1
Sclareol exhibits antiinflammatory activity in both lipopolysaccharide-stimulated macrophages and the λ-carrageenan-induced paw edema model
Guan-Jhong Huang,† Chun-Hsu Pan,‡ and Chieh-Hsi Wu‡,*
Institute of Chinese Pharmaceutical Sciences, China Medical University, Taichung 40402, Taiwan;
School of Pharmacy, China Medical University, Taichung 40402, Taiwan
* To whom correspondence should be addressed. Tel: +886-4-22053366#5101; Fax:
+886-4-22073709; E-mail: [email protected].
† Institute of Chinese Pharmaceutical Sciences, China Medical University
‡ School of Pharmacy, China Medical University
2
Sclareol is a natural fragrance compound and being widely used in the cosmetic and food
industries. The lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages and the
λ-carrageenan (Carr)-induced edema paw models were applied to examine the antiinflammatory property of Sclareol (1) and its possible molecular mechanisms. Experimental results
demonstrated that 1 inhibited cell growth, nitric oxide (NO) production, and inducible nitric
oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) proteins expression in LPS-stimulated
macrophages. Compound 1 also reduced paw edema, tissue content of NO, tumor necrosis factor-alpha (TNF-) and malondialdehyde (MDA), iNOS and COX-2 proteins expression, and neutrophil infiltration within the tissues by Carr stimulation. The present study suggested that the
antiinflammatory mechanisms of 1 might be related to the decrease of the inflammatory cytokine
and the increase of antioxidant enzymes activity, which will subsequently reduce inflammatory
responses.
3
The acute inflammatory response is a series of local cellular and vascular responses that occurs
immediately following tissue damage, and this complex biological response is a protective
mechanism of organisms to remove the injurious stimuli, such as pathogens, irritants or physical
injury, from the tissues and to initiate the healing process. However, chronic inflammation has
been reported to involve in the development of several diseased conditions or disorders such as
Alzheimer disease,1 asthma,2 atherosclerosis,3 autoimmune diseases,4 cancers5 and rheumatoid
arthritis,6 which may lead to progressive destruction of the tissue, fibrosis, and necrosis.7, 8
Numerous molecules have been mentioned to contribute the local tissue destruction during
chronic inflammation.9-11 Of these, inducible nitric oxide synthase (iNOS), a member of the NOS
protein family, catalyzes the formation of nitric oxide (NO) from L-arginine.12 NO can activate
guanylate cyclase to induce smooth muscle relaxation in the normal physiological condition.
High-output NO produced by the activated macrophage via iNOS has been found to play a major
role as antimicrobial molecule.13 However, highly level of NO have the opportunity to react with
superoxide resulting in peroxynitrite formation and cell toxicity, which are found to play
important roles in inflammation and carcinogenesis. The expression of COX-2 (cyclooxygenase 2)
has also been mentioned to implicate the response for the prostaglandin biosynthesis involved in
inflammation and pain, and clinical application of highly selective inhibitors of COX-2 has been
demonstrated to provide effective antiinflammatory activity with marked reduction in
4
gastrointestinal toxicity as compared to traditional NSAIDs (non-steroidal antiinflammatory
drugs).14 Similarly, TNF- (tumor necrosis factor-alpha), an endotoxin-induced glycoprotein, is a
critical modulator of host immune response to infection, but inappropriate or excessive
production can be harmful. Receiving anti-TNF- antibody and oral administration of soluble TNF receptors have been demonstrated to control the inflammatory conditions.11
Compound 1 (labd-14-ene-8,13-diol) belongs to the member of labdane type diterpenes and
first isolated from the plant Salvia sclarea L.(Lamiaceae), whose natural fragrance ingredient was
widely applied in both cosmetic and food industries. Several studies demonstrated that 1 has been
identified as a biologically active molecule to provide the cytotoxic or cytostatic effects against
numerous human cancer cell lines.15-18 Noori et al. noticed that 1 modulated the immune response
through shifting cytokines pattern in the splenocytes obtained from intra-tumorally injected
mice.19 Although this report implicated that 1 might has potential activity to regulate the
inflammatory response, the possible pharmacological mechanisms are unclear until now. Thereby,
the present study designed both in vitro and in vivo study to examine whether 1 has potential
effects against inflammatory response in the lipopolysaccharide (LPS)-stimulated macrophages and the -carrageenan-induced edema paw model, and to clarify its possible molecular
mechanisms, which will help us to further evaluate the clinical therapeutic potential of 1 on
antiinflammation.
5
RESULTS AND DISCUSSION
Effect of Sclareol (1) on Cell Viability of Macrophage. The growth regulation of 1 on
macrophages cell viability was examined by a MTT assay (Figure S1, Supporting Information).
