Cancer Medicine. 2018;7:2993–3002. wileyonlinelibrary.com/journal/cam4
|
2993O R I G I N A L R E S E A R C H
Loss of cyclin- dependent kinase- like 2 predicts poor prognosis in gastric cancer, and its overexpression suppresses cells growth and invasion
Chia-Lang Fang
1,2| Yih-Huei Uen
3| Han-Kun Chen
4| You-Cheng Hseu
5,6|
Chih-Chan Lin
7| Shih-Ting Hung
7| Ding-Ping Sun
4,8| Kai-Yuan Lin
7,91Department of Pathology, School of Medicine, College of Medicine, Taipei Medical University, Taipei, Taiwan
2Department of Pathology, Wan Fang Hospital, Taipei Medical University, Taipei, Taiwan
3Department of Surgery, Asia University Hospital, Taichung, Taiwan
4Department of Surgery, Chi Mei Medical Center, Tainan, Taiwan
5Department of Cosmeceutics, China Medical University, Taichung, Taiwan
6Department of Health and Nutrition Biotechnology, Asia University, Taichung, Taiwan
7Department of Medical Research, Chi Mei Medical Center, Tainan, Taiwan
8Department of Nutrition, Chia Nan University of Pharmacy and Science, Tainan, Taiwan
9Department of Biotechnology, Chia Nan University of Pharmacy and Science, Tainan, Taiwan
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
© 2018 The Authors. Cancer Medicine published by John Wiley & Sons Ltd.
Partial findings of this work were presented at The European Society for Medical Oncology Asia 2017 Conference, 17-19 November 2017, Singapore.
Correspondence
Kai-Yuan Lin, Department of Medical Research, Chi Mei Medical Center, Tainan, Taiwan.
Email: [email protected] and
Ding-Ping Sun, Department of Surgery, Chi Mei Medical Center, Tainan, Taiwan.
Email: [email protected] Funding information
Chi Mei Medical Center, Grant/Award Number: 103CM-TMU-10 and 105CM- TMU-10
Abstract
Cyclin- dependent kinase- like 2 (CDKL2), a new member of the cyclin- dependent kinase family, may be involved in gastric cancer (GC) progression. Thus, we conducted this study to explore the clinical effect of CDKL2 in GC. Immunohistochemistry was used to measure CDKL2 levels in gastric tissues. The association of a high CDKL2 level with clinical and pathological characteristics, and the correlation between the CDKL2 level and disease- free and overall survival were analyzed. Transfection was employed to over- express CDKL2 in GC cells and to investigate the effect of CDKL2 overexpression on cell proliferation and invasion. Loss of CDKL2 was positively correlated with several clinical and pathological characteristics, and patients with a low CDKL2 level had sig- nificantly poorer disease- free and overall survival than those with a high level (P = .005 and .001, respectively). Univariate analysis using the Cox proportional hazards model indicated that a low CDKL2 level was a prognosticator for inferior disease- free survival (P = .007). Based on immmunoblotting data, AGS and HGC- 27 GC cells were chosen for CDKL2 overexpression. Cellular studies revealed that CDKL2 overexpression im- paired cell proliferation and invasion. Loss of CDKL2 may serve as a biomarker for predicting GC patient outcomes and a potential therapeutic target for GC treatment.
K E Y W O R D S
CDKL2, gastric cancer, immunohistochemistry, prognosis
1 | INTRODUCTION
Gastric cancer (GC) represents a serious health threat. It is the fourth most common cancer and the third most common cause of cancer- related death worldwide.1 The outcomes of patients with GC remain poor due to a poorly under- stood pathogenesis and lack of novel therapeutic options.2,3 Therefore, identifying potential novel biomarkers may im- prove the prediction of relapse and metastasis and enhance the prognosis and therapeutic responsiveness of patients with GC. GC has evolved among various genetic alterations, and molecular pathology studies may provide an understanding of the molecular variables that cause GC and useful prognos- tic biomarkers.4-10
Cancer is a disease involving uncontrolled cell growth, and carcinogenesis is usually linked to a series of changes in the activity of cell growth regulators.11 Cell cycle progres- sion is strictly modulated by orchestrated actions of cyclins with cyclin- dependent kinases (CDKs).12 The CDK- like (CDKL) family has similar attributes to the CDK family but is not known to bind to cyclins. It includes 5 members (CDKL1 to CDKL5) and is considered a separate branch of the CDK family.13 CDKL1 and CDKL2 are similar and pre- sumably derive from an early vertebrate duplication. cdkl2, located on chromosome 4, was firstly cloned from a human fetal brain.14 Its protein product, CDKL2, accumulates pri- marily in the cytoplasm, with lower levels in the nucleus.
Little is known about the expression and function of CDKLs.
CDKL1 exists predominantly in the brain, lung, kidney, and ovary,13,14 and its overexpression has been observed in glial cells during gliosis.15 Using immunohistochemistry, a study by Kim et al16 reported that the CDKL1 protein level was aug- mented during postnatal heart development in rats. CDKL3 was increased in fast- growing (suspension) HeLa cells, and CDKL3 overexpression in slow- growing (attached) HeLa cells promoted cell proliferation. Moreover, flow cytometric analysis has demonstrated that cells with an insert of cdkl3 could move from the G0/G1 phases to the S phase faster than control cells.17,18 CDKL2 also exists in various brain neurons in mice, and its expression has been reported to be induced in rabbit brains during a learning test. Knockout mice data have indicated a role for CDKL2 in cognitive function.19-22 According to the publicly available Oncomine database, the CDKL2 level in nontumor tissues is higher than that in tumor tissues in all reported cancer types (including brain tumor, colorectal cancer, kidney cancer, lung cancer, and breast cancer). By contrast, one study revealed that CDKL2 was upregulated in breast cancer.23 The expression of CDKL2 in GC is still unknown.
