國立台灣大學生命科學院生態學與演化生物學研究所 碩士論文
Department of Ecology and Evolutionary Biology College of Life Science
National Taiwan University Master Thesis
尼泊爾埋葬蟲族群在兩島嶼上的溫度適應
Thermal adaptation in two island populations of burying beetles (Nicrophorus nepalensis Hope 1831)
范郁盟 Yu-Meng Fan
指導教授:沈聖峰 博士 Advisor : Sheng-Feng Shen, Ph.D.
中華民國 108 年 3 月 March 2019
I
摘要
全球暖化對生物造成全面性的衝擊,生物如何適應不同的氣候差異,產生不同的溫 度適應模式,是預測暖化如何影響生物續存的重要問題。過去探討生物如何適應不 同的氣候主要有兩個假說:改變溫度耐受範圍的氣候變異度假說以及改變最適合 溫度的最佳溫度假說。生存及繁殖決定了生物的續存,但意義卻不相同,其適應模 式也不可一概而論,而過去研究大多著重於生存方面的溫度適應,對於繁殖的溫度 適應探討甚少,但繁殖才是生物終其一生的目標。我們以廣泛分布於亞洲的尼泊爾 埋葬蟲為實驗物種,比較各溫度下不同族群繁殖表現的差異,以日本奄美族群及台 灣合歡山族群進行比較,我們發現生存指標-溫度耐受度符合氣候變異度假說的 預測,奄美族群耐受範圍大於合歡山族群。而繁殖方面兩族群對溫度的適應則是比 較複雜,在繁殖成功率上兩族群並無顯著的不同,適存度(Fitness)也沒有顯著差異,
但在埋葬行為方面合歡山族群成功埋葬屍體的比顯著高於奄美族群;而奄美族群 雖然埋葬屍體的比例較低,但埋葬屍體後的繁殖成功率則顯著較高。且奄美族群成 功埋葬屍體後的適存度都高於合歡山族群,顯示兩族群有不同的繁殖策略。本研究 結果顯示,溫度耐受度的適應比較簡單,溫度變異大的環境耐受度也要增加,符合 氣候變異度假說的預測,而繁殖不同階段的策略明顯不同,表示繁殖是相對複雜的 行為,從埋葬屍體到子代孵化,其中包含了不只一個行為表現,並且各自的策略並 不相同,這說明了要充分了解氣候對生物適應的影響,除了生存以外也必須仔細考 慮其他的生活史特徵與適應性成分。
關鍵字:溫度適應、表現曲線、生活使、繁殖、存活
ABSTRACT
Climate change has a wide range of influences on organisms. Understanding how organism adapt to thermal variation is critical to forecast the fate of species in the warming world. Most of the previous studies focused on species’ thermal tolerance, but not other important fitness components such as reproduction, varied across different thermal environments. Therefore, how the different fitness components were influenced by climate remains poorly understood. Here, we test two hypotheses about climate adaptation: climate variation hypothesis, and thermal optimum hypothesis, through investigating the thermal tolerance and breeding performance of two populations of burying beetles (Nicrophorus nepalensis Hope 1831) from different latitudes (Amami
island: 28°15'12.7"N, and Mt. Hehuan: 24°10'48.2"N). We found that the range of thermal tolerance of the Amami population was broader than that of the Hehuan population, supporting the climate variation hypothesis. However, beetles from the Amami population had better breeding performance than those from the Hehuan population after successfully preparing their breeding resources (i.e. burying carcasses) in every tested temperature. On the other hand, beetles from the Hehuan population had better burying performance than those from Amami population especially in high temperature, and the two populations had no difference in finally fitness at every tested temperature. Our