The cells were pre-treated with 1 at the concentrations (0, 1, 5 and 10 g/mL) for 1 h and then co-incubated with 100 ng/mL of LPS for further 24 h. The results indicated that 100 ng/mL of
LPS did not change cell viability of macrophages. Similarly, the cell viability of macrophages
also did not be influenced even if treating the highest concentration (10 μg/mL) of 1 with
presence of LPS. However, several studies reported that 1 can markedly reduce the cell viability
of numerous human leukemic cell lines at a range of IC50 value from 6.0 to 24.2 g/mL,15, 18
inhibit human breast cancer cells at a range of IC50 value from 7.0 to 11.6 g/mL,17, 18 and suppress human colon cancer cells (HCT116) at 10.6 g/mL of IC50 value.18 In the normal cell such as peripheral blood mononuclear (PBMC) cells, 1 still exhibited its growth inhibition at IC50
value of 10.7 g/mL.18
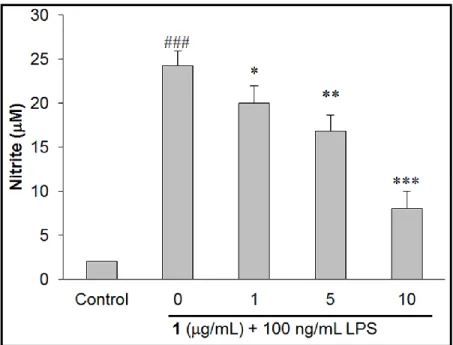
Inhibitory Effect of Sclareol (1) on LPS-Stimulated NO, iNOS and COX-2. Various concentration (0, 1, 5 and 10 g/mL) of 1 were used on macrophages to test whether 1 can reverse LPS-induced dramatically accumulation of NO (Figure 1). The results revealed that 100
ng/mL LPS can evidently increase NO production as compared with control group (p < 0.001),
6
and this effect can be markedly suppressed in a dose-dependent manner by pre-treatment of 1 (1,
5 and 10 g/mL) as compared to those in LPS treated only group. Additionally, 1 (0, 1, 5 and 10
g/mL) was also tested on macrophages to examine whether it can reduce protein expression of inflammation-associated molecules triggered by LPS (Figure 2). The experimental results
suggested that 100 ng/mL of LPS can significantly stimulated protein expression of iNOS and COX-2 (p < 0.001), and pre-treatment of 1 at concentration of 5 and 10 g/mL can obviously down-regulate expression of these LPS-stimulated proteins as compared to LPS treated only
group (p < 0.01 and p < 0.001). It has been mentioned that 1 can increase expression of
interferon-gamma (INF-γ) and decrease interleukin-4 (IL-4) in the splenocytes isolated from
intra-tumorally injected mice.19 Chan and Riches demonstrated that co-incubation of LPS and
INF-γ can induce both TNF-α and IL-1β production, which up-regulated protein expression of
iNOS via activating ERK and JNK MAPK pathways in the RAW 264.7γNO(−) cells that do not
produce NO with IFN-γ stimulation alone.20 Roach et al. also reported that co-treatment of LPS
and INF-γ will increase both transcriptional and translational levels of iNOS in the
macrophages.21 Moreover, the up-regulatory effect of INF-γ on the expression of iNOS also has
been identified in aortic smooth muscle cells, epithelial cells, and saphenous vein
endothelium.22-24 In human foreskin fibroblasts, IFN-γ has been reported to suppress COX-2
promoter activity.25 On the other hand, IL-4 has been evidenced to down-regulate
7
IFN-γ-inducible iNOS transcription in the murine macrophage cell line RAW264.7.26 Cui et al.
also noticed that IL-4 has the capacity to inhibit COX-2 mRNA transcription in human non-small
cell lung cancer and follicular dendritic cells.27, 28 In contrast to the regulation of IFN-γ, Guo et al.
noticed that IL-4 played a critical role for mRNA stabilization of iNOS in primary human airway
epithelial cells stimulated by IFN-γ.29 To date, it is still unknown whether 1 can modulate the
expression of INF-γ and IL-4 in the macrophages. In the present study, experimental data
suggested that 1 can markedly decrease LPS-stimulated iNOS expression and then reduce NO
production in the macrophages.