The role of CDKLs in cancer progression has gained increasing attention in recent years. CDKL1 overexpressed is greater in breast cancer tissues than in benign tissues.
CDKL1- knockdown breast cancer cells were reported to
be arrested at the G2/M phase and were more sensitive to cell cycle chemotherapeutic drugs.24 In addition, the CDKL1 level was considerably higher in GC tissues than in paired normal tissues, and CDKL1 silencing in GC cells decreased the amount of proliferating cell nuclear antigen and increased that of Bik pro- apoptotic protein and then suppressed cell proliferation and induced apoptosis.25 Re- analysis of a breast cancer GWAS study suggested that CDKL2 may contribute to cancer. Li et al demonstrated that human mammary gland epithelial cells that expressed CDKL2 had increased epithelial- mesenchymal transition (EMT) and stem cell properties, which were obtained from the activation of a positive feedback loop compris- ing ZEB1, E- cadherin, and β- catenin. Moreover, CDKL2 promoted xenograft proliferation and metastasis in vivo. In particular, CDKL2 is overexpressed in mesenchymal breast cancer cells compared with epithelial cells, and its over- expression is negatively correlated with disease- free sur- vival.23 In summary, the described studies have revealed crucial roles of CDKLs in EMT and carcinogenesis and suggested that CDKLs could be potential biomarkers for prognosis as well as gene targets for cancer therapy. To our knowledge, the role played by CDKL2 in human GC is still unknown.
The aims of this study were to measure the CDKL2 lev- els in normal and GC tissues and cell lines, to evaluate the prognostic effect of CDKL2 in GC, and to study the role of CDKL2 in GC tumorigenicity.
2 | MATERIALS AND METHODS 2.1 | Patients and specimens
Paired GC tissues and adjacent nontumor tissues from 151 patients who underwent surgical resection between 1998 and 2011 at Wan Fang Hospital (Taipei, Taiwan) were collected. All patients with GC in this study received radical total or subtotal gastrectomy with D2 lymph node dissection, serving as a standard radical surgery for GC.
Postoperatively, all patients were evaluated for the ne- cessity of further adjuvant treatment, based on patho- logic TNM staging and prognostic factors. In accordance with the standard practice guideline for GC at Wan Fang Hospital, in this study, patients with T3 or T4 tumors and nodal status of N2 or N3 received postoperative adjuvant chemotherapy. Tumor and nontumor pairs of gastric tis- sues were analyzed for CDKL2 expression. Clinical and pathological characteristics were listed, as provided by the American Joint Committee on Cancer (AJCC) classifica- tion. Disease- free survival was defined as the length of time after surgery during which no relapse is found, based on medical records. Surgically resected tissues from each patient were used to examine CDKL2 levels. All patients
provided written informed consents, and the study was ap- proved by the Institutional Review Board of Wan Fang Hospital (Approval No. 99049). We confirm that all ex- periments were performed in keeping with the relevant guidelines and regulations.
2.2 | Immunohistochemistry
The surgical specimens of gastrectomy were fixed in neutral buffered formalin for 12- 15 hours before sampling blocks.
The representative gastric tissue formalin- fixed paraffin- embedded blocks were used for immunohistochemistry.
Five- micrometer sections were sliced and adhered to micro- scope slides (catalog number: 5196, Muto Pure Chemicals, Tokyo, Japan). Positive control staining for CDKL2 was per- formed using a normal kidney. To retrieve antigen, deparaffi- nized sections were placed in sodium citrate buffer (pH, 6.0;
catalog number: TA00H01, BIOTnA Biotech, Kaohsiung, Taiwan) and boiled for 40 minutes. Five percent of normal goat serum (catalog number: ab7481, Abcam, Cambridge, UK) was applied to block nonspecific staining. The sections were then incubated with the primary antibody (1:100 di- lution in Antibody Diluent (catalog number: S3022, Dako, Glostrup, Denmark) of mouse monoclonal anti- CDKL2 (cat- alogue number: LS- B4479, LifeSpan BioSciences, Seattle, WA) for 2 hours at room temperature. CDKL2 staining was detected using the avidin- biotin- peroxidase complex pro- tocol according to the manufacturer’s instructions (Dako REAL EnVision Detection System, catalog number: K5007, Dako). Diaminobenzidine was used for color development, and hematoxylin was used as a nuclear counterstain. The immunoreactivity of cancer cells and normal gastric glandu- lar epithelial cells was interpreted under a light microscope (Olympus BX51) by a pathologist (CLF) who was blinded to the clinical data. For every case, five 200× fields were evalu- ated and scored and the average immunoreactivity was cal- culated and recorded on a semiquantitative scale: 0 for none, 1 for up to 25%, 2 for 25%- 50%, and 3 for >50% of the tissue
showing positive staining. Sections with a score of 0 or 1 were considered to have low CDKL2 expression, and those with a score of 2 or 3 were considered to have high CDKL2 expression.
2.3 | Cell culture
A human normal gastric cell line (Hs738.St/Int, cata- log number: CRL- 7869) and 4 GC cell lines (AGS, cat- alog number: BCRC60102; TMC- 1, catalog number:
BCRC 60379; HGC- 27, catalog number: 94042256; and 23132/87, catalog number: ACC 201) were obtained from the American Type Culture Collection (ATCC, Manassas, VA), the Bioresource Collection and Research Center (BCRC, Hsinchu, Taiwan), the European Collection of Cell Cultures (ECACC, Salisbury, UK), and Creative Bioarray (Shirley, NY), respectively. All cell lines were authenticated by the ATCC, BCRC, ECACC, and Creative Bioarray cell biology program and were not passaged for longer than 40 passages before original frozen stocks were thawed and used or a new cell aliquot was purchased.