III
carcass burial, and larva hatching, is affected by more complicated mechanisms than that of adult physiological tolerance. Our findings also showed the necessity to comprehensively understand the adaptive models of different life history traits.
Key words: thermal adaptation, performance curve, life-history, reproduction, survival
目錄
目錄 ... IV
Chapter 1 前言 ... 1
Chapter 2 材料與方法 ... 3
2.1 研究物種 ... 3
2.2 野外密度調查 ... 3
2.3 室內實驗 ... 3
2.3.1 實驗室埋葬蟲族群建立 ... 4
2.3.2 溫度耐受度實驗 ... 4
2.3.3 繁殖表現 ... 5
2.4 資料分析 ... 6
2.4.1 不同族群的溫度耐受度 ... 6
2.4.2 不同族群繁殖表現隨溫度的變化 ... 6
Chapter 3 結果 ... 7
3.1 棲地環境與溫度耐受度的關係 ... 7
3.2 繁殖表現與溫度的關係 ... 7
3.2.1 適存度 ... 8
3.2.2 埋葬屍體成功率 ... 8
3.2.3 埋葬屍體後的繁殖成功率 ... 8
3.2.4 埋葬屍體後的適存度 ... 8
Chapter 4 討論 ... 9
Chapter 5 參考文獻 ... 11
V
圖目錄
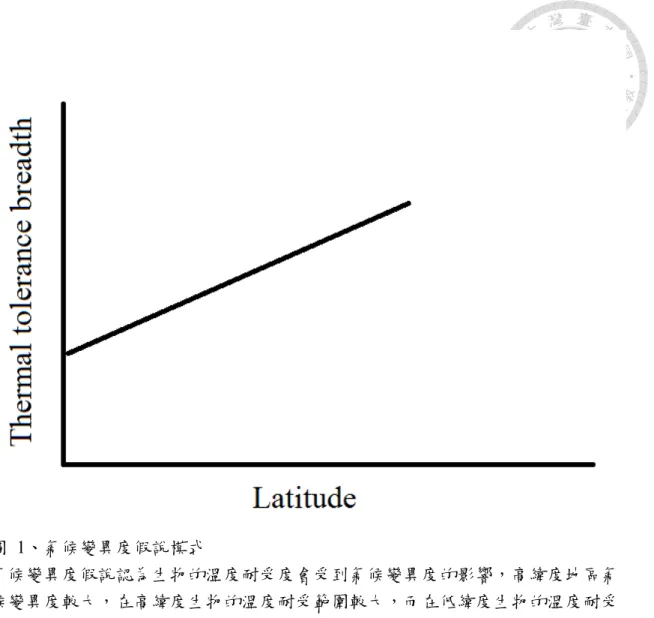
圖 1、氣候變異度假說模式 ... 14
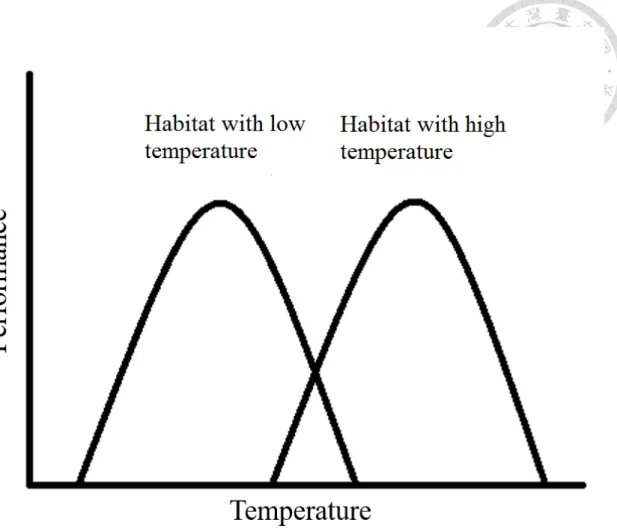
圖 2、最佳溫度假說模式 ... 15
圖 3、奄美族群與合歡山族群高溫耐受度差異 ... 16
圖 4、奄美族群與合歡山族群低溫耐受度差異 ... 17
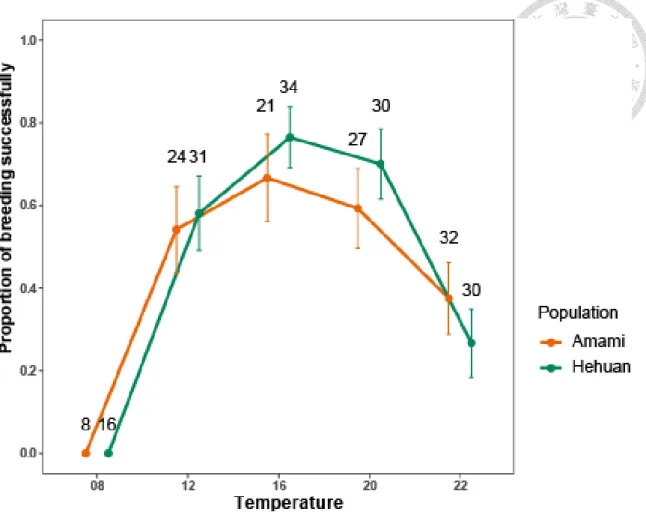
圖 5、各溫度下的繁殖成功率 ... 18
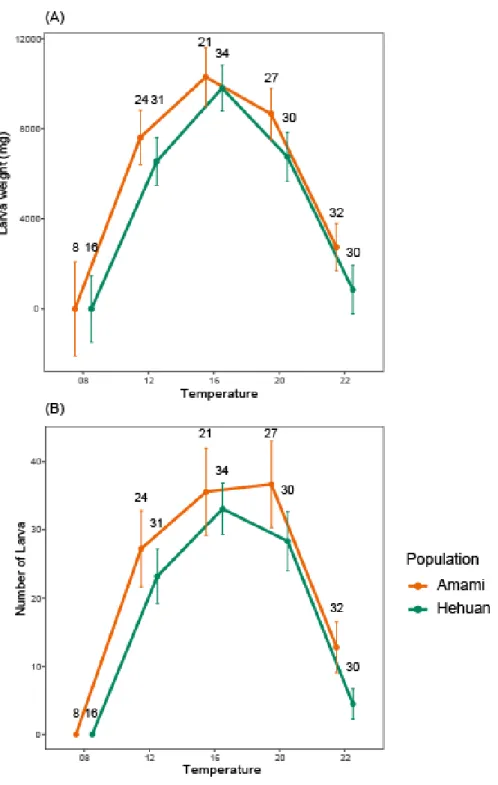
圖 6、各溫度下的適存度 ... 19
圖 7、各溫度下埋葬屍體的成功率 ... 20
圖 8、各溫度下埋葬屍體後的繁殖成功率 ... 21
圖 9、各溫度下埋葬屍體後的適存度 ... 22
表目錄
表 1、兩族群繁殖成功率隨溫度變化的變異數分析表(ANOVA table) ... 23
表 2、各溫度下的適存度的變異數分析表(ANOVA table) ... 24
表 3、各溫度下埋葬屍體成功率的變異數分析表(ANOVA table) ... 25
表 4、各溫度下埋葬屍體後繁殖成功率的變異數分析表(ANOVA table) ... 26
表 5、各溫度下埋葬屍體後適存度的變異數分析表(ANOVA table) ... 27
1
Chapter 1 前言
近年來人類活動造成全球暖化(Santer et al. 2003, Stott 2003, Barnett et al.
2005),從生理反應:植物開花發芽(Fitter and Fitter 2002)、動物繁殖遷徙 (Parmesan and Yohe 2003),到族群動態:分布改變(Root et al. 2003)、數量減少 (Crick et al. 1997, Thomas et al. 2004, Alan Pounds et al. 2006)都受到影響。研究 物種如何適應溫度的變化有助於預測暖化可能造成的影響,因此這個領域近 年逐漸備受重視(Merilä et al. 2000, Fields 2001, Karlsson and Van Dyck 2005, Forseth et al. 2009)。過去暖化研究大多以種間研究為主(Crick et al. 1997, Fitter and Fitter 2002, Parmesan and Yohe 2003, Thomas et al. 2004, Alan Pounds et al.