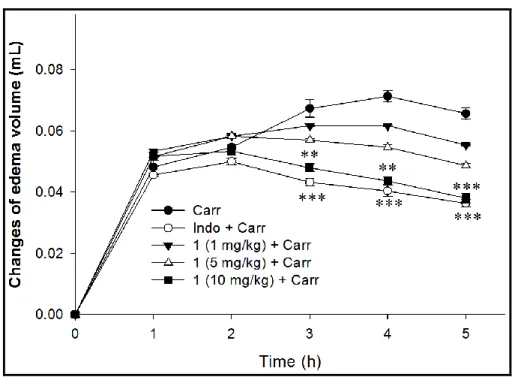
Inhibitory Effect of Sclareol (1) on Carr-Induced Paw Edema. Carr-induced paw edema
model was used to evaluate the in vivo antiinflammatory effect of 1 (Figure 3). The results
showed that Carr injection will stimulate local inflammation and then induce edema of paw
tissues. Indo, a common clinical NSAID, was used as positive control to indicate that
pre-treatment of 10 mg/kg Indo can effectively reduce paw edema after 3 h Carr stimulation (p <
0.001). Similarly, pre-treatment of 10 mg/kg of 1 can also markedly attenuate paw edema after 3
h Carr stimulation, as same as the result of Indo + Carr group (p < 0.01). Sclareol belongs to a
diterpene alcohol that exhibits hormone-like property similar to that of estrogen according to the
structural similarity. It has been proved that the estrogen receptor ligands possess
antiinflammatory activity with potential use in the treatment of inflammatory diseases.30 Thereby,
8
sclareol may act via the activation of the ligand-induced estrogen-like pathway to exhibit its
antiinflammatory effect.
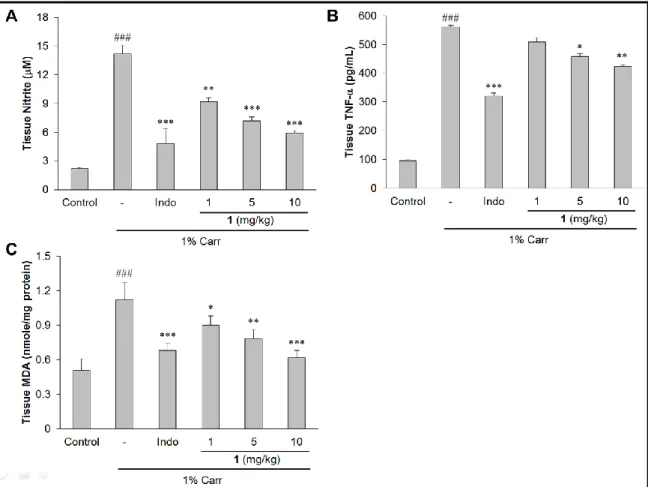
Inhibitory Effect of Sclareol (1) on NO, TNF-α and MDA within Carr-Induced Edema Paw. In Figure 4A, the NO level increased significantly in the edema serum at the 5 h after Carr
injection (p < 0.001), which can be markedly reversed by 1 as concentration more than 1 mg/kg
(p < 0.01). Likewise, both TNF-α and MDA level were increased significantly in the edema paw
at the 5 h after Carr injection (p < 0.001), and this effect was decreased significantly by treatment
with 1 as well as 10 mg/kg Indo (Figures 4B and 4C). The Carr test is highly sensitive to
non-steroidal antiinflammatory drugs, and has long been accepted as a useful phlogistic tool for
investigating new drug therapies.31 It is well known that the third phase of the edema-induced by
Carr, in which the edema reaches its highest volume, is characterized by the presence of
prostaglandins and other compounds of slow reaction found that the injection of Carr into the rat
paw induces the liberation of bradykinin, which later induces the biosynthesis of prostaglandin
and other autacoids, which are responsible for the formation of the inflammatory exudates.32 In
the present study, statistical analysis revealed that 10 mg/kg of Indo and 10 mg/kg of 1
significantly inhibited the development of edema 3 h after treatment (p < 0.001 or p < 0.01)
(Figure 3). L-arginine–NO pathway has been proposed to play an important role in the
Carr-induced inflammatory response,33 and the expression of the inducible isoform of NO
9
synthase has been proposed as an important mediator of inflammation.34 The results confirm that
Carr-induced paw edema model results in the production of NO, and the level of NO was
decreased significantly by treatment with 1, 5, and 10 mg/kg of 1 (Figure 4A). These results
suggest the antiinflammatory mechanism of 1 may be through the L-arginine−NO pathway
because 1 significantly inhibits the NO production. TNF-α is also a mediator of Carr-induced
inflammatory incapacitation, and is able to induce the further release of kinins and leukotrienes,
which is suggested to have an important role in the maintenance of long-lasting nociceptive
response.35 In this study, the results showed that 1 obviously decreased the level of serum TNF-α
after Carr injection by treatment with 5 and 10 mg/kg of 1 (Figure 4B).