The companies performed cell line characterization using short tandem repeat profiling. Cell lines were maintained in DMEM (Hs738.St/Int; catalog number: 10569- 010, Life Technologies, Grand Island, NY), F- 12K (AGS;
catalog number: 10- 025, Corning, Corning, NY), RPMI- 1640 (TMC- 1 and HGC- 27; catalog number: A1049101, Life Technologies), and MEM (23132/87; catalog num- ber: 10- 009, Corning) with 10% fetal bovine serum (cata- log number: 04- 001- 1A, Biological Industries, Cromwell CT), 100 units/mL penicillin G, 100 μg/mL streptomycin sulfate, and 250 ng/mL amphotericin B (catalog number:
15240- 062, Life Technologies).
2.4 | Whole protein extraction
The cells were lysed in the protease/phosphatase- containing RIPA Protein Extraction Reagent (catalog number: 89900,
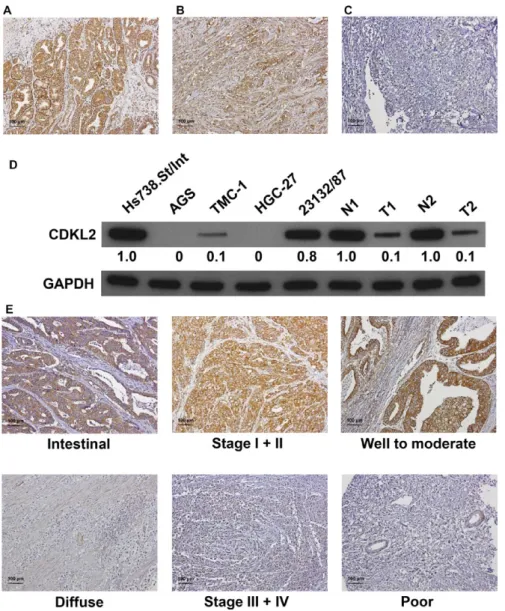
TABLE 1 Demographic data and survival in different stages of GC according to the AJCC classification
Stage I (n = 26) Stage II (n = 38) Stage III (n = 69) Stage IV (n = 18) Total (n = 151) Gender of patients
Male 16 25 48 11 100
Female 10 13 21 7 51
Age of patients (y)a 67.0 (12.1) 75.3 (10.2) 70.0 (12.8) 59.0 (14.9) 69.5 (13.1)
Follow- up period
(d)a 1575.0 (1142.7) 946.0 (696.5) 819.5 (822.5) 360.3 (285.8) 926.7 (877.5)
Survival
Yes 17 18 15 2 52
No 9 20 54 16 99
aAge of patients and follow- up period are expressed as the mean (SD).
Pierce Biotechnology, Rockford, IL) according to the manu- facturer’s protocol. The protein concentration was assayed with the BCA Protein Assay Kit (catalog number: 23225, Pierce Biotechnology), with bovine serum albumin serving as a standard. The whole proteins were frozen at −80°C be- fore immunoblotting.
2.5 | Immunoblotting
Ten percent SDS- PAGE gels were used to separate pro- teins (30 μg/well). After electrophoresis, the separated pro- teins were transferred to nitrocellulose membranes (catalog number: NBA085C001EA, PerkinElmer, Waltham, MA) using a wet transfer method. The membranes were blocked with 5% nonfat milk (catalog number: sc- 2324, Santa Cruz Biotechnology, Dallas, TX) in 1× PBS (catalog number:
21- 040, Life Technologies) and incubated at 4°C overnight.
The membranes were then incubated with a different mouse monoclonal anti- CDKL2 antibody (1:100 dilution in 5%
milk/0.05% 1× PBST; catalog number: H00008899- M01;
Abnova, Taipei, Taiwan) at room temperature for 1 hour.
Subsequently, the peroxidase- conjugated secondary antibod- ies (1:100 000 dilution in 5% milk/0.05% 1× PBST; cata- log number: A4416, Sigma, Saint Louis, MO) were added to the membranes for 45 minutes at room temperature. The protein bands were detected using the Western Lighting ECL Ultra Chemiluminescence Substrate (catalog num- ber: NEL113001EA, PerkinElmer) and analyzed with Fuji Image Gauge software (Fuji Photo Film Co., Tokyo, Japan).
GAPDH acted as a protein loading control.