2006),少有探討不同族群受到的影響。但對於廣泛分布的物種來說,不同族 群可能會有不同的溫度適應,因此受到暖化的影響也會有所不同。另一方面,
大部分溫度適應的研究中,往往只探討某一特定生理表現隨溫度的變化,例如 生長(Drent 2002, Sun and Friedmann 2005)、發育(Berven et al. 1979)、存活(Merilä et al. 2000)及運動(Wilson 2001)等,其中又以存活被廣泛的討論。在探討生物 的續存時,除了生存之外,繁殖是另一個重要的生活史特徵。生物終其一生的 目標是傳遞自己的基因,但與繁殖有關的適應研究卻鮮少被關注。故本研究將 探討繁殖在不同島嶼上的適應機制,比較不同族群面對不同環境所演化出來 的適應模式。
過去研究生物在生理上的適應大部分以探討溫度適應為主,較為經典的就 是氣候變異度假說(Climate variability hypothesis, CVH)(Stevens 1989)。此假說 認為生物的溫度耐受度會受到氣候變異度的影響,高緯度地區氣候變異度大,
溫度耐受範圍也較寬(Addo-Bediako et al. 2000, Sunday et al. 2011)(參考圖 1)。
若生物繁殖的溫度適應也受到生理溫度耐受度的限制,則可以預測繁殖的溫
度適應也會受到氣候變異度的影響,在氣候變異度大的地區能繁殖的溫度範 圍也較大,但權衡(trade-off)的結果可能造成平均子代數較少。
氣候變異度假說大多是符合物種間差異的結果(Addo-Bediako et al. 2000, Sunday et al. 2011),若是進行族群間比較則大多會符合最佳溫度假說(Thermal optimum hypothesis)的模式(Ståhlberg et al. 2001, Wilson 2001, Castaneda 2004)。
最佳溫度假說認為溫度適應不是在耐受寬度上的適應,而是最適合溫度上的 適應。在較冷環境下最適合表現溫度較低,反之較高。根據此假說我們預測年 均溫較低的地區,族群耐受溫度較低溫,適合繁殖的溫度也較低,但溫度耐受 寬度及平均子代數則沒有差異(參考圖 2)。
本研究以尼泊爾埋葬蟲作為實驗物種,選用台灣合歡山族群以及日本奄美 大島族群進行實驗,由於尼泊爾埋葬蟲族群分布橫跨的緯度夠大,又屬於狹溫 性物種(黃文伯 2007),很適合用來探討不同族群的溫度適應。此外尼泊爾埋 葬蟲具有相當特殊的埋葬行為,會將小型屍體埋入土中,以避免其他競爭者使 用。屍體是尼泊爾埋葬蟲可用來繁殖的唯一資源,因此埋葬行為是直接關係到 能否繁殖成功的關鍵。以埋葬行為進行實驗,可以更好的反應出溫度對於埋葬 蟲在演化策略上的影響。我們將探討不同埋葬蟲族群如何適應不同環境,發展 出最適合的策略,更進一步了解不同族群溫度適應的演化機制。
3
Chapter 2 材料與方法
2.1 研究物種
本研究物種尼泊爾埋葬蟲(Nicrophorus nepalensis Hope 1831)是屬於埋葬蟲科覆 葬甲屬的一種食屍性昆蟲,屍體除了是他們的食物來源,更是用來養育後代的唯一 資源。他們會將找到的屍體去除毛髮及蛆,並以分泌物進行防腐,再將屍體摺疊成 球狀埋入土中形成巢室,並在巢室附近產卵,孵化後直接以屍球進行育幼,幼蟲達 三齡時則離開巢室準備化蛹,但屍體在野外是屬於高度珍稀的資源(Scott and Gladstein 1993),種間競爭非常強烈,因此埋葬行為就顯得非常重要。尼泊爾埋葬 蟲廣泛分布於亞洲地區,巴基斯坦、台灣、馬來西亞、日本等地(Bala and Singh 2017),
族群間橫跨的緯度巨大,卻又屬於狹溫性昆蟲,對溫度變化非常敏感(黃文伯 and 環境與生態學報 2011)。
綜合以上特徵,尼泊爾埋葬蟲同時具有對溫度變化敏感、族群間面對的氣候差異巨 大、特殊埋葬行為等特點,因此很適合用來探討氣候對生存及繁殖的影響。
2.2 野外密度調查
本研究分別於中部橫貫公路及奄美大島設置採樣點,一樣點設置一懸吊式誘捕 陷阱進行尼泊爾埋葬蟲的密度調查。將100 ± 10 克之豬絞肉放入 500 毫升、開口 直徑4.5 公分之塑膠瓶內,在瓶口處套上 24 × 24 公分之方形木板供埋葬蟲停棲,
瓶口上方10 公分處加上相同大小之木板作為遮雨裝置。整個陷阱懸吊於離地 1 公
尺以上的高度,避免脊椎動物取食。陷阱在野外經過4 個晚上後收回,並紀錄尼泊
爾埋葬蟲之數量、性別,紀錄結束後將埋葬蟲原地野放。陷阱旁會再架設溫度濕度 紀錄儀器,記錄環境資料,同樣在4 個晚上後收回。
中部橫貫公路調查時間為2014-2015 年,奄美大島則是 2015-2018 年
2.3 室內實驗
2.3.1 實驗室埋葬蟲族群建立
為了瞭解不同族群在溫度上是否有適應分化,我們將野外的尼泊爾埋葬蟲帶回 室內繁殖建立族群,再進行室內的操作實驗。
我們於2018 年 4 月將日本奄美大島的野外尼泊爾埋葬蟲帶回實驗室內飼養,將一 公一母隨機配對放入土深10 公分之中型飼養箱(20x13x13cm)中,再放入平均 75 g
± 10% 的老鼠屍體作為繁殖資源,再將飼養箱置於步入式生長箱。約十四天後,將 離巢之三齡幼蟲取出秤重並放入已填土的十四格盒中供其化蛹。十四格盒同樣至
於步入式生長箱,約45 天後取出羽化之成蟲,紀錄性別、背板寬、體重。之後再
放入320 毫升的塑膠方盒中飼養,每周餵食一次麥皮蟲,直至離巢日算起 75 天後 性成熟即可開始做實驗。此世代我們稱之為野生型(wildtype),接下來每一代則是 F1、F2…以此類推。為了避免野外因子可能造成的影響,所有的實驗都只以野生型 以及之後的世代進行實驗,以確保所有實驗用蟲都是在我們所控制的環境下成長。
上述步入式生長箱的溫溼設定是模擬合歡山地區尼泊爾埋葬蟲分布密度最高的環 境之溫濕。
2.3.2 溫度耐受度實驗
我們根據 Sheldon 及 Tewksbury(Sheldon and Tewksbury 2014)所提出的方法測量 臨界溫度(Critical temperature),得到溫度耐受度的資料。臨界溫度是指生物在此溫 度下會失去正常的反應(loss of the righting response),導致生物無法逃離該環境的 各種威脅,最終造成死亡,分別有臨界高溫及臨界低溫。