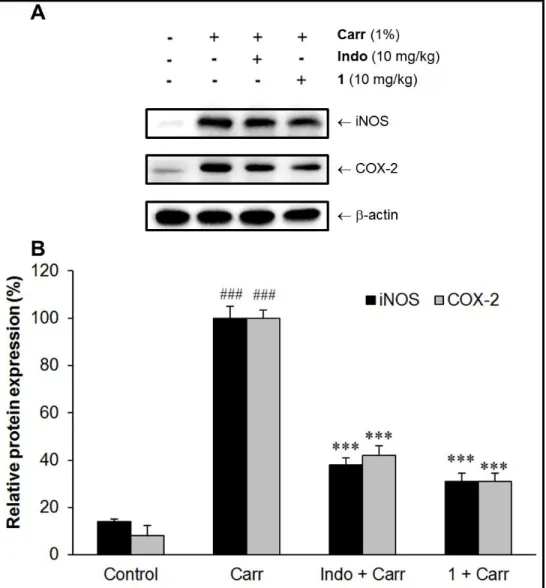
Inhibitory Effect of Sclareol (1) on iNOS and COX-2 within Carr-Induced Edema Paw.
Experimental results showed that 1 (10 mg/kg) can obviously inhibit (p < 0.001) both iNOS and
COX-2 proteins expression in edema paw as compared to Carr-treated alone group (Figure 5).
The experiments showed an average of 69.6 % and 70.3 % down-regulation of iNOS and COX-2
protein, respectively, after treatment with 1 at 10 mg/kg compared with the Carr-induced alone
(Figure 5B). In addition, the protein expression showed an average of 61.5 % and 58.1 %
down-regulation of iNOS and COX-2 protein after treatment with Indo at 10 mg/kg compared
with the Carr-induced alone (Figure 5B). The potency of 1 (10 mg/kg) on down-regulating the
expression of iNOS and COX-2 proteins was similar to that of Indo (10 mg/kg).
10
Inhibitory Effect of Sclareol (1) on neutrophils infiltration within Carr-Induced Edema
Paw. In Figure S2 (Supporting Information), neutrophils increased with Carr treatment (p <
0.001). As Indo and 1 (10 mg/kg) could significantly decrease the neutrophils numbers as
compared to the Carr-treated group (p < 0.001). The Carr-induced inflammatory response has
been linked to neutrophils infiltration and the production of neutrophils-derived free radicals as
well as the release of other neutrophils-derived mediators.35 It has been demonstrate that free
radical and NO will be released when administrating with Carr, and increasing free radical might
attack plasma membrane and result in the accumulation of MDA. The present study demonstrated
that 10 mg/kg of 1 the same as Indo can markedly decrease neutrophils infiltration and
accumulation of MDA within edema paw after Carr treatment (Figures S2 and 5C).
In Vivo Regulatory Effect of Sclareol (1) on the Activity of Antioxidative Enzymes. At 5h
after the intrapaw injection of Carr, liver tissues were analyzed for the biochemical parameters
such as CAT, SOD, and GPx activities (Table 1). CAT, SOD, and GPx activities in liver tissue
were decreased significantly by Carr administration, which were increased significantly after
treated with 5 mg/kg of 1 (p > 0.05 or p < 0.01) and 10 mg/kg Indo (p < 0.01). Glutathione (GSH)
plays an important role against Carr-induced local inflammation,36 and endogenous GSH can
reduce MDA production. In the present study, increases of CAT, SOD, and GPx activities were
found in the group with 1 treatment (Figure 4C and Table 1). Thereby, the suppression of MDA
11
production is probably due to the increases of CAT, SOD, and GPx activities. Besides, it has been
also demonstrated that sclareol (1) possess the antioxidant activity in vitro,37 whose effect may be
an alternative explanation for the reduced MDA found in hepatic tissue.
Conclusions. These results suggested that the antiinflammatory mechanism of 1 may be
related to the inhibitions of iNOS and COX-2, and it is associated with the increase in the
activities of antioxidant enzymes (CAT, SOD, and GPx). Based on reported bioactivities above, it
might be partially explained why 1 can exhibit the antiinflammatory effect in the LPS-stimulated
macrophages and the Carr-induced paw edema model. Compound 1 may be used as a
pharmacological agent or hormone-like ligand in the prevention or treatment of inflammatory
diseases.
12
Figure 1. Sclareol (1) reduced LPS-induced NO production in macrophages. ### p < 0.001
compared with control group (normal saline treated only). * p < 0.05, ** p < 0.01 and *** p <
0.001 compared to LPS treated alone group.
13
Figure 2. Sclareol (1) inhibits LPS-stimulated expression of inflammation-associated proteins in macrophages. (A) Western blot showing expression of iNOS and COX-2 proteins. The bar chart
indicated the relative protein expression of iNOS and COX-2 in macrophages after treatments (B).
### p < 0.001 compared with control group (normal saline treated only). ** p < 0.01 and *** p <
0.001 compared with LPS treated only group.