2.6 | Transfection
Commercial human CDKL2 cDNA ORF or empty vectors (catalog numbers: RC510780 and PS100001, OriGene, Rockville, MD, USA) were transfected into AGS and HGC- 27 cells using TurboFect transfection reagent accord- ing to the manufacturer’s instructions (catalog number:
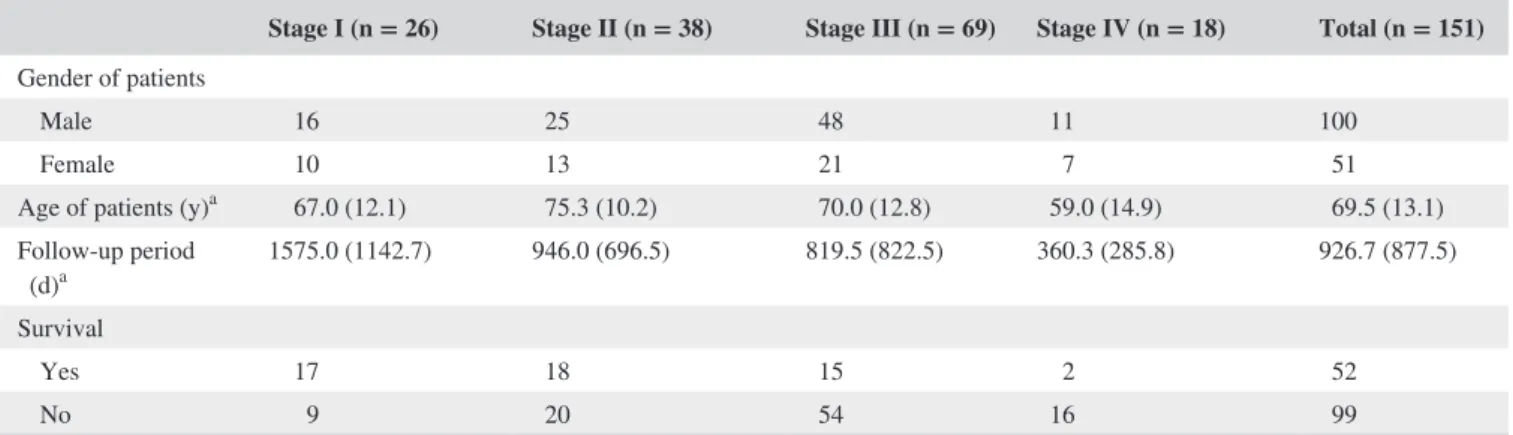
FIGURE 1 CDKL2 expression in gastric tissues and cell lines. Panels A to C. Gastric tissue specimens analyzed by immunohistochemistry with an antibody against CDKL2. Panel A shows a sample of nontumor tissue with high CDKL2 expression; Panel B shows a tumor specimen with low CDKL2 expression; Panel C shows a tumor specimen without CDKL2 expression. D, CDKL2 protein expression was examined in 5 gastric cell lines and 2 nontumor/tumor pairs of gastric tissues.
N, nontumor; T, tumor. E, Representative CDKL2 staining for different parameters.
The blots were first hybridized with CDKL2 antibody and, after stripping, rehybridized with β- actin antibody. The immunoblots in the figure were cropped
R0531, Thermo Scientific, Waltham, MA). Briefly, the cells were seeded in a 6- cm dish in complete medium 24 hours before transfection. The cells were then tran- siently transfected with 6 μg CDKL2 cDNA ORF or empty vector for 24 hours. G418 (500 μg/mL; catalog number:
G8168, Sigma)- resistant stable clones were selected.
Immunoblotting was performed to evaluate the transfec- tion efficiency.
2.7 | Colony formation assay
Cells with a density of 500 cells per well were seeded into 6- well plates. After 12 days postplating, individual colonies (>50 cells/colony) were fixed, stained with 1% crystal violet/
methanol solution, and then scanned with a Scanjet 2200c scanner (HP, Palo Alto, CA). Next, methanol was added to solubilize the crystal violet within the cells. The absorbance was then detected at a wavelength of 540 nm to quantify the number of colonies formed. The assay was performed in trip- licate, and the data are shown as the mean ± the standard de- viation (SD).
2.8 | In vitro invasion assay
The invasive capacity of the cells was determined using a Cell Invasion Assay Kit (catalog number: ECM55, Merck Millipore, Darmstadt, Germany) according to manufacturer’s protocol.
Briefly, serum- free media containing 2 × 105 cells were added to ECMatrix- layered cell culture inserts (containing polycar- bonate membranes with an 8 μm pore size) after 24- well plates were filled with complete media. The cells were then cultured for 24 hours. After incubation, the media and noninvasive cells were removed. The inserts were then dipped in the staining so- lution (containing crystal violet) to stain invaded cells on the lower surface of the membranes. The cultures were photo- graphed (100× magnification, using a Leica DMIRB micro- scope), and the number of invaded cells was counted. The assay TABLE 2 CDKL2 expression in GC and its correlation with
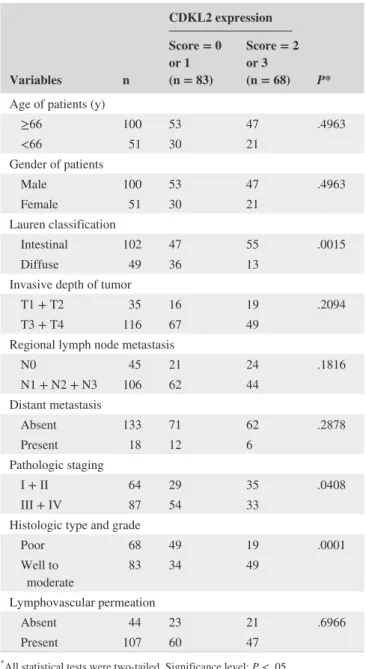
clinical and pathological parameters
Variables n
CDKL2 expression
P*
Score = 0 or 1 (n = 83)
Score = 2 or 3 (n = 68) Age of patients (y)
≥66 100 53 47 .4963
<66 51 30 21
Gender of patients
Male 100 53 47 .4963
Female 51 30 21
Lauren classification
Intestinal 102 47 55 .0015
Diffuse 49 36 13
Invasive depth of tumor
T1 + T2 35 16 19 .2094
T3 + T4 116 67 49
Regional lymph node metastasis
N0 45 21 24 .1816
N1 + N2 + N3 106 62 44
Distant metastasis
Absent 133 71 62 .2878
Present 18 12 6
Pathologic staging
I + II 64 29 35 .0408
III + IV 87 54 33
Histologic type and grade
Poor 68 49 19 .0001
Well to
moderate 83 34 49
Lymphovascular permeation
Absent 44 23 21 .6966
Present 107 60 47
*All statistical tests were two- tailed. Significance level: P < .05.
