342 (blue, I). In the merge image J, it shows that the immunopositive staining of nif.S could be found (white arrow), but did not colocalize with XAP1. The study observed this kind of pattern in some fields of adult pineal gland. Scale bars = 25 μm.
Figure 12b.
Tables
Table 1. List of antibodies used for immunohistochemistry and western blot in the current study
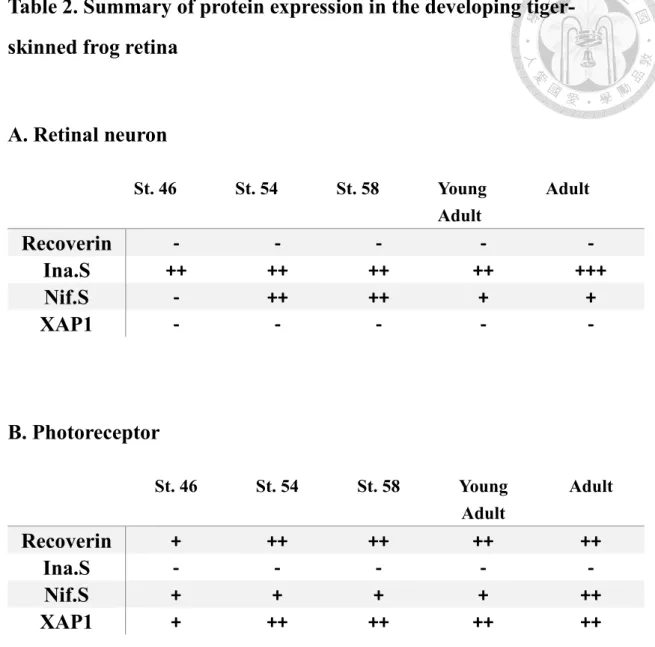
Table 2. Summary of protein expression in the developing tiger- skinned frog retina
A. Retinal neuron
St. 46 St. 54 St. 58 Young
Adult
Adult
Recoverin - - - - -
Ina.S ++ ++ ++ ++ +++
Nif.S - ++ ++ + +
XAP1 - - - - -
B. Photoreceptor
St. 46 St. 54 St. 58 Young
Adult
Adult
Recoverin + ++ ++ ++ ++
Ina.S - - - - -
Nif.S + + + + ++
XAP1 + ++ ++ ++ ++
Abbreviations: ina.S, internexin neuronal intermediate filament protein alpha S homeolog; nif.S, low molecular weight neuronal intermediate filament S homeolog.
Expression levels of staining were judged as follow: -, no immunoreactivity; +, weak immunoreactivity; ++, moderate immunoreactivity. +++, strong immunoreactivity.
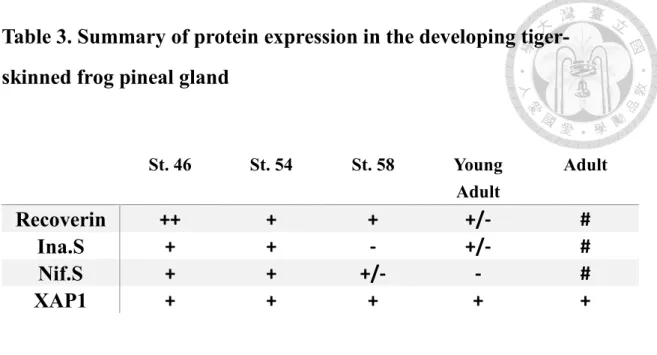
Table 3. Summary of protein expression in the developing tiger- skinned frog pineal gland
St. 46 St. 54 St. 58 Young
Adult
Adult
Recoverin ++ + + +/- #
Ina.S + + - +/- #
Nif.S + + +/- - #
XAP1 + + + + +
Abbreviations: ina.S, internexin neuronal intermediate filament protein alpha S homeolog; nif.S, low molecular weight neuronal intermediate filament S homeolog.
Expression levels of staining were judged as follow: -, no immunoreactivity; #, could be detected in some fields; +, weak immunoreactivity; ++, moderate immunoreactivity.
+++, strong immunoreactivity.
References
Badiu, C., Badiu, L., Coculescu, M., Vilhardt, H., & Møller, M. (2001). Presence of oxytocinergic neuronal‐like cells in the bovine pineal gland: an
immunocytochemical and in situ hybridization study. Journal of pineal research, 31(3), 273-280.
Biswas, S., Haselier, C., Mataruga, A., Thumann, G., Walter, P., & Muller, F. (2014).
Pharmacological analysis of intrinsic neuronal oscillations in rd10 retina. PLoS One, 9(6), e99075.
Blackiston, D. J., & Levin, M. (2013). Ectopic eyes outside the head in Xenopus tadpoles provide sensory data for light-mediated learning. The Journal of Experimental Biology, 216(6), 1031-1040.
Bousfield, J., & Pessoa, V. (1980). Changes in ganglion cell density during post- metamorphic development in a neotropical tree frog Hyla raniceps. Vision research, 20(6), 501-510.
Bowmaker, J. K. (1990). Visual pigments of fishes. In The visual system of fish (pp. 81- 107): Springer.
Browne, R. K., Seratt, J., Vance, C., & Kouba, A. (2006). Hormonal priming, induction of ovulation and in-vitro fertilization of the endangered Wyoming toad (Bufo baxteri). Reproductive Biology and Endocrinology, 4(1), 34.
Charnas, L. R., Szaro, B. G., & Gainer, H. (1992). Identification and developmental expression of a novel low molecular weight neuronal intermediate filament protein expressed in Xenopus laevis. J Neurosci, 12(8), 3010-3024.