14
Figure 3. Sclareol (1) reduced mice hind paw edema induced by -carrageenan (Carr). ** p <
0.01 and *** p < 0.001 compared with the Carr group. Carr, 1% -carrageenan; Indo, 10 mg/kg indomethacin.
15
Figure 4. Sclareol (1) inhibits tissue content of NO (A), TNF- (B) and MDA (C) in
Carr-induced paw edema model. ### p <0.001 compared with control group (normal saline
treated only). * p < 0.05, ** p < 0.01 and *** p < 0.001 compared with the Carr group. Indo, 10
mg/kg indomethacin.
16
Figure 5. Sclareol (1) reduced expression of inflammation-associated proteins in edema paw induced by Carr. (A) Western blot showing protein expression of iNOS and COX-2. The bar chart
indicated the relative protein expression of iNOS and COX-2 within the paw tissue after
treatments (B). ### p < 0.001 compared with control group (normal saline treated only). *** p <
0.001 compared with Carr treated only group. Carr, 1% -carrageenan; Indo, 10 mg/kg indomethacin.
17
Table 1. Regulation of sclareol (1) on the enzyme activities of catalase, superoxide dismutase and
glutathione peroxidase in mouse liver.
Groups Catalase Glutathione
peroxidase
Superoxide dismutase
Control 5.3 ± 0.2 24.4 ± 0.3 3.4 ± 0.1
Carr 3.6 ± 0.3### 15.6 ± 0.3### 1.9 ± 0.2###
Indo + Carr 4.7 ± 0.2** 22.5± 0.6** 2.8 ± 0.2**
1 (1 mg/kg) + Carr 3.8 ± 0.3* 17.2 ± 0.5* 2.0 ± 0.3
1 (5 mg/kg) + Carr 4.4 ± 0.2** 19.8 ± 0.4** 2.6 ± 0.4*
1 (10 mg/kg) + Carr 5.0 ± 0.5*** 22.9 ± 0.6** 2.8 ± 0.2**
Activity unit of enzymes is presented as U/mg protein. Carr, 1% -carrageenan; Indo, 10 mg/kg indomethacin. ### p < 0.001 compared with the control group. * p < 0.05, ** p < 0.01 and *** p
< 0.001 compared with the Carr group.
EXPERIMENTAL SECTION
Chemicals. Sclareol (1), lipopolysaccharide (LPS) from Escherichia coli (serotype 0127:B8), carrageenan (Carr), indomethacin (Indo) and other chemicals were purchased from
Sigma-Aldrich (St. Louis, MO, USA). TNF-α was purchased from Biosource International Inc
(Camarillo, CA, USA). Anti-iNOS, anti-COX-2, anti-β-actin antibody (Santa Cruz Biotechnology,
18
CA, USA) and a protein assay kit (Bio-Rad Lab, Watford, Herts, UK) were obtained as indicated.
Polyvinylidene fluoride (PVDF) membrane (Immobilon-P® ) was obtained from Millipore Corp
(Bedford, MA, USA).
Cell Culture. A murine macrophage cell line RAW264.7 (BCRC No. 60001) was purchased from the Bioresources Collection and Research Center (BCRC) of the Food Industry Research
and Development Institute (Hsinchu, Taiwan). Cells were cultured in culture dishes containing
Dulbecco's Modified Eagle Medium (DMEM; Sigma-Aldrich) supplemented with 10% fetal
bovine serum (FBS; Sigma-Aldrich) in a CO2 incubator (5% CO2 in air) at 37°C and subcultured
every 3 days at a dilution of 1:5 using 0.05% trypsin–0.02% EDTA in Dulbecco's
phosphate-buffered saline (DPBS) without Ca2+ and Mg2+ ions.
Mice Model of Carr-Induced Paw Edema. Twenty-four male ICR mice were obtained from the BioLASCO Taiwan Co., Ltd. The animals housed in Plexiglas cages with free access to food
and water, and maintained at a constant temperature of 22 ± 1°C and relative humidity of 55 ± 5
% with a photocycle of 12-h light/dark. The experimental procedures were performed according
to the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals. In
addition, all tests were conducted under the guidelines of the International Association for the
Study of Pain.38 After a 2-week adaptation period, the mice (about 18-25 g) were randomly
assigned to four groups (n = 6) for further experiments. The control group receives normal saline,
19
and the other three groups include a Carr alone, Carr + Indo (a positive control), and 1
administered groups (1+ Carr). The Carr-induced hind paw edema model was used for
determination of antiinflammatory activity.39 Animals were treated with normal saline, Indo or 1
(1, 5 and 10 mg/kg) with intraperitoneal injection, 30 min prior to injection of 1% Carr (50 μL) in
the plantar side of right hind paws of the mice. The paw volume was measured immediately after
Carr injection and at 1, 2, 3, 4, and 5 h intervals after the administration of the edematogenic
agent using a Plethysmometer (model 7159; Ugo Basile, Varese, Italy). The degree of swelling
induced was evaluated by the ratio A/B, where A is the volume of the right hind paw after Carr
treatment, and B is the volume of the right hind paw before Carr treatment. Finally, the animals
were sacrificed and all of right hind paw were dissected and stored at -80 ºC. Also, blood were
withdrawn and kept at -80 ºC.