我們使用實驗室內的繁殖蟲進行實驗,為了避免年齡造成的影響,只使用離巢日 75 到 105 天的成蟲進行實驗。將水浴槽(Yihder Co., Ltd, BL-720D)設至定溫(CTmin
設置-10℃,CTmax設置50℃),實驗前一小時先將埋葬蟲預先移至培養箱外,並且 測量前胸背板寬(mm)及體重(mg),架設好熱像儀(FLIR Systems, Inc., SC300 Series),
準備進行拍攝。每次實驗皆放入一隻埋葬蟲至樂扣玻璃盒中,樂扣玻璃盒會先放置
5
一層厚約5mm 的培養土,以利於埋葬蟲穩定移動,並在玻璃盒內壁塗上凡士林,
避免埋葬蟲爬上內壁。開始實驗前先用熱像儀拍照記錄蟲體的前測體溫,再將埋葬 蟲放入樂扣玻璃盒後沉入水域槽中,淹沒至開口邊緣。過程中埋葬蟲體溫將會慢慢 與外界溫度達到平衡,並且逐漸失去行動能力,直至埋葬蟲無法繼續移動,用熱像 儀拍照紀錄,就可以得到休克時的體溫,即是Critical temperature,並快速將埋葬
蟲移回室溫之中,即會恢復正常活動能力,並且經過24 小時確定沒有死亡,才視
為可用資料。實驗過程中皆不會接觸蟲體,且同一隻埋葬蟲不會重複進行實驗,最 後再觀看熱像儀影像,手動圈選影像中蟲體的前胸背板寬,紀錄前胸背板均溫、最 高溫及最低溫。
每一族群一臨界溫度做30 重複。
2.3.3 繁殖表現
要了解生物性能表現隨溫度變化的反應,溫度表現曲線(Thermal performance curves)是一個很好的工具。該曲線描述了生物性能表現與溫度之間的關係(Huey and Kingsolver 1989, Kingsolver et al. 2004),透過該曲線可以得知生物性能表現隨 溫度變化的特徵:
1. 最佳溫度(Thermal optimum),性能表現最佳的溫度。
2. 最低臨界溫度(Critical temperature minimum, CTmin),具有性能表現的最低溫度,
低於此溫度則失去性能表現能力。
3. 最高臨界溫度(Critical temperature maximum, CTmax),具有性能表現的最高溫度,
高於此溫度則失去性能表現能力。
4. 表現寬度(Performance breadth),具有性能表現的溫度範圍,即是最高臨界溫度 到最低臨界溫度的溫度區間。
繁殖表現使用實驗室內的繁殖蟲進行,為了避免年齡對子代的影響,我們只選用離 巢日75 天到 105 天之埋葬蟲進行實驗。配對方式為兩巢的公母互相配對(例如:A 公 x B 母、A 母 x B 公)。將公母蟲測量完背板及秤重後放入土深 10 公分之中型
飼養箱(20x13x13cm)中,用平均 75 g ± 10% 的老鼠屍體作為繁殖資源,再將飼養 箱放入不同環境溫度中(8、12、16、20、22℃),紀錄屍體埋入土中之天數。我們根 據野外資料取得各溫度下蛆消耗屍體的天數,只要埋葬蟲超過該天數仍沒將屍體 埋入土中則視為繁殖失敗,分別為8、12℃:6 天,16℃:3 天,20、22℃:2 天。
當屍體埋入土中則開始計算離巢日,當三齡幼蟲離巢時紀錄數量及重量,並記錄公 母蟲存活狀況後冰入-80℃冰箱。
繁殖表現方面我們統計了繁殖成功率、埋葬屍體比例以及適存度(Fitness)。繁殖成 功率我們以有無三齡幼蟲離巢來定義繁殖成功與否。適存度是以族群為單位,所以 該族群無論繁殖成功與否都計算在內。
每一族群一個溫度處理做30 重複。
2.4 資料分析
本研究所有分析皆使用統計軟體 R 3.3.2 進行,並以 95%信賴區間為顯著標準,
當p-value 小於 0.05 則視為統計顯著。
2.4.1 不同族群的溫度耐受度
為檢驗兩族群間的CTmax 及 CTmin 是否有差異,我們以 T 檢定分別對 CTmax 及CTmin 進行兩族群的比較。
2.4.2 不同族群繁殖表現隨溫度的變化
所有的分析皆是以地區及溫度以及兩者的交互作用當作變數進行雙因子變異數 分析(two way ANOVA)。若族群間有顯著差異則再進行事後的成對比較,找出兩族 群有差異的溫度處理,成對比較是使用Tukey Test。
7
Chapter 3 結果
3.1 棲地環境與溫度耐受度的關係
為了預測最佳溫度假說與氣候變異度假說,我們從CRU(Climatic Research Unit) 取得了 1901 年到 2016 年合歡山地區及奄美地區的溫度資料,計算兩地區的年溫 差及年均溫。結果顯示奄美地區的年溫差為14.97 度,合歡山地區則是 10.14 度,
有顯著差異(t-test, t = -37.15, p < 0.001, n = 232)。奄美地區的年均溫為 20.01 度,合 歡山為10.33 度,兩地均溫也有顯著差異(t-test, t = -58.09, p < 0.001, n = 2784)。此 外年均溫方面根據蔡祥瑜(2017)的研究,我們計算兩族群各自適合繁殖的月份均溫。
合歡山族群的繁殖季為夏季(長日照,日照>12 小時),均溫為 13 度。奄美族群繁殖 季則是冬季(短日照,日照<12 小時),均溫為 16.17 度。根據繁殖季的均溫,奄美 地區顯著高於合歡山地區(t-test, t = 20.71, p < 0.05)。除了根據日照定義出繁殖季之 外,我們還根據野外的實驗記錄,得到埋葬蟲有繁殖成功紀錄的月份去計算均溫。
根據野外資料合歡山地區全年都有繁殖成功的紀錄,均溫為10.33 度。奄美地區有 繁殖成功紀錄地月份僅有12 月及 1 月,均溫為 13.88 度。根據實際繁殖成功的均 溫,奄美地區顯著高於合歡山地區(t-test, t = 26.35, p < 0.05)。綜合以上結果,奄美 地區的均溫都顯著高於合歡山地區。因此根據最佳溫度假說的預測,奄美族群臨界 高溫比合歡山族群高,臨界低溫也比合歡山族群高,並且溫度耐受寬度沒有差異。
而氣候變異度假說的預測則是奄美族群溫度耐受度大於合歡山族群。