FIGURE 2 Survival analysis of patients with GC stratified by CDKL2 immunoreactivity. Panel A shows disease- free survival. Patients with low CDKL2 expression had a 5- y disease- free survival rate of 34.4% compared with 58.0% for patients with high CDKL2 expression. Panel B shows overall survival. Patients with low CDKL2 expression had a 5- y overall survival rate of 19.5% compared with 42.5% for patients with high CDKL2 expression. Two- tailed P < .05 was considered statistically significant
was conducted 3 times independently, and the results are pre- sented as the mean ± SD.
2.9 | Statistical analysis
Critical clinical and pathological characteristics were analyzed, which are listed as follows: age and sex of patients, Lauren clas- sification, invasive depth of tumor, regional lymph node me- tastasis, distant metastasis, pathologic staging, histologic type
and grade, and lymphovascular permeation. The chi- square test was used to analyze the association between CDKL2 level and each clinical and pathological characteristic. Survival curves, namely disease- free survival and overall survival, were plotted using the Kaplan- Meier method, and differences in disease- free survival were determined using the univariate log- rank test. A statistically significant difference was defined as a P value of less than .05. Characteristics demonstrating signifi- cant P values from the univariate analysis were entered into the
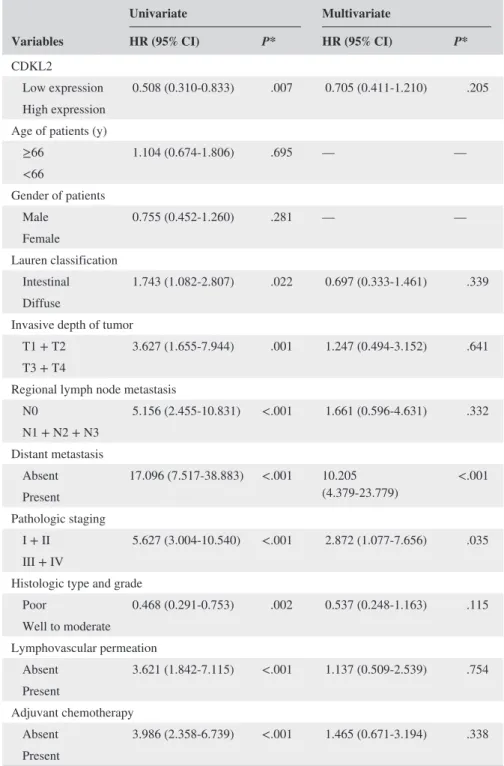
Variables
Univariate Multivariate
HR (95% CI) P* HR (95% CI) P*
CDKL2
Low expression 0.508 (0.310- 0.833) .007 0.705 (0.411- 1.210) .205 High expression
Age of patients (y)
≥66 1.104 (0.674- 1.806) .695 — —
<66
Gender of patients
Male 0.755 (0.452- 1.260) .281 — —
Female
Lauren classification
Intestinal 1.743 (1.082- 2.807) .022 0.697 (0.333- 1.461) .339 Diffuse
Invasive depth of tumor
T1 + T2 3.627 (1.655- 7.944) .001 1.247 (0.494- 3.152) .641 T3 + T4
Regional lymph node metastasis
N0 5.156 (2.455- 10.831) <.001 1.661 (0.596- 4.631) .332 N1 + N2 + N3
Distant metastasis
Absent 17.096 (7.517- 38.883) <.001 10.205
(4.379- 23.779) <.001 Present
Pathologic staging
I + II 5.627 (3.004- 10.540) <.001 2.872 (1.077- 7.656) .035 III + IV
Histologic type and grade
Poor 0.468 (0.291- 0.753) .002 0.537 (0.248- 1.163) .115 Well to moderate
Lymphovascular permeation
Absent 3.621 (1.842- 7.115) <.001 1.137 (0.509- 2.539) .754 Present
Adjuvant chemotherapy
Absent 3.986 (2.358- 6.739) <.001 1.465 (0.671- 3.194) .338 Present
HR, hazard ratio; CI, confidence interval.
aThis table shows disease- free survival.
*All statistical tests were two- tailed. Significance level: P < .05.
TABLE 3 Univariate and multivariate analyses of prognostic biomarkers in 151 patients with GCa
multivariate Cox regression model, and the hazard ratio (HR) and independence of prognostic impact were calculated in a stepwise backward fashion. A two- tailed Student’s t test was used to analyze the differences in cell proliferation and inva- sion between CDKL2 overexpressed cells and control cells.
All statistical analyses were performed using SPSS 24.0 soft- ware (IBM, New York, NY).
3 | RESULTS
3.1 | Basic data of patients
In total, 151 patients with GC—100 men and 51 women—
were enrolled in this study (Table 1). The mean age for all patients at first diagnosis was 69.5 years (ranging from 34 to 96 years). According to the AJCC classification, 26 stage I patients, 38 stage II patients, 69 stage III patients, and 18 stage IV patients were present. The mean follow- up time for all patients was 926 days (ranging from 5 to 3709 days).
During follow- up, 99 patients died.
3.2 | Correlation between downregulation of CDKL2 and clinical and pathological
characteristics in GC
Immunohistochemical analysis was employed to examine the CDKL2 level in GC tissues (Figure 1A- C). The examined GC tissues showed negative or weak CDKL2 expression, whereas the nontumor tissues had strongly positive CDKL2 expression (P < .001). Among the 151 tumor and nontumor pairs, 68 GC tissues (45.0%) showed low CDKL2 levels (scores of 0 or 1) and 89 nontumor tissues (58.9%) showed high CDKL2 levels (scores of 2 or 3). Immunoblotting confirmed that the CDKL2 level was substantially decreased in the GC tissues and cell lines compared with normal tissues and cell lines (Figure 1D).
Table 2 shows that low CDKL2 expression was correlated with Lauren classification, pathologic staging and histologic type, and grade (P = .0015, .0408, and .0001, respectively).