Chien, C. L., & Liem, R. K. (1995). The neuronal intermediate filament, α-internexin is
transiently expressed in amacrine cells in the developing mouse retina.
Experimental eye research, 61(6), 749-756.
Chien, C. L., Lee, T. H., & Lu, K. S. (1998). Distribution of neuronal intermediate filament proteins in the developing mouse olfactory system. J Neurosci Res, 54(3), 353-363.
Chien, C. L., Mason, C., & Liem, R. (1996). α‐internexin is the only neuronal intermediate filament expressed in developing cerebellar granule neurons.
Journal of neurobiology, 29(3), 304-318.
Chou, W. H., & Lin, J. Y. (1997). Tadpoles of Taiwan. National Museum of Natural Sciences in Taiwan, Special Publication, 7, 1-98.
Collin, J. P. (1971). Differentiation and regression of the cells of the sensory line in the epiphysis cerebri. The pineal gland, 79-125.
Curcio, C. A., Sloan, K. R., Kalina, R. E., & Hendrickson, A. E. (1990). Human photoreceptor topography. J Comp Neurol, 292(4), 497-523.
doi:10.1002/cne.902920402
Denton, E. J., & Wyllie, J. (1955). Study of the photosensitive pigments in the pink and green rods of the frog. The Journal of Physiology, 127(1), 81-89.
Diesmos, A. C., Brown, R. M., & Gee, G. V. (2004). Preliminary report on the
amphibians and reptiles of Balbalasang-Balbalan national Park, Luzon Island, Philippines. Sylvatrop, The Technical Journal of Philippine Ecosystems and Natural Resources, 13, 63-80.
Dodd, M. H. I., & Dodd, J. M. (1976). The biology of metamorphosis. Physiology of the Amphibia, 3, 467-599.
Donner, K., & Reuter, T. (1976). Visual pigments and photoreceptor function. In Frog neurobiology (pp. 251-277): Springer.
Dunlop, S., & Beazley, L. (1981). Changing retinal ganglion cell distribution in the frog Heleioporus eyrei. Journal of Comparative Neurology, 202(2), 221-236.
Eisthen, H. (2000). Amphibian Biology. Volume 3. Sensory Perception (Vol. 2000).
Falcón, J. (1999). Cellular circadian clocks in the pineal. Progress in neurobiology, 58(2), 121-162.
Falcón, J., Thibault, C., Begay, V., Zachmann, A., & Collin, J.-P. (1992). Regulation of the rhythmic melatonin secretion by fish pineal photoreceptor cells. In Rhythms in fishes (pp. 167-198): Springer.
Feehan, J. M., Chiu, C. N., Stanar, P., Tam, B. M., Ahmed, S. N., & Moritz, O. L.
(2017). Modeling dominant and recessive forms of retinitis pigmentosa by editing three rhodopsin-encoding genes in Xenopus laevis using Crispr/Cas9.
Scientific reports, 7(1), 6920.
Fliegner, K. H., Kaplan, M. P., Wood, T. L., Pintar, J. E., & Liem, R. K. (1994).
Expression of the gene for the neuronal intermediate filament protein α‐
internexin coincides with the onset of neuronal differentiation in the developing rat nervous system. Journal of Comparative Neurology, 342(2), 161-173.
Gervasi, C., Stewart, C. B., & Szaro, B. G. (2000). Xenopus laevis peripherin (XIF3) is expressed in radial glia and proliferating neural epithelial cells as well as in neurons. J Comp Neurol, 423(3), 512-531.
Gervasi, C., & Szaro, B. G. (1997). Sequence and expression patterns of two forms of the middle molecular weight neurofilament protein (NF-M) of Xenopus laevis.
Molecular brain research, 48(2), 229-242.
Gordon, J., & Hood, D. C. (1976). Anatomy and physiology of the frog retina. In The Amphibian Visual System (pp. 29-86): Elsevier.
Hao, C. M. (2018). The development of photoreceptor-like cells in chicken pineal gland.
(master). National Taiwan University, Available from Airiti AiritiLibrary database. (2018)
Harris, W. A., & Messersmith, S. L. (1992). Two cellular inductions involved in photoreceptor determination in the Xenopus retina. Neuron, 9(2), 357-372.
Heatwole, H. (1998). Amphibian Biology: Sensory perception: Surrey Beatty & Sons.
Hárosi, F. I. (1994). An analysis of two spectral properties of vertebrate visual pigments.
Vision research, 34(11), 1359-1367.
Kaplan, M. P., Chin, S., Fliegner, K. H., & Liem, R. (1990). Alpha-internexin, a novel neuronal intermediate filament protein, precedes the low molecular weight neurofilament protein (NF-L) in the developing rat brain. Journal of Neuroscience, 10(8), 2735-2748.
Kappers, J. A. (1979). Short history of pineal discovery and research. In Progress in brain research (Vol. 52, pp. 3-22): Elsevier.
Kelber, A., Balkenius, A., & Warrant, E. J. (2003). Colour vision in diurnal and nocturnal hawkmoths. Integrative and Comparative Biology, 43(4), 571-579.
Kelber, A., Vorobyev, M., & Osorio, D. (2003). Animal colour vision–behavioural tests and physiological concepts. Biological Reviews, 78(1), 81-118.
Kim, S., & Coulombe, P. A. (2007). Intermediate filament scaffolds fulfill mechanical, organizational, and signaling functions in the cytoplasm. Genes & development, 21(13), 1581-1597.
Klein, D. C. (1985). Photoneural regulation of the mammalian pineal gland. Paper presented at the Ciba Found Symp.
Klein, D. C. (2004). The 2004 Aschoff/Pittendrigh lecture: theory of the origin of the pineal gland—a tale of conflict and resolution. Journal of biological rhythms, 19(4), 264-279.