我們分析兩族群的臨界溫度差異,結果顯示在奄美族群的臨界高溫顯著高於合 歡山族群(t-test, t = 2.937, p = 0.004, n = 117)(圖 3);奄美族群的臨界低溫卻顯著低 於合歡山族群(t-test, t = -2.354, p = 0.02, n = 108)(圖 4),臨界高溫較高,但臨界低溫 較低,造成奄美族群的溫度耐受範圍比合歡山族群的溫度耐受範圍大,支持氣候變 異度假說的預測。
3.2 繁殖表現與溫度的關係
在繁殖成功率方面,兩族群繁殖成功率隨溫度變化的趨勢相同(ANOVA, region
× temperature, p = 0.728, 表格 1),並且兩族群在各溫度下的成功率無顯著差異 (ANOVA, p = 0.641,表格 1)(圖 5)。
3.2.1 適存度
我們以數量及重量當作適存度的指標。重量是以一對公母所產下的子代總重當 作指標而非平均重。結果發現族群間無論是數量或是總重都沒有顯著差異(ANOVA, p = 0.063, p = 0.097, 表格 2(A)、(B)),而且兩族群的子代數量及總重隨溫度變化的 趨勢相同(ANOVA, region × temperature, p = 0.904, p = 0.937, 表格 2(A)、(B)) (圖 6)。
總重的結果也是一樣(圖 7)。
3.2.2 埋葬屍體成功率
在埋葬屍體的成功率上,族群間隨溫度變化的趨勢相同(ANOVA, region × temperature, p = 0.617)。但兩族群成功埋葬屍體的比例卻有顯著的差異(ANOVA, p
< 0.001, 表格 3)。除了 8 度幾乎都無法埋葬屍體之外,高溫環境下合歡山族群成功 埋葬屍體的比例顯著高於奄美族群 (圖 7)。
3.2.3 埋葬屍體後的繁殖成功率
接著繼續檢視埋葬屍體後的繁殖成功率,結果發現兩族群埋葬屍體後的繁殖成 功率有顯著的不同(ANOVA, p < 0.001, 表格 4),雖然族群間隨溫度變化的趨勢相 同(ANOVA, region × temperature, p = 0.561, 表格 4),但成功埋葬屍體後奄美族群的 繁殖成功率遠高於合歡山族群 (圖 8)。
3.2.4 埋葬屍體後的適存度
埋葬屍體後的子代品質兩族群隨溫度變化的趨勢相同,(ANOVA, region × temperature, p = 0.39, p = 0.744, 表格 5(A)、(B)),但各溫度下奄美族群的子代數量 及總重都大於合歡山族群(ANOVA, p < 0.001 表格 5(A)、(B)) (圖 9)。
9
Chapter 4 討論
本研究藉由測試來自不同族群的尼泊爾埋葬蟲在不同溫度下的繁殖表現,檢測 溫度適應的兩個假說,以了解繁殖的適應模式,結果顯示繁殖的適應與過去研究生 存的適應模式並不相同。
溫度耐受度符合氣候變異度假說的預測,在年溫差大的奄美族群其溫度耐受範 圍也會較大,與過去的研究結果相同;繁殖表現若是從成功率、子代品質的角度分 析則會得出兩族群沒有溫度上分化適應的結果。然而若看埋葬屍體之後的結果,則 會發現奄美族群的成功率、子代品質皆高於合歡山族群,但就埋葬屍體的成功率而 言,則是合歡山族群高於奄美族群。上述結果顯示出了兩族群的策略有所不同。
生物的溫度耐受範圍實驗在過去已有不少研究,最為熱門的議題就屬溫度耐受 度與溫度變異的關係,在溫度變異大的地區容易選擇出溫度耐受度較大的生物,相 反 的 在 溫 度 變 異 小 的 地 區 容 易 使 生 物 的 溫 度 耐 受 度 演 化 成 較 專 一 的 特 化 型 (Sheldon and Tewksbury 2014),相關的研究也都證實溫度耐受度符合氣候變異度假 說的預測(Addo-Bediako et al. 2000, Sunday et al. 2011, Sheldon and Tewksbury 2014)。
溫度耐受度是受到溫差的影響,與本實驗的結果一致,並且本實驗結果也顯示了除 了物種間的適應模式之外,族群間的溫度耐受度適應模式也同樣適用氣候變異度 假說的預測,這在過去的研究是較少被關注的部分。此外過去的研究針對不同類群 有發展出不同的溫度耐受度測量值,但基本都是找出生物失去自發反應時的溫度 為概念。相對的,本實驗是以臨界溫度(Critical temperature)當作指標,在實驗效率 上比過去常用的致死溫度(Lethal temperature)來的高,並且實驗結果是一致的,說 明臨界溫度與致死溫度對生物的意義是類似的,在生態的角度也有其代表性。
繁殖的溫度適應研究則是相對比較少(Angilletta Jr and Angilletta 2009),近年來 較為完整的繁殖研究也只比較了不同棲地類型的影響(Karlsson and Van Dyck 2005),
並且只探討環境均溫,而繁殖表現則是以卵數為指標。但在本研究中發現即使有產 卵也不一定會有子代的孵化,若只考慮卵數有可能高估了繁殖的表現,而該研究的
結果顯示產卵表現是適應均溫,棲地均溫較高的族群在高溫的產卵數較多,反之亦 然。
本研究結果顯示,族群間的溫度適應有其差異存在,但過往氣候變遷的研究中 大多是在物種的層次進行(Parmesan and Yohe 2003, Root et al. 2003, Thomas et al.
2004),族群層次較少被考慮。但族群間的適應差異可能是該物種能否在變遷環境 下生存的關鍵。此外,生理耐受度與繁殖耐受度代表的生態意義並不相同,生理耐 受度代表能分布的範圍,但能否建立族群則是要看繁殖耐受度。而繁殖行為是相對
複雜的行為表現,尤其是在具有特殊繁殖行為(處理繁殖資源:例如埋葬屍體)的物
種中,該行為會間接影響繁殖能否成功。因此更需要仔細了解不同行為的適應模式,
不能僅僅只探討產下的子代品質,了解更多繁殖行為的適應模式,有助於我們更準 確的預測暖化對生物造成的影響。
11
Chapter 5 參考文獻
Addo-Bediako, A., S. L. Chown, and K. J. Gaston. 2000. Thermal tolerance, climatic variability and latitude. Proceedings of the Royal Society of London B:
Biological Sciences 267:739-745.