Representative photographs of CDKL2 expression for differ- ent characteristics are shown in Figure 1E. Other clinical and pathological characteristics were found not to be significantly correlated with the CDKL2 level (Table 2).
3.3 | Loss of CDKL2 is a poor prognosticator for GC
The correlations of the patients’ outcomes with CDKL2 ex- pression are shown in Figure 2. Patients with low CDKL2 expression had significantly poorer disease- free survival and overall survival than patients with high CDKL2 expression (Figure 2A, B, P = .005 and .001, respectively). The 5- year disease- free survival rate of patients with low CDKL2 levels was 0.344 (95% confidence interval [CI] 0.2068 to 0.4812), whereas that of patients with high CDKL2 levels was 0.580 (95% CI 0.4506 to 0.7094). The 5- year overall survival rate of patients with low CDKL2 levels was 0.195 (95% CI 0.0911 to 0.2989), whereas that of patients with high CDKL2 levels was 0.425 (95% CI 0.3015 to 0.5485).
FIGURE 3 Verification of CDKL2 overexpression in AGS and HGC- 27 cells and the effect of CDKL2 overexpression on cell proliferation and invasion. The immunoblotting results (A) indicate that CDKL2 was efficiently overexpressed by transfection.
The blots were first hybridized with CDKL2 antibody and, after stripping, rehybridized with β- actin antibody. The immunoblots in the figure were cropped. B, CDKL2 overexpression suppressed cell proliferation. The histogram represents OD540 (presented as mean ± SD). The assay was performed 3 times. Red circles were used to show individual values. Significance level: P < .05. C, CDKL2 overexpression repressed cell invasion. The histogram represents cell invasion (presented as mean ± SD). The assay was performed 3 times.
Red circles were used to show individual values. Significance level:
P < .05. The differences in cell proliferation and invasion between CDKL2 overexpressed and control cells were analyzed using Student’s t test. Significance level: P < .05
Adjuvant chemotherapy was added as a variable, and data of the univariate analysis of the prognostic biomarkers of GC are summarized in Table 3. Loss of CDKL2 (P = .007), Lauren classification (P = .022), invasive depth of tumor (P = .001), regional lymph node metastasis (P < .001), dis- tant metastasis (P < .001), pathologic staging (P < .001), histologic type and grade (P = .002), lymphovascular per- meation (P < .001), and adjuvant chemotherapy (P < .001) were significantly correlated with disease- free survival. The multivariate analysis demonstrated that only distant metasta- sis (HR 9.470, 95% CI 4.092 to 21.918, P < .001) remained as an independent prognostic biomarker, even after other prognostic biomarkers were controlled for. Loss of CDKL2, however, was not an independent prognostic biomarker (HR 0.705, 95% CI 0.411 to 1.210, P = .205) (Table 3).
3.4 | CDKL2 overexpression inhibited cell proliferation and invasion in GC cells
To determine the effect of CDKL2 overexpression on cell pro- liferation, 2 GC cell lines with low CDKL2 levels—AGS and HGC- 27 cells—were transfected with CDKL2 cDNA ORF vectors to generate CDKL2- overexpressed cells (Figures 1D and 3A). According to our results, compared with control vectors, the proliferative abilities of AGS and HGC- 27 cells transfected with CDKL2 cDNA ORF vectors were signifi- cantly impaired (Figure 3B). Finally, the role of CDKL2 in the invasiveness of the AGS and HGC- 27 cells was investi- gated. We also found that cell invasion was significantly in- hibited in the CDKL2- overexpressed cells than in the control
cells (Figure 3C). These results indicate that CDKL2 overex- pression hinders GC cell proliferation and invasion in vitro.
4 | DISCUSSION
Similar to most cancers, GC has a molecular genetic basis that depends on the abnormalities in normal cellular regu- latory mechanisms that govern cell proliferation.26 In this study, we measured CDKL2 expression in GC and analyzed the relationship between CDKL2 expression and different clinical and pathological characteristics. Our results reveal a significant downregulation of the CDKL2 protein in human GC cells and tissues, and the decreased CDKL2 level was positively correlated with Lauren classification, pathologic staging, histologic type and grade, and short patient sur- vival. Furthermore, CDKL2 downregulation is an unfavora- ble prognosticator for GC, and forced CDKL2 expression in human GC cell lines hindered cell proliferation and impaired invasiveness.
Studies on CDKL expression in various cancers are scarce and controversial. Jones et al described a gene signature in which CDKL1 was downregulated in renal cell cancer, but Qin et al revealed that CDKL1 was upregulated in colorec- tal cancer.27,28 Kawahara and colleagues demonstrated that CDKL5 was overexpressed in leukemia cells but not in normal T cells.29 To date, only one study performed by Li and col- leagues showed the expression of CDKL2 and indicated that CDKL2 was considerably overexpressed in human breast can- cer tissues and cells compared with normal breast tissues and
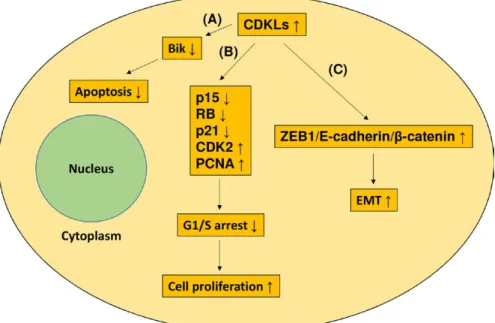
FIGURE 4 Cellular effects of CDKL overexpression in GC. CDKL overexpression can elicit 3 possible effects. A, CDKL overexpression can inhibit apoptosis through inactivating the Bik pro- apoptotic protein. B, CDKL overexpression can downregulate the p15, RB, and p21 expression, upregulate CDK2 and PCNA expression, and then, promote G1- S transition and cell proliferation. C, CDKL2 can facilitate EMT through activating a positive feedback loop comprising ZEB1, E- cadherin, and β- catenin
cells.23 By contrast, our data show the loss of CDKL2 in GC.