MTT Cell Viability Assay. Macrophages (2 × 105) were cultured in 96-well plate containing DMEM supplemented with 10% FBS for 1 day to become nearly confluent. Then cells were pre-treated with several concentrations (0, 1, 5 and 10 g/mL) of 1 for 1 h and then co-stimulated with 100 ng/mL of LPS for 24 h. After that, the cells were washed twice with DPBS and
incubated with 100 L of 0.5 mg/mL MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-
diphenyltetrazolium bromide) for 2 h at 37°C, and then the medium was discarded and 100 L of dimethyl sulfoxide (DMSO) was added. After 30 min incubation, absorbance at 570 nm was read
20
using a microplate reader.
Measurement of Nitric Oxide/Nitrite. NO production was indirectly assessed by measuring the nitrite levels in the cultured media and serum determined according to previous study.39 The
cells were pre-incubated with 1 (0, 1, 5 and 10 g/mL) for 1 hr and then co-treated with 100 ng/mL LPS at 37°C for 24 h. Subsequently, 100 L of each collected culture medium was mixed with the same volume of Griess reagent (1% sulfanilamide, 0.1% naphthyl ethylenediamine
dihydrochloride and 5% phosphoric acid) and incubated at room temperature for 10 min. The
absorbance of mixture was measured at 540 nm with a Micro-Reader (Molecular Devices,
Orleans Drive, Sunnyvale, CA). Homogenized tissue samples were diluted four times with
distilled water and deproteinized by adding 1/20 volume of zinc sulfate (300 mg/mL) to a final
concentration of 15 mg/mL. After centrifugation at 10,000 ×g for 5 min at room temperature, 100 μL of supernatant was applied into a microtiter plate, followed by 100 μL of Griess reagent. After 10 min of color development at room temperature, the absorbance was measured at 540 nm with
a Micro-Reader. By using sodium nitrite to generate a standard curve, the concentration of nitrite
was measured by absorbance at 540 nm.
Western Blot Analysis. The stimulated macrophages were washed with PBS and lysed in an ice-cold lysis buffer [10% glycerol, 1% Triton X-100, 1mM sodium orthovanadate, 1mM EGTA,
10mM sodium fluoride, 1mM sodium pyrophosphate, 20 mM Tris buffer (pH 7.9), 100 mM
21
-glycerophosphate, 137 mM sodium chloride, 5 mM EDTA and one protease inhibitor cocktail tablet (Roche, Indianapolis, IN, USA)] on ice for 1 h, followed by centrifugation at 12,000 ×g for
30 min at 4°C. Soft tissues were removed from individual mice paws and homogenized in a
solution containing 10 mM CHAPS, 1 mM phenylmethylsulphonyl fluoride (PMSF), 5 g/mL, aprotinin, 1 M pepstatin and 10 M leupeptin. The homogenates were centrifuged at 12,000 ×g for 20 min, and the supernatant was collected for Western blot analysis. Protein concentration
was measured by the Bio-Rad protein assay kit with bovine serum albumin as a standard. About 30 g of protein from the supernatants was then separated on 10% sodium dodecylsulphate- polyacrylamide gel (SDS-PAGE) and transferred to PVDF membranes. After transfer, the
membrane was blocked for 2 h at room temperature with 5% skim milk in TBST buffer (20 mM
Tris, 500 mM NaCl, pH 7.5 and 0.1% Tween 20). The membranes were then incubated with
mouse monoclonal anti-iNOS or anti-COX-2 antibody in 5% skim milk in TBST buffer for 2 h at
room temperature. The membranes were washed three times with TBST at room temperature and
then incubated with a 1:2000 dilution of anti-mouse IgG secondary antibody conjugated to
horseradish peroxidase (Sigma-Aldrich) in 2.5% skim milk in TBST for 1 h at room temperature.
The membranes were washed three times and the immunoreactive proteins were detected by
enhanced chemiluminescence (ECL) using Hyperfilm® ECL reagent (Amersham International,
Buckinghamshire, UK). The results of Western blot analysis were quantified by measuring the
22
relative intensity compared to the control using Kodak Molecular Imaging Software Ver.4.0.5
(Eastman Kodak Company, Rochester, NY, USA) and represented in the relative intensities. The results for iNOS and COX-2 were normalized to the band density of internal control (-actin), and the relative proteins expression were calculated according to the values of LPS treated alone
group as 100%.