Alan Pounds, J., M. R. Bustamante, L. A. Coloma, J. A. Consuegra, M. P. L. Fogden, P.
N. Foster, E. La Marca, K. L. Masters, A. Merino-Viteri, R. Puschendorf, S. R.
Ron, G. A. Sánchez-Azofeifa, C. J. Still, and B. E. Young. 2006. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 439:161.
Angilletta Jr, M. J., and M. J. Angilletta. 2009. Thermal adaptation: a theoretical and empirical synthesis. Oxford University Press.
Bala, M., and N. Singh. 2017. Geographical distribution of some forensically important species of beetles (Coleoptera: silphidae) from North India (INDIA).
Barnett, T. P., D. W. Pierce, K. M. AchutaRao, P. J. Gleckler, B. D. Santer, J. M. Gregory, and W. M. Washington. 2005. Penetration of Human-Induced Warming into the World's Oceans. Science 309:284-287.
Berven, K. A., D. E. Gill, and S. J. Smith‐Gill. 1979. Countergradient selection in the green frog, Rana clamitans. Evolution 33:609-623.
Castaneda, L. 2004. Adaptive latitudinal shift in the thermal physiology of a terrestrial isopod.
Crick, H. Q. P., C. Dudley, D. E. Glue, and D. L. Thomson. 1997. UK birds are laying eggs earlier. Nature 388:526.
Drent, J. 2002. Temperature responses in larvae of Macoma balthica from a northerly and southerly population of the European distribution range. Journal of Experimental Marine Biology and Ecology 275:117-129.
Fields, P. A. 2001. Review: Protein function at thermal extremes: balancing stability and flexibility. Comparative Biochemistry and Physiology Part A: Molecular &
Integrative Physiology 129:417-431.
Fitter, A. H., and R. S. R. Fitter. 2002. Rapid Changes in Flowering Time in British Plants.
Science 296:1689-1691.
Forseth, T., S. Larsson, A. J. Jensen, B. Jonsson, I. Näslund, and I. Berglund. 2009.
Thermal growth performance of juvenile brown trout Salmo trutta: no support for thermal adaptation hypotheses. Journal of Fish Biology 74:133-149.
Huey, R. B., and J. G. Kingsolver. 1989. Evolution of thermal sensitivity of ectotherm performance. Trends in Ecology & Evolution 4:131-135.
Karlsson, B., and H. Van Dyck. 2005. Does habitat fragmentation affect temperature-
related life-history traits? A laboratory test with a woodland butterfly.
Proceedings of the Royal Society of London B: Biological Sciences 272:1257- 1263.
Kingsolver, J. G., G. J. Ragland, and J. G. Shlichta. 2004. Quantitative genetics of continuous reaction norms: thermal sensitivity of caterpillar growth rates.
Evolution 58:1521-1529.
Merilä, J., A. Laurila, A. T. Laugen, K. Räsänen, and M. Pahkala. 2000. Plasticity in age and size at metamorphosis in Rana temporaria - comparison of high and low latitude populations. Ecography 23:457-465.