Our data are in line with other data described in the Oncomine database. One reason for the discrepancy between our study and Li’s study may result from the different molecules exam- ined. In our study, CDKL2 protein was measured, and in an- other, CDKL2 mRNA was detected. Another explanation for the discrepancy is that the CDKL2 expression is cell context- specific. However, notably, even in breast cancer, the data from Li’s study are different from those described in the Oncomine database. The reason for the discrepancy between Li’s study and the Oncomine database may result from the different sam- ple size. Overall, these studies suggested that the expression of CDKLs in human cancers seems to be more complicated than expected and warrants additional studies. This is the first study to report the expression of CDKL2 in GC.
The CDK family is crucial in the regulation of cell cycle progression at the G1/S and G2/M checkpoints.30 The CDKL family, which is considered separate from the CDK family, was recently identified through biochemical and genetic ap- proaches.13 The role that CDKLs play in cancer is not fully understood. Several studies have shown that CDKLs were potential oncogenes and had roles in tumor development, and these results are summarized in Figure 4. For instance, cell growth, tumor invasion, and cell cycle progression of colorectal cells were considerably hindered through CDKL1 silencing.28 CDKL1 was also revealed to downregulate the expression of p15 and RB and then promote G1- S transition.
Similar results were also observed in melanoma: Suppression of CDKL1 in melanoma cells considerably delayed cell growth, induced cell apoptosis, and stopped cell cycle pro- gression at the G1 phase.31 Furthermore, CDKL1 was shown to facilitate cell cycle progression through decreasing the ex- pression of p21 and increasing the expression of CDK2. One study conducted on GC demonstrated that CDKL1 knock- down decreased cellular proliferation and increased apopto- sis.25 Moreover, CDKL1 was shown to inhibit the activation of Bik pro- apoptotic protein and enhance the expression of PCNA. CDKL2- expressing human mammary epithelial cells enhanced EMT and stem cell properties. In addition, CDKL2 promoted xenograft proliferation and metastasis in vivo.23 CDKL2 was reported to facilitate EMT by activat- ing a positive feedback loop comprising ZEB1, E- cadherin, and β- catenin. However, in this study, CDKL2 functioned as a tumor suppressor and enforced expression of CDKL2 in- hibited GC cell proliferation and invasion. According to the publicly available Oncomine database, the CDKL2 level in nontumor tissues is higher than that in tumor tissues in sev- eral cancers. The mechanisms that make CDKLs to be tumor suppressors have not been reported. These mechanisms must be elucidated in additional studies.
Few studies exist to suggest the effects of CDKLs on tumor prognosis. A study demonstrated that an increased copy number of cdkl4 in colorectal cancer was predictive
of poorer patient survival.32 Varghese’s group reported that CDKL5 overexpression was associated with poor prognosis for patients with glioblastoma.33 The only study conducted on breast cancer showed that patients with upregulated CDKL2 expression had a significantly poorer survival rate compared with patients without this change.23 However, our results in- dicate that loss of CDKL2 was negatively correlated with patient survival. In accordance with the aforementioned ex- planation of the expression profiles of CDKL2 in breast can- cer and GC, the reason for the discrepancy may result from the different molecules examined. Furthermore, multivariate Cox regression analysis revealed that loss of CDKL2 was not an independent prognostic biomarker. It suggested that loss of CDKL2 in GC can be a useful prognostic predictor in con- junction with other conventional important prognostic factors such as pathologic stage and distant metastasis. A group of advanced GC patients with low CDKL2 expression should be considered for aggressive treatment and be clinically eval- uated and followed up more closely. This is the first study to reveal that loss of CDKL2 is a prognostic biomarker for GC.
In conclusion, our findings indicate that loss of CDKL2 promotes a malignant phenotype of GC and illustrate the clinical significance of loss of CDKL2 in GC.
ACKNOWLEDGMENTS
This work was supported by grants of Chi Mei Medical Center (103CM- TMU- 10 and 105CM- TMU- 10).
CONFLICT OF INTEREST
The authors have no conflict of interest to declare.
ORCID
Kai-Yuan Lin http://orcid.org/0000-0001-8225-8221
REFERENCES
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A.
Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87‐108.
2. Fuchs CS, Mayer RJ. Gastric carcinoma. N Engl J Med.
1995;333:32‐41.
3. Hartgrink HH, Jansen EP, van Grieken NC, van de Velde CJ.
Gastric cancer. Lancet. 2009;374:477‐490.
4. Jin ZL, Jiang WH, Wang LW. Biomarkers for gastric can- cer: progression in early diagnosis and prognosis. Oncol Lett.
2015;9:1502‐1508.
5. Lin KY, Wang LH, Hseu YC, et al. Clinical significance of in- creased guanine nucleotide exchange factor Vav3 expression in human gastric cancer. Mol Cancer Res. 2012;10:750‐759.
6. Fang CL, Hseu YC, Lin YF, et al. Clinical and prognostic associ- ation of transcription factor SOX4 in gastric cancer. PLoS ONE.
2012;7:e52804.
7. Fang CL, Hseu YC, Lin YF, et al. Clinicopathological correlation and prognostic significance of protein kinase Cα overexpression in human gastric carcinoma. PLoS ONE. 2013;8:e56675.
8. Sun DP, Fang CL, Chen HK, et al. EPAC1 overexpression is a prognostic marker and its inhibition shows promising therapeutic potential for gastric cancer. Oncol Rep. 2017;37:1953‐1960.