Malondialdehyde Assay. Malondialdehyde (MDA) from Carr-induced edema foot was
evaluated by the thiobarbituric acid reacting substance (TRARS) method.39 Briefly, MDA reacted
with thiobarbituric acid in the acidic high temperature and formed a red-complex TBARS. The
absorbance of TBARS was determined at 532 nm.
Measurement of Serum TNF-. Serum levels of TNF- were determined using a
commercially available ELISA kit (Biosource International Inc., Camarillo, CA) according to the manufacturer's instruction. The concentration of serum TNF- was presented as pg/mL and determined according to the regression equation of the standard curve.
Measurement of Antioxidant Enzymes Activity. The following biochemical parameters were analyzed to check the hepatoprotective activity of 1 by the methods given below. Total
superoxidase dismutase (SOD) activity was determined by the inhibition of cytochrome c
reduction.40 The reduction of cytochrome c was mediated by superoxide anions generated by the
xanthine/xanthine oxidase system and monitored at 550 nm. One unit of SOD was defined as the
23
amount of enzyme required to inhibit the rate of cytochrome c reduction by 50%. Total catalase
(CAT) activity was measured according to previous study.41 In brief, the reduction of 10 mM
hydrogen peroxide in 20 mM of phosphate buffer (pH 7.0) was monitored by measuring the
absorbance at 240 nm. The activity was calculated using a molar absorption coefficient, and the
enzyme activity was defined as nanomoles of dissipating hydrogen peroxide per milligram
protein per minute. Total glutathione peroxidase (GPx) activity in cytosol was determined
according to Paglia and Valentine's method.42 The enzyme solution was added to a mixture
containing hydrogen peroxide and glutathione in 0.1 mM Tris buffer (pH 7.2) and the absorbance
at 340 nm was measured. Activity was evaluated from a calibration curve, and the enzyme
activity was defined as nanomoles of NADPH oxidized per milligram protein per minute.
Statistical Analysis. Data are expressed as mean ± standard error of the mean (SEM).
Statistical evaluation was carried out by one-way analysis of variance (ANOVA, Scheffe's
post-hoc test). A value of p < 0.05 was regarded as being statistically significant.
ACKNOWLEDGEMENTS
The authors want to thank the financial supports from the National Science Council (NSC
97-2313-B-039-001-MY3), China Medical University (CMU) (CMU99-S-29, CCM-P99-RD-042,
and CMU99-COL-10) and Taiwan Department of Heath Clinical Trial and Research Center of
24
Excellence (DOH100-TD-B-111-004).
SUPPORTING INFORMATION
The experimental procedure and pathological examination regarding the effect of sclareol (1) on
neutrophil infiltration within Carr-induced edema paw were provided online.
REFERENCES AND NOTES
(1) Weninger, S. C.; Yankner, B. A. Nat. Med. 2001, 7, 527-528.
(2) Bousquet, J.; Jeffery, P. K.; Busse, W. W.; Johnson, M.; Vignola, A. M. Am. J. Respir. Crit.
Care. Med. 2000, 161, 1720-1745.
(3) Libby, P. Nature 2002, 420, 868-874.
(4) Flavell, R. A. Curr Top Microbiol. Immunol. 2002, 266, 1-9.
(5) Rajput, S.; Wilber, A. Front. Biosci. (Schol Ed) 2010, 2, 176-183.
(6) Christodoulou, C.; Choy, E. H. Clin. Exp. Med. 2006, 6, 13-19.
(7) Liu, H.; Pope, R. M. Rheum. Dis. Clin. North Am. 2004, 30, 19-39, v.
(8) Wynn, T. A.; Barron, L. Semin. Liver Dis. 2010, 30, 245-257.
(9) Suschek, C. V.; Schnorr, O.; Kolb-Bachofen, V. Curr. Mol. Med. 2004, 4, 763-775.
(10) Crofford, L. J. J. Rheumatol. Suppl. 1997, 49, 15-19.
25
(11) Bradley, J. R. J. Pathol. 2008, 214, 149-160.
(12) Knowles, R. G.; Moncada, S. Biochem. J. 1994, 298 ( Pt 2), 249-258.
(13) James, S. L. Microbiol. Rev. 1995, 59, 533-547.
(14) Antoniou, K.; Malamas, M.; Drosos, A. A. Expert Opin. Pharmacother. 2007, 8, 1719-1732.
(15) Dimas, K.; Kokkinopoulos, D.; Demetzos, C.; Vaos, B.; Marselos, M.; Malamas, M.;
Tzavaras, T. Leukemia Res. 1999, 23, 217-234.
(16) Dimas, K.; Hatziantoniou, S.; Tseleni, S.; Khan, H.; Georgopoulos, A.; Alevizopoulos, K.;
Wyche, J. H.; Pantazis, P.; Demetzos, C. Apoptosis 2007, 12, 685-694.
(17) Dimas, K.; Papadaki, M.; Tsimplouli, C.; Hatziantoniou, S.; Alevizopoulos, K.; Pantazis, P.;
Demetzos, C. Biomed. Pharmacother. 2006, 60, 127-133.
(18) Hatziantoniou, S.; Dimas, K.; Georgopoulos, A.; Sotiriadou, N.; Demetzos, C. Pharmacol.
Res. 2006, 53, 80-87.
(19) Noori, S.; Hassan, Z. M.; Mohammadi, M.; Habibi, Z.; Sohrabi, N.; Bayanolhagh, S.
Cellular Immunol. 2010, 263, 148-153.
(20) Chan, E. D.; Riches, D. W. Am. J. Physiol.Cell Physiol. 2001, 280, C441-450.
(21) Roach, T. I.; Barton, C. H.; Chatterjee, D.; Liew, F. Y.; Blackwell, J. M. Immunology 1995,
85, 106-113.
(22) Teng, X.; Zhang, H.; Snead, C.; Catravas, J. D. Am. J. Physiol.Cell Physiol. 2002, 282,
26
C144-152.
(23) Arany, I.; Brysk, M. M.; Brysk, H.; Tyring, S. K. Cancer letters 1996, 110, 93-96.
(24) Stefano, G. B.; Salzet, M.; Magazine, H. I.; Bilfinger, T. V. J. Cardiovasc. Pharm. 1998, 31,
813-820.
(25) Deng, W. G.; Montero, A. J.; Wu, K. K. Arterioscl. Throm. Vas. 2007, 27, 1752-1759.
(26) Coccia, E. M.; Stellacci, E.; Marziali, G.; Weiss, G.; Battistini, A. Int. Immunol. 2000, 12,
977-985.
(27) Cui, X.; Yang, S. C.; Sharma, S.; Heuze-Vourc'h, N.; Dubinett, S. M. Biochem. Bioph. Res.
Co. 2006, 343, 995-1001.
(28) Cho, W.; Kim, Y.; Jeoung, D. I.; Kim, Y. M.; Choe, J. Mol. Immunol. 2011, 48, 966-972.
(29) Guo, F. H.; Uetani, K.; Haque, S. J.; Williams, B. R.; Dweik, R. A.; Thunnissen, F. B.;
Calhoun, W.; Erzurum, S. C. J. Clin. Invest. 1997, 100, 829-838.
(30) Steffan, R. J.; Matelan, E.; Ashwell, M. A.; Moore, W. J.; Solvibile, W. R.; Trybulski, E.;
Chadwick, C. C.; Chippari, S., Kenney, T.; Winneker, R.C.; Eckert, A.; Borges-Marcucci, L.;
Adelman, S. J.; Xu, Z.; Mosyak, L.; Harnish, D. C. Curr. Top. Med. Chem. 2006, 6, 103-111.
(31) Spector, W. G.; Willoughby, D. A. Bacteriol. Rev. 1963, 27, 117-154.
(32) Tohda, C.; Nakayama, N.; Hatanaka, F.; Komatsu, K. Evid. Based Complement Alternat.
Med. 2006, 3, 255-260.
27
(33) Cuzzocrea, S.; Zingarelli, B.; Calapai, G.; Nava, F.; Caputi, A. P. Life Sci. 1997, 60, 215-220.
(34) Pan, M. H.; Lai, C. S.; Dushenkov, S.; Ho, C. T. J. Agr. Food Chem. 2009, 57, 4467-4477.
(35) Dawson, J.; Sedgwick, A. D.; Edwards, J. C.; Lees, P. Int. J. Tiss. Teact. 1991, 13, 171-185.
(36) Chaturvedi, P. Evid. Based Complement Alternat. Med. 2008, 5, 55-59.
(37) Kolak, U.; Hacibekirŏglu, I.; Öztürk, M.; Özgökçe, F.; Topçu, G.; Ulubelen, A. Turk. J.
Chem. 2009, 33, 813-823.
(38) Zimmermann, M. Pain 1983, 16, 109-110.
(39) Chang, H. Y.; Sheu, M. J.; Yang, C. H.; Lu, T. C.; Chang, Y. S.; Peng, W. H.; Huang, S. S.;
Huang, G. J. Evid. Based Complement Alternat. Med. 2009, (In press)
(40) Flohe, L.; Otting, F. Methods Enzymol. 1984, 105, 93-104.
(41) Aebi, H. Methods Enzymol. 1984, 105, 121-126.
(42) Paglia, D. E.; Valentine, W. N. J. Lab. Clin. Med. 1967, 70, 158-169.
28
Table of Contents Graphic