Parmesan, C., and G. Yohe. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37.
Root, T. L., J. T. Price, K. R. Hall, S. H. Schneider, C. Rosenzweig, and J. A. Pounds.
2003. Fingerprints of global warming on wild animals and plants. Nature 421:57.
Santer, B. D., M. F. Wehner, T. Wigley, R. Sausen, G. Meehl, K. Taylor, C. Ammann, J.
Arblaster, W. Washington, and J. Boyle. 2003. Contributions of anthropogenic and natural forcing to recent tropopause height changes. science 301:479-483.
Scott, M. P., and D. S. J. E. E. Gladstein. 1993. Calculating males? An empirical and theoretical examination of the duration of paternal care in burying beetles.
7:362-378.
Sheldon, K. S., and J. J. Tewksbury. 2014. The impact of seasonality in temperature on thermal tolerance and elevational range size. Ecology 95:2134-2143.
Stevens, G. C. 1989. The latitudinal gradient in geographical range: how so many species coexist in the tropics. The American Naturalist 133:240-256.
Ståhlberg, F., M. Olsson, and T. Uller. 2001. Population divergence of developmental thermal optima in Swedish common frogs, Rana temporaria. Journal of Evolutionary Biology 14:755-762.
Stott, P. A. 2003. Attribution of regional‐scale temperature changes to anthropogenic and natural causes. Geophysical Research Letters 30.
Sun, H. J., and E. I. Friedmann. 2005. Communities adjust their temperature optima by shifting producer-to-consumer ratio, shown in lichens as models: II.
Experimental verification. Microbial ecology 49:528-535.
Sunday, J. M., A. E. Bates, and N. K. Dulvy. 2011. Global analysis of thermal tolerance and latitude in ectotherms. Proceedings of the Royal Society of London B:
Biological Sciences 278:1823-1830.
Thomas, C. D., A. Cameron, R. E. Green, M. Bakkenes, L. J. Beaumont, Y. C.
Collingham, B. F. N. Erasmus, M. F. de Siqueira, A. Grainger, L. Hannah, L.
Hughes, B. Huntley, A. S. van Jaarsveld, G. F. Midgley, L. Miles, M. A. Ortega-
13
Extinction risk from climate change. Nature 427:145.
Wilson, R. S. 2001. Geographic variation in thermal sensitivity of jumping performance in the frog <em>Limnodynastes peronii</em>. Journal of Experimental Biology 204:4227-4236.
黃文伯. 2007. 環境變遷監測-氣溫對狹溫性甲蟲活動之影響. 林業研究專訊 14:7- 10.
黃文伯, and 葛. J. 環境與生態學報. 2011. 哈盆自然保留區屍食性甲蟲物種生物多 樣性監測與氣候變遷之關係. 4:17-34.
蔡祥瑜. 2017. 尼泊爾埋葬蟲繁殖策略的地區適應演化. 臺灣大學生態學與演化生 物學研究所學位論文.
圖 1、氣候變異度假說模式
氣候變異度假說認為生物的溫度耐受度會受到氣候變異度的影響,高緯度地區氣 候變異度較大,在高緯度生物的溫度耐受範圍較大,而在低緯度生物的溫度耐受 範圍較小。
15
圖 2、最佳溫度假說模式
最佳溫度假說認為生物會適應不同的均溫,在較冷環境下最佳表現溫度較低,反之 較高,由於耐受溫度的寬度沒有改變,因此平均表現量沒有差異,僅僅是最適合溫 度上的平移。
圖 3、奄美族群與合歡山族群高溫耐受度差異