9. Fang CL, Sun DP, Chen HK, et al. Overexpression of mitochon- drial GTPase MFN2 represents a negative prognostic marker in human gastric cancer and its inhibition exerts anti- cancer effects.
J Cancer. 2017;8:1153‐1161.
10. Lin CC, Fang CL, Sun DP, et al. High expression of mitochon- drial intermembrane chaperone TIMM9 represents a nega- tive prognostic marker in gastric cancer. J Formos Med Assoc.
2017;116:476‐483.
11. Wu WK, Cho CH, Lee CW, et al. Dysregulation of cellular signal- ing in gastric cancer. Cancer Lett. 2010;295:141‐153.
12. Malumbres M, Harlow E, Hunt T, et al. Cyclin- dependent ki- nases: a family portrait. Nat Cell Biol. 2009;11:1275‐1276.
13. Meyerson M, Enders GH, Wu CL, et al. A family of human cdc2- related protein kinases. EMBO J. 1992;11:2909‐2917.
14. Taglienti CA, Wysk M, Davis RJ. Molecular cloning of the epi- dermal growth factor- stimulated protein kinase p56 KKIAMRE.
Oncogene. 1996;13:2563‐2574.
15. Yen SH, Kenessey A, Lee SC, Dickson DW. The distribution and biochemical properties of a Cdc2- related kinase, KKIALRE, in normal and Alzheimer brains. J Neurochem. 1995;65:2577‐2584.
16. Kim SO, Katz S, Pelech SL. Expression of second messenger- and cyclin- dependent protein kinases during postnatal develop- ment of rat heart. J Cell Biochem. 1998;69:506‐521.
17. Jaluria P, Betenbaugh M, Konstantopoulos K, Shiloach J.
Enhancement of cell proliferation in various mammalian cell lines by gene insertion of a cyclin- dependent kinase homolog. BMC Biotechnol. 2007;7:71.
18. Szyniarowski P, Corcelle-Termeau E, Farkas T, et al. A compre- hensive siRNA screen for kinases that suppress macroautophagy in optimal growth conditions. Autophagy. 2011;7:892‐903.
19. Sassa T, Gomi H, Sun W, Ikeda T, Thompson RF, Itohara S.
Identification of variants and dual promoters of murine ser- ine/threonine kinase KKIAMRE. J Neurochem. 2000;74:
1809‐1819.
20. Sassa T, Gomi H, Itohara S. Postnatal expression of Cdkl2 in mouse brain revealed by LacZ inserted into the Cdkl2 locus. Cell Tissue Res. 2004;315:147‐156.
21. Gomi H, Sun W, Finch CE, Itohara S, Yoshimi K, Thompson RF.
Learning induces a CDC2- related protein kinase, KKIAMRE. J Neurosci. 1999;19:9530‐9537.
22. Gomi H, Sassa T, Thompson RF, Itohara S. Involvement of cyclin- dependent kinase- like 2 in cognitive function required for contextual and spatial learning in mice. Front Behav Neurosci.
2010;4:17.
23. Li L, Liu C, Amato RJ, Chang JT, Du G, Li W. CDKL2 promotes epithelial- mesenchymal transition and breast cancer progression.
Oncotarget. 2014;5:10840‐10853.
24. Tang L, Gao Y, Yan F, Tang J. Evaluation of cyclin- dependent kinase- like 1 expression in breast cancer tissues and its regulation in cancer cell growth. Cancer Biother Radiopharm. 2012;27:392‐398.
25. Sun W, Yao L, Jiang B, Shao H, Zhao Y, Wang Q. A role for Cdkl1 in the development of gastric cancer. Acta Oncol.
2012;51:790‐796.
26. Kim JY, Jeon TJ, Bae BN, et al. The prognostic significance of growth factors and growth factor receptors in gastric adenocarci- noma. APMIS. 2013;121:95‐104.
27. Jones J, Otu H, Spentzos D, et al. Gene signatures of progres- sion and metastasis in renal cell cancer. Clin Cancer Res.
2005;11:5730‐5739.
28. Qin C, Ren L, Ji M, et al. CDKL1 promotes tumor prolifer- ation and invasion in colorectal cancer. Onco Targets Ther.
2017;10:1613‐1624.
29. Kawahara M, Hori T, Matsubara Y, Okawa K, Uchiyama T.
Cyclin- dependent kinase like 5 is a novel target of immunother- apy in adult T- cell leukemia. J Immunother. 2007;30:499‐505.
30. Morgan DO. Principles of CDK regulation. Nature.
1995;374:131‐134.
31. Song Z, Lin J, Sun Z, Ni J, Sha Y. RNAi- mediated downregu- lation of CDKL1 inhibits growth and colony- formation ability, promotes apoptosis of human melanoma cells. J Dermatol Sci.
2015;79:57‐63.
32. Lin M, Zhang Y, Li A, et al. High- throughput RNAi screening of human kinases identifies predictors of clinical outcome in colorectal cancer patients treated with oxaliplatin. Oncotarget.
2015;6:16774‐16785.
33. Varghese RT, Liang Y, Guan T, Franck CT, Kelly DF, Sheng Z.
Survival kinase genes present prognostic significance in glioblas- toma. Oncotarget. 2016;7:20140‐20151.
SUPPORTING INFORMATION
Additional supporting information may be found online in the Supporting Information section at the end of the article.
How to cite this article: Fang C-L, Uen Y-H, Chen H-K, et al. Loss of cyclin- dependent kinase- like 2 predicts poor prognosis in gastric cancer, and its overexpression suppresses cells growth and invasion. Cancer Med. 2018;7:2993–3002.
https://doi.org/10.1002/cam4.1577