我們以臨界溫度為指標,比較不同族群的溫度耐受度,高溫部分的結果顯示奄美 族群的高溫耐受度顯著高於合歡山族群(t-test, t = 2.937, p = 0.004)。誤差線為標準 誤差,數字為樣本數。
17
圖 4、奄美族群與合歡山族群低溫耐受度差異
我們以臨界溫度為指標,比較不同族群的溫度耐受度,低溫部分的結果顯示奄美 族群的低溫耐受度顯著低於合歡山族群(t-test, t = -2.354, p = 0.02)。誤差線為標準 誤差,數字為樣本數。
圖 5、各溫度下的繁殖成功率
族群間繁殖成功率隨溫度變化的趨勢相同(ANOVA, region × temperature, p = 0.617),
且成功率無顯著差異(ANOVA, p = 0.641),僅在溫度間有差(χ2 = 59.087, p < 0.001),
極端溫度的繁殖成功率顯著降低。
點與誤差線分別代表平均值及標準誤,誤差線上的數字為樣本數。
19
圖 6、各溫度下的適存度
子代品質我們以重量及數量當作指標,(A)、(B)分別為一對公母所產下子代的總重 及數量,兩者呈現的結果相同,不論是總重或數量兩族群隨溫度變化的趨勢相同 (ANOVA, region × temperature, p = 0.937、p = 0.904),且兩族群子代品質無顯著差 異(ANOVA, p = 0.097, p = 0.063),僅在溫度間有差,極端溫度的子代品質顯著降 低。
點與誤差線分別代表平均值及標準誤,誤差線上的數字為樣本數。
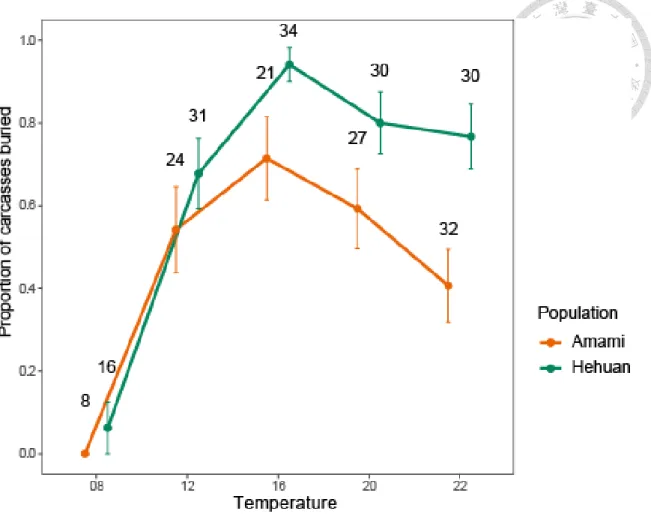
圖 7、各溫度下埋葬屍體的成功率
兩族群埋葬屍體的比例隨溫度變化的趨勢相同(ANOVA, region × temperature, p = 0.617),但族群間埋葬屍體的成功率有顯著差異(ANOVA, p < 0.001),合歡山族 群埋葬屍體的比例顯著高於奄美族群,且22 度埋葬屍體的成功率顯著下降。
點與誤差線分別代表平均值及標準誤,誤差線上的數字為樣本數。
21
圖 8、各溫度下埋葬屍體後的繁殖成功率
兩族群在埋葬屍體後的繁殖成功率隨溫度變化的趨勢相同(ANOVA, region × temperature, p = 0.561),但族群間的繁殖成功率有顯著差異(ANOVA, p < 0.001),
合歡山族群埋葬屍體後的繁殖成功率顯著低於奄美族群,並且合歡山族群22 度的
繁殖成功率顯著降低。
點與誤差線分別代表平均值及標準誤,誤差線上的數字為樣本數。
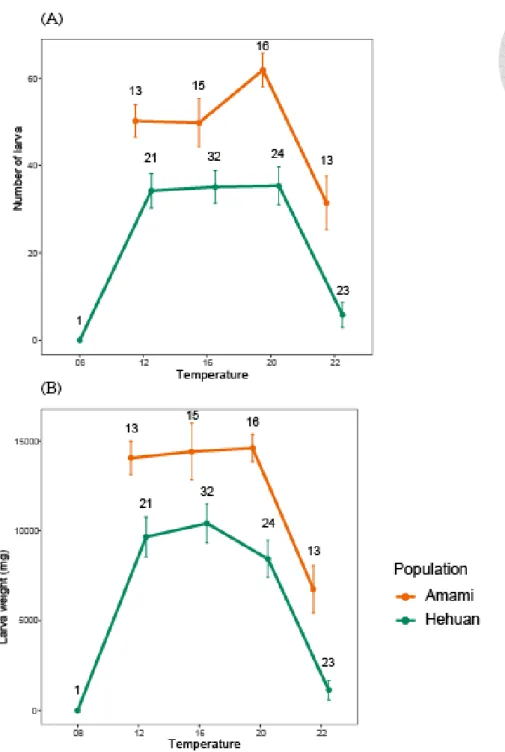
圖 9、各溫度下埋葬屍體後的適存度
(A)、(B)分別為一對公母所產下子代的數量及總重,兩者呈現的結果相同,在埋葬 屍 體 後 不 論 是 總 重 或 數 量 兩 族 群 隨 溫 度 變 化 的 趨 勢 相 同(ANOVA, region × temperature, p = 0.744、p = 0.39),但埋葬屍體後奄美族群的子代品質顯著優於合歡 山族群(ANOVA, p < 0.001、p < 0.001)。
點與誤差線分別代表平均值及標準誤,誤差線上的數字為樣本數。
23
表 1、兩族群繁殖成功率隨溫度變化的變異數分析表(ANOVA table)
族群及溫度間無交互作用,繁殖成功率僅在不同溫度有差異,族群間無顯著差 異。
χ2 Df Pr(>χ2) region 0.217 1 0.6414 temperature 59.087 4 4.51E-12 region × temperature 2.044 4 0.7277
表 2、各溫度下的適存度的變異數分析表(ANOVA table)
(A) 子代數量、(B)子代總重。
子代數量及重量都呈現相同的趨勢,在族群間無顯著差異,僅在溫度間有差。
(A)
χ2 Df Pr(>χ2) region 3.454 1 0.06309 temperature 70.86 4 1.49E-14 region × temperature 1.035 4 0.90445 (B)
χ2 Df Pr(>χ2) region 2.749 1 9.73E-02 temperature 87.207 4 <2e-16 regionn ×
temperature 0.807 4 0.93747
25
表 3、各溫度下埋葬屍體成功率的變異數分析表(ANOVA table)
合歡山族群埋葬屍體成功率顯著高於奄美族群。
χ2 Df Pr(>χ2) region 15.959 1 6.47E-05 temperature 58.496 4 6.00E-12 region × temperature 2.659 4 0.6165
表 4、各溫度下埋葬屍體後繁殖成功率的變異數分析表(ANOVA table)
埋葬屍體後的繁殖成功率兩族群顯著的不同,奄美族群埋葬後的繁殖成功率顯著 高於合歡山族群。
χ2 Df Pr(>χ2) region 18.2592 1 1.93E-05 temperature 24.2052 4 7.27E-05 region × temperature 2.0534 3 0.5614
27
表 5、各溫度下埋葬屍體後適存度的變異數分析表(ANOVA table)
(A) 、(A-1)為子代數量,(B)、(B-1)為子代總重
埋葬屍體後的子代數量及重量都呈現相同的趨勢,兩族群有顯著的差異,奄美族 群埋葬屍體後的子代數量或總重都顯著高於合歡山族群。
(A)
χ2 Df Pr(>χ2) region 43.292 1 4.72E-11 temperature 60.489 4 2.29E-12 region × temperature 3.013 3 0.3897 (B)
χ2 Df Pr(>χ2) region 38.728 1 4.87E-10 temperature 81.926 4 <2.2e-16 region × temperature 1.238 3 